

Physiological Regulation of Photosynthetic-Related Indices, Antioxidant Defense, and Proline Anabolism on Drought Tolerance of Wild Soybean (Glycine soja L.)

 and
and 
Abstract
1. Introduction
2. Results
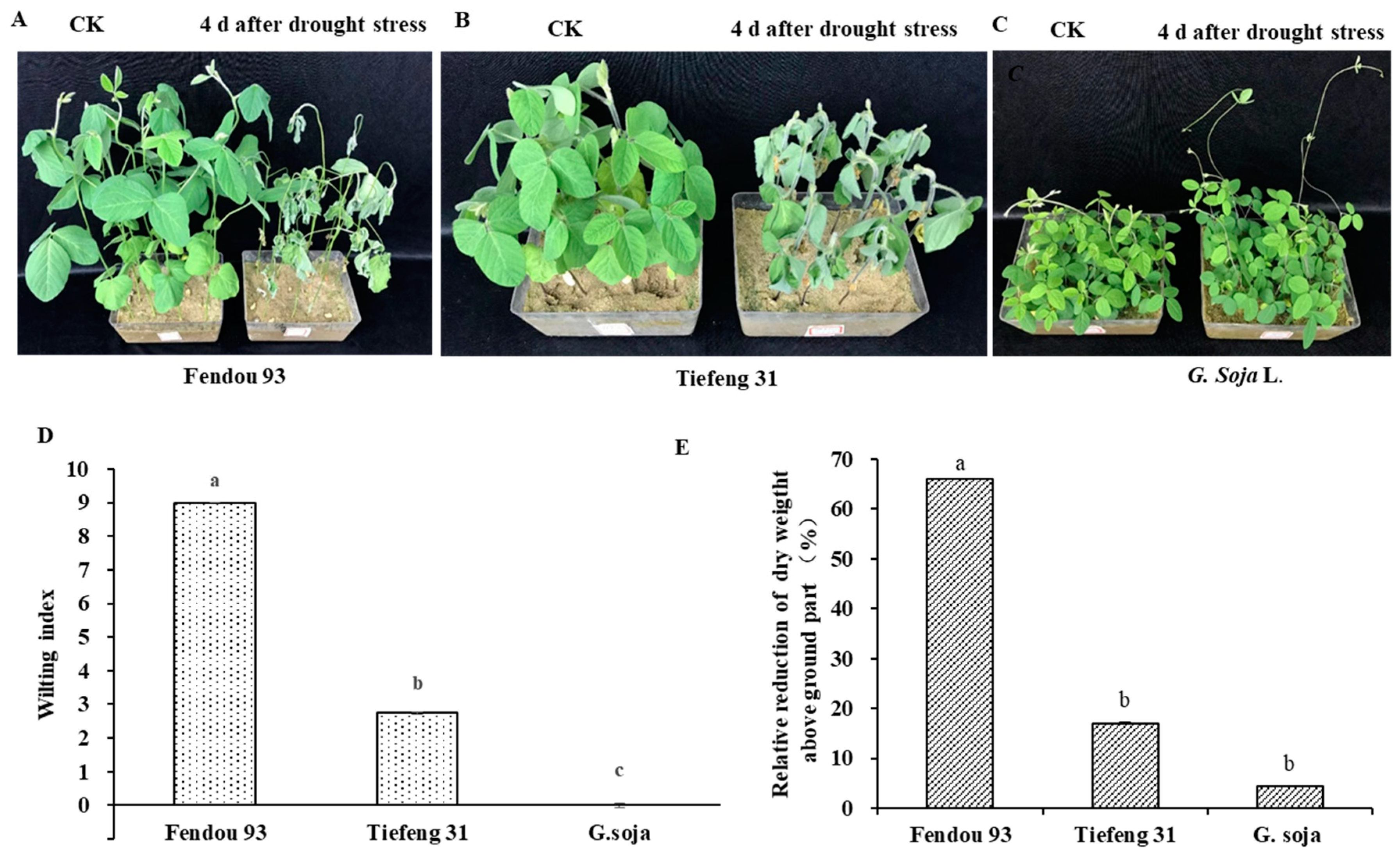
2.1. Drought Stress on Plant Growth
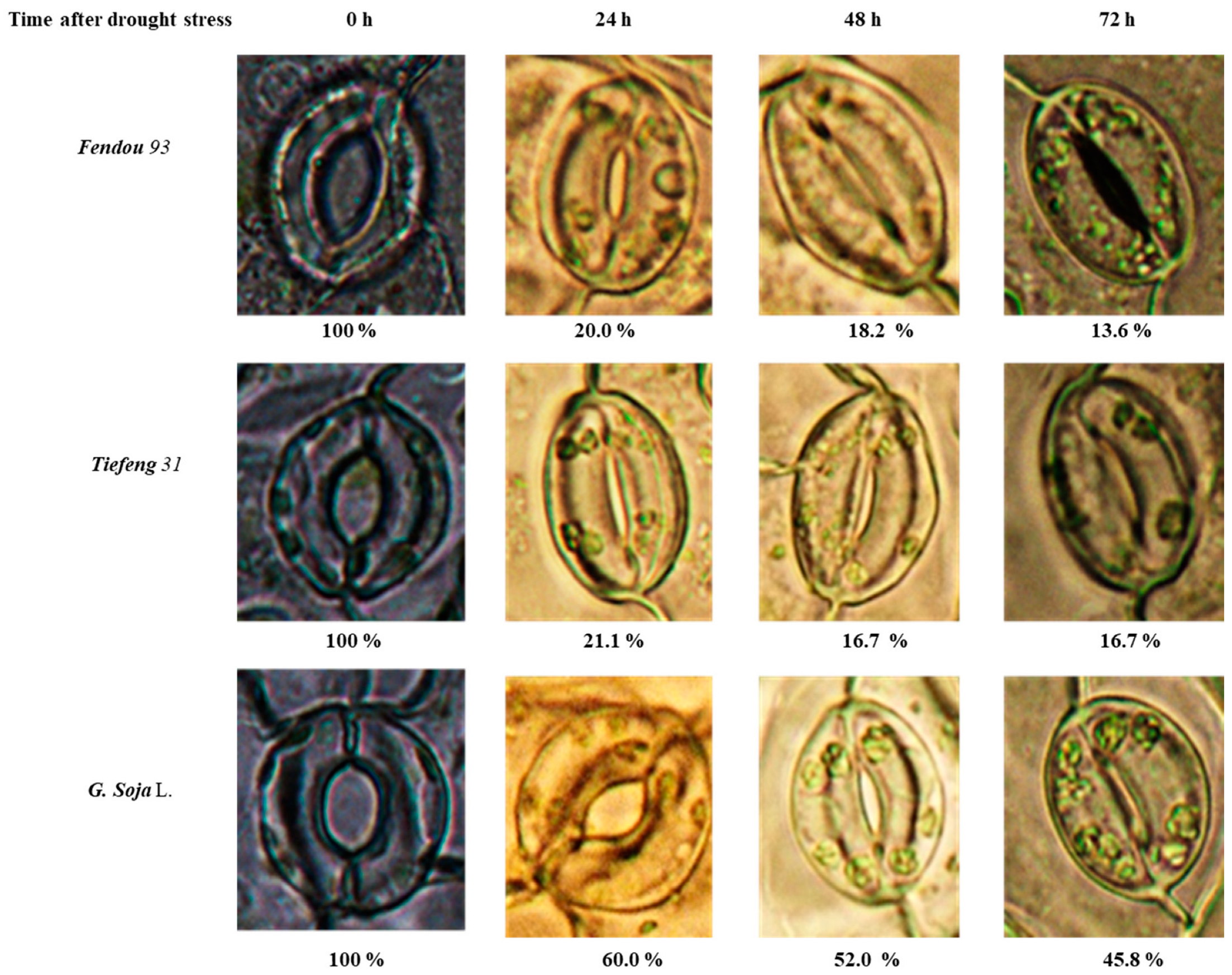
2.2. Stomatal Opening and Photosynthesis-Related Indexes
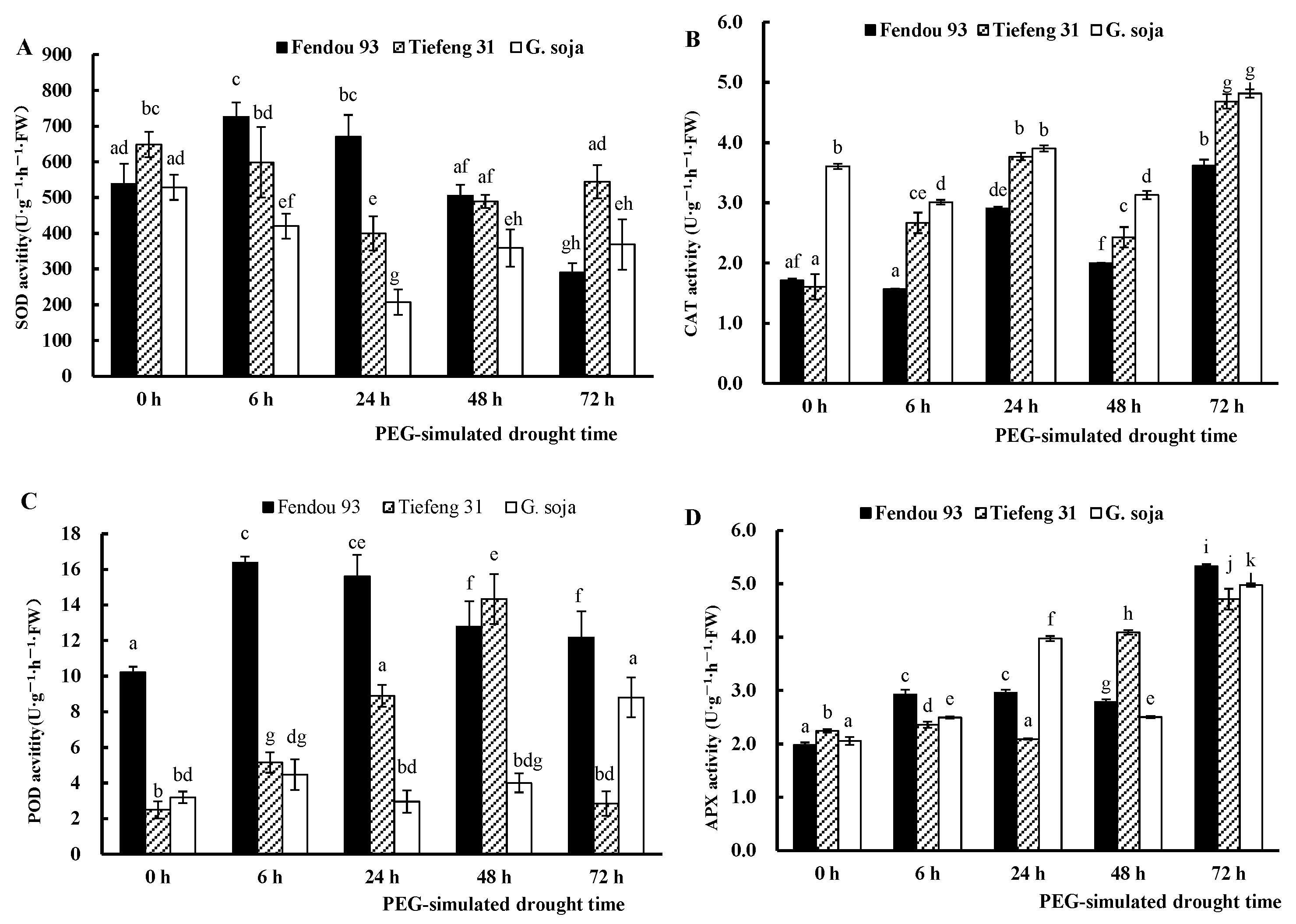
2.3. MDA Content and Antioxidant Enzyme Activities
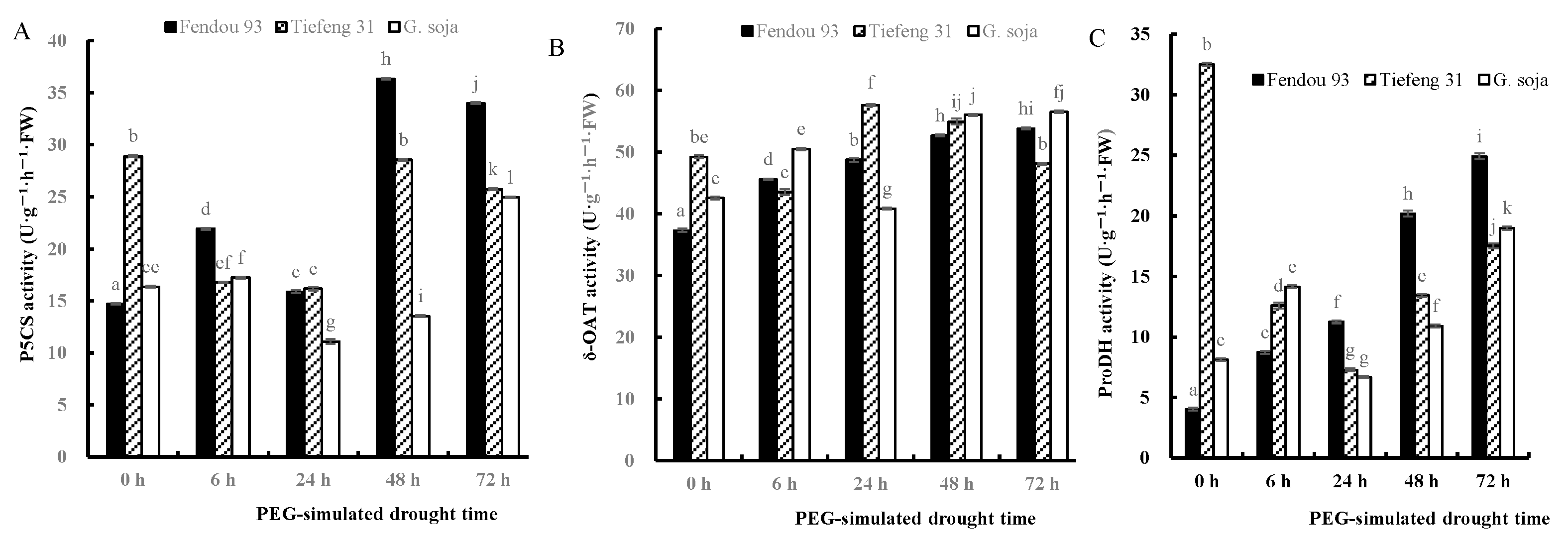
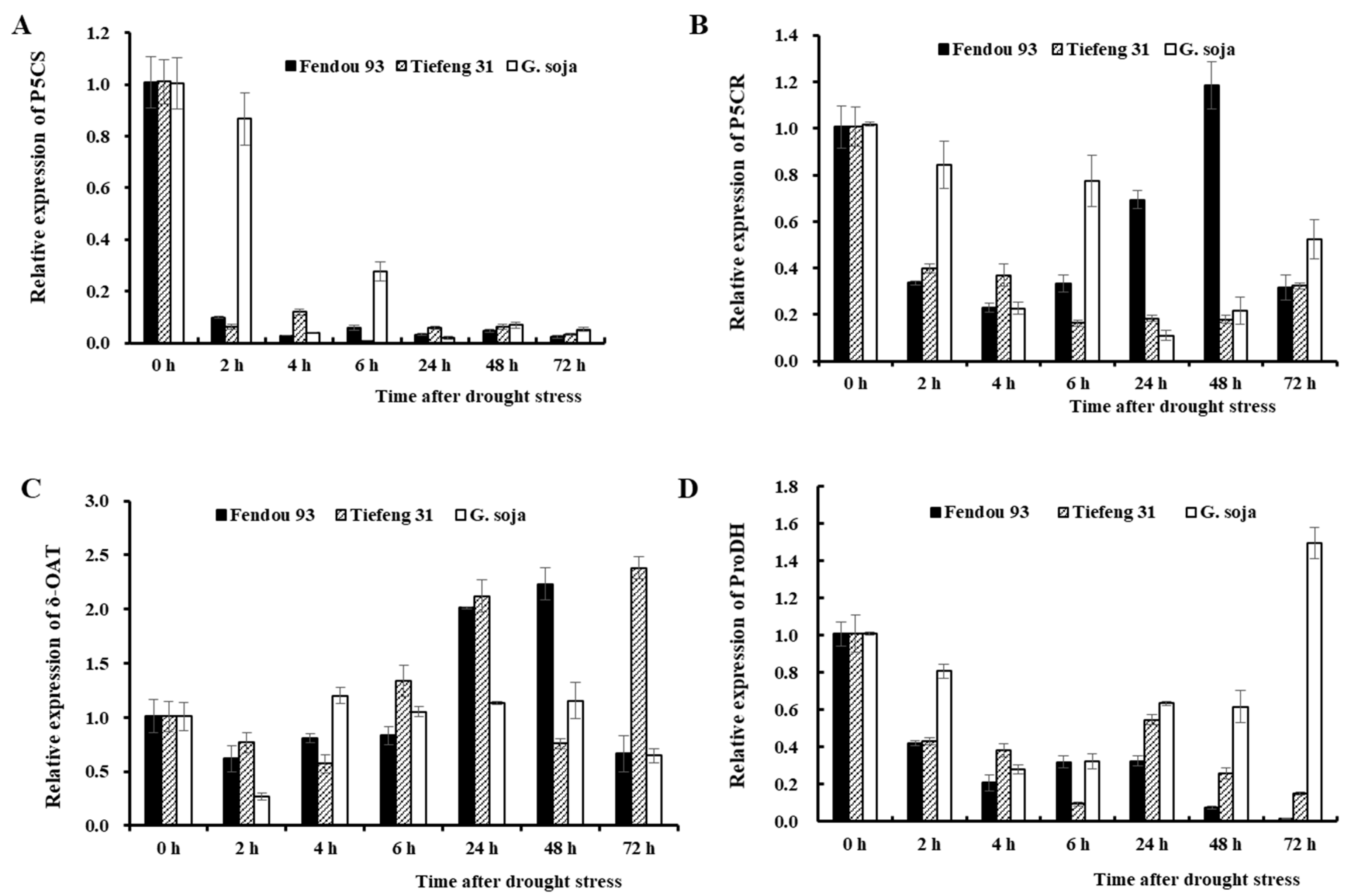
2.4. Osmolytes, Proline Anabolism-Related Enzymes, and Gene Expression
3. Discussion
4. Materials and Method
4.1. Plant Materials and Culture Condition
4.2. Measurement of Wilting Index and Above-Ground Dry Weight
4.3. Measurement of Stomatal Opening and Photosynthetic Indices
4.4. Measurement of Antioxidant Enzyme Activities and Malondialdehyde (MDA) Content
4.5. Measurement of the Contents of Soluble Sugar, Soluble Protein, and Free Proline
4.6. Measurement of Proline-Metabolizing Enzymes
4.7. Relative Expression of Proline-Metabolizing Genes by qRT-PCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, A.; Rico-Medina, A.; Cano-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Dubey, A.; Kumar, A.; Allah, E.F.A.; Hashem, A.; Khan, M.L. Growing More with Less: Breeding and Developing Drought Resilient Soybean to Improve Food Security. Ecolo. Indic. 2019, 105, 425–437. [Google Scholar] [CrossRef]
- Wei, Y.; Jin, J.; Jiang, S.; Ning, S.; Liu, L. Quantitative Response of Soybean Development and Yield to Drought Stress During Different Growth Stages in the Huaibei Plain, China. Agronomy 2018, 8, 97. [Google Scholar] [CrossRef]
- Dogan, E.; Kirnak, H.; Copur, O. Deficit Irrigations during Soybean Reproductive Stages and CROPGRO-Soybean Simulations under Semi-arid Climatic Conditions. Field Crop. Res. 2007, 103, 154–159. [Google Scholar] [CrossRef]
- Poudel, S.; Vennam, R.R.; Shrestha, A.; Reddy, K.R.; Wijewardane, N.K.; Reddy, K.N.; Bheemanahalli, R. Resilience of Soybean Cultivars to Drought Stress During Flowering and Early-seed Setting Stages. Sci. Rep. 2023, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.P.; Tran, L. Potentials Toward Genetic Engineering of Drought-Tolerant Soybean. Crit. Rev. Biotech. 2012, 32, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Satbhai, S.B.; Ristova, D.; Busch, W. Underground Tuning: Quantitative Regulation of Root Growth. J. Exp. Bot. 2015, 66, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Buezo, J.; Sanz-Saez, A.; Moran, J.F.; Soba, D.; Aranjuelo, I.; Esteban, R. Drought Tolerance Response of High-Yielding Soybean Varieties to Mild Drought: Physiological and Photochemical Adjustments. Physiol. Planta. 2019, 166, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Rabara, R.C.; Reese, R.N.; Miller, M.A.; Rohila, J.S.; Subramanian, S.; Shen, Q.J.; Morandi, D.; Buecking, H.; Shulaev, V.; et al. A Toolbox of Genes, Proteins, Metabolites and Promoters for Improving Drought Tolerance in Soybean Includes the Metabolite Coumestrol and Stomatal Development Genes. BMC Genom. 2016, 17, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Guzzo, M.C.; Costamagna, C.; Salloum, M.S.; Rotundo, J.L.; Monteoliva, M.I.; Luna, C.M. Morpho-physiological Traits Associated with Drought Responses in Soybean. Crop. Sci. 2021, 61, 672–688. [Google Scholar] [CrossRef]
- Oraee, A.; Tehranifar, A. Evaluating the Potential Drought Tolerance of Pansy through Its Physiological and Biochemical Responses to Drought and Recovery Periods. Sci. Hortic. 2020, 265, 109225. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF Inoculation and Phosphorus Supplementation Alleviates Drought Induced Growth and Photosynthetic Decline in Nicotiana tabacum by up Regulating Antioxidant Metabolism and Osmolyte Accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Guo, J.; Wang, Y.; Song, C.; Zhou, J.; Qiu, L.; Huang, H.; Wang, Y. A Single Origin and Moderate Bottleneck during Domestication of Soybean (Glycine max): Implications from Microsatellites and Nucleotide Sequences. Ann. Bot. 2010, 106, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.A.; Yang, S.H.; Chung, G. Wild Soybeans: An Opportunistic Resource for Soybean Improvement. In Rediscovery of Landraces as a Resource for the Future; Oscar, G., Ed.; Intech Open: London, UK, 2018; ISBN 978-1-78923-725-2. [Google Scholar]
- Lam, H.M.; Xu, X.; Liu, X.; Chen, W.; Yang, G.; Wong, F.L.; Li, M.W.; He, W.; Qin, N.; Wang, B.; et al. Resequencing of 31 Wild and Cultivated Soybean Genomes Identifies Patterns of Genetic Diversity and Selection. Nat. Genet. 2010, 42, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhou, Q.; He, M.; Zhong, X.; Tang, G. Wilting Index and Root Morphological Characteristics Used as Drought-Tolerance Variety Selection at the Seedling Stage in Soybean (Glycine max L.). Plant Growth Regul. 2020, 92, 29–42. [Google Scholar] [CrossRef]
- Silvente, S.; Sobolev, A.P.; Lara, M. Metabolite Adjustments in Drought Tolerant and Sensitive Soybean Genotypes in Response to Water Stress. PLoS ONE 2012, 7, 38554. [Google Scholar] [CrossRef]
- Zhang, H.; Mittal, N.; Leamy, L.J.; Barazani, O.; Song, B.H. Back into the Wild—Apply Untapped Genetic Diversity of Wild Relatives for Crop Improvement. Evolu. Appli. 2017, 10, 5–24. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Annu. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- King, C.A.; Purcell, L.C.; Brye, K.R. Differential Wilting among Soybean Genotypes in Response to Water Deficit. Crop. Sci. 2009, 49, 290–29822. [Google Scholar] [CrossRef]
- Bagherzadi, L.; Sinclair, T.R.; Zwieniecki, M.; Secchi, F.; Hoffmann, W.; Carter, T.E.; Rufty, T.W. Assessing Water-Related Plant Traits to Explain Slow-Wilting in Soybean PI 471938. J. Crop Improv. 2017, 31, 400–417. [Google Scholar] [CrossRef]
- Nogueira, M.; Livingston, D.; Tuong, T.; Sinclair, T.R. Xylem Vessel Radii Comparison between Soybean Genotypes Differing in Tolerance to Drought. J. Crop. Improv. 2020, 34, 404–413. [Google Scholar] [CrossRef]
- Makhlouf BS, I.; Khalil SR, A.; Saudy, H.S. Efficacy of humic Acids and Chitosan for Enhancing Yield and Sugar Quality of Sugar Beet under Moderate and Severe Drought. J. Soil. Sci. Plant Nutr. 2022, 2, 1676–1691. [Google Scholar] [CrossRef]
- Shaaban, A.; Mahfouz, H.; Megawer, E.A.; Saudy, H.S. Physiological Changes and Nutritional Value of Forage Clitoria Grown in Arid Agro-Ecosystem as Influenced by Plant Density and Water Deficit. J. Soil. Sci. Plant Nutr. 2023, 3, 3735–3750. [Google Scholar] [CrossRef]
- Kusum, N.; Brent, E.; Elise, P. Sap Flux-Scaled Transpiration and Stomatal Conductance Response to Soil and Atmospheric Drought in a Semi-Arid Sagebrush Ecosystem. J. Hydrol. 2012, 464–465, 176–185. [Google Scholar] [CrossRef]
- Flexas, J.; Ribas-Carbo, M.; Diaz-Espejo, A.; Galmes, J.; Medrano, H. Mesophyll Conductance to CO2: Current Knowledge and Future Prospects. Plant Cell. Environ. 2008, 31, 602–621. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Medrano, H. Drought-Inhibition of Photosynthesis in C3 Plants: Stomatal and Non-stomatal Limitations Revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Zivcak, M.; Kalaji, H.M.; Shao, H.B.; Olsovska, K.; Brestic, M. Photosynthetic Proton and Electron Transport in Wheat Leaves under Prolonged Moderate Drought Stress. J. Photochem. Photobiol. 2014, 137, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, C.; Pan, D.; Zhang, Y.; Luo, B.; Ji, J. Effects of Drought Stress on Photosynthesis and Chlorophyll Fluorescence Images of Soybean (Glycine max) Seedlings. Int. J. Agric. Biol. Eng. 2018, 11, 196–201. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, H.; Teng, A.; Zhang, C.; Lei, L.; Ba, Y.; Wang, Z. Photosynthetic Characteristics, Yield and Quality of Sunflower Response to Deficit Irrigation in a Cold and Arid Environment. Front. Plant Sci. 2023, 14, 1280347. [Google Scholar] [CrossRef]
- Stolf-Moreira, R.; Medri, M.E.; Neumaier, N.; Lemos, N.G.; Pimenta, J.A.; Tobita, S.; Brogin, R.L.; Marcelino-Guimaraes, F.C.; Oliveira, M.C.N.; Farias, J.R.B.; et al. Soybean Physiology and Gene Expression during Drought. Genet. Mol. Res. 2010, 9, 1946–1956. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natic´, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Agarwal, S.; Sairam, R.K.; Srivastava, G.C.; Meena, R.C. Changes in Antioxidant Enzymes Activity and Oxidative Stress by Abscisic Acid and Salicylic Acid in Wheat Genotypes. Biol. Plant. 2005, 49, 541–550. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Sloane, R.J.; Patterson, R.P.; Carter, T.E., Jr. Field Drought Tolerance of a Soybean Plant Introduction. Crop. Sci. 1990, 30, 118–123. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Guttikonda, S.K.; Tran, L.P.; Nguyen, H.T. Physiological and Molecular Approaches to Improve Drought Resistance in Soybean. Plant Cell. Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef]
- Dubrovna, O.V.; Mykhalska, S.I.; Komisarenko, A.G. Using Proline Metabolism Genes in Plant Genetic Engineering. Cytol. Genet. 2022, 56, 361–378. [Google Scholar] [CrossRef]
- Deng, G.; Liang, J.; Xu, D.; Long, H.; Pan, Z.; Yu, M. The Relationship between Proline Content, the Expression Level of P5CS (Δ1-pyrroline-5-carboxylate synthetase), and Drought Tolerance in Tibetan Hulless Barley (Hordeum vulgare var. nudum). Russ. J. Plant Phys. 2013, 60, 693–700. [Google Scholar] [CrossRef]
- Yosefi, M.; Yosefi, M.; Sharafzadeh, S.; Amiri, B. Application of Jasmonic Acid can Mitigate Water Deficit Stress in Cotton through Yield-Related Physiological Properties. Acta. Agro. 2018, 71, 1741. [Google Scholar] [CrossRef]
- Nyawade, S.; Gitari, H.I.; Karanja, N.N.; Gachene, C.K.K.; Schulte-Geldermann, E.; Sharma, K.; Parker, M.L. Enhancing Climate Resilience of Rain-Fed Potato through Legume Intercropping and Silicon Application. Front. Sustain. Food 2020, 4, 566345. [Google Scholar] [CrossRef]
- Garg, A.K.; Kim, J.K.; Owens, T.G.; Ranwala, A.P.; Choi, Y.D.; Kochian, L.V.; Wu, R.J. Trehalose Accumulation in Rice Plants Confers High Tolerance Levels to Different Abiotic Stresses. Proc. Natl. Acad. Sci. USA 2002, 99, 15898–15903. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, Salt, and Temperature Stress-induced Metabolic Rearrangements and Regulatory Networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Norwood, M.; Truesdale, M.R.; Richter, A.; Scott, P. Photosynthetic Carbohydrate Metabolism in the Resurrection Plant Craterostigma plantagineum. J. Exp. Bot. 2000, 51, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, A.; Bochicchio, A.; Vazzana, C.; Lindsey, G.; Farrant, J. Changes in Leaf Hexokinase Activity and Metabolite Levels in Response to Drying in the Desiccation-Tolerant Species Sporobolus stapfianus and Xerophyta viscosa. J. Exp. Bot. 2001, 52, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Rontein, D.; Basset, G.; Hanson, A.D. Metabolic Engineering of Osmoprotectant Accumulation in Plants. Metab. Eng. 2002, 4, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Delorge, I.; Janiak, M.; Carpentier, S.; Van Dijck, P. Fine Tuning of Trehalose Biosynthesis and Hydrolysis as Novel Tools for the Generation of Abiotic Stress Tolerant Plants. Front. Plant Sci. 2014, 5, 147. [Google Scholar] [CrossRef]
- Khater, M.; Dawood, M.G.; Sadak, M.S.; Shalaby, M.; El-Awadi, M.; El-Din, K.G. Enhancement the Performance of Cowpea Plants Grown under Drought Conditions via Trehalose Application. Middle East. J. Agric. Res. 2018, 7, 782–800. [Google Scholar]
- Li, H.W.; Zang, B.S.; Deng, X.W.; Wang, X.P. Overexpression of the Trehalose-6-Phosphate Synthase Gene OsTPS1 Enhances Abiotic Stress Tolerance in Rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef]
- de Ronde, J.A.; Laurie, R.N.; Caetano, T.; Greyling, M.M.; Kerepesi, I. Comparative Study between Transgenic and Non-transgenic Soybean Lines Proved Transgenic Lines to be more Drought Tolerant. Euphytica 2004, 138, 123–132. [Google Scholar] [CrossRef]
- Dien, D.C.; Mochizuki, T.; Yamakawa, T. Effect of Various Drought Stresses and Subsequent Recovery on Proline, Total Soluble Sugar and Starch Metabolisms in Rice (Oryza sativa L.) Varieties. Plant Produ. Sci. 2019, 22, 530–545. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Chung, I.M. Physiological and Molecular Level Studies on the Toxicity of Silver Nanoparticles in Germinating Seedlings of Mung Bean (Vigna radiata L.). Acta Physiol. Plant. 2015, 37, 1–11. [Google Scholar] [CrossRef]
- Nie, L.; Guo, J.; Liu, Y. Study on Cold-stress Tolerance of Transgenic Alfalfa (Medicago sativa L.) with AmDHN Gene. Acta. Bot. Boreali-Occident. Sin. 2014, 34, 1727–1734. (In Chinese) [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. California Agr. Exp. Station. Cir. 1938, 347, 1–39. [Google Scholar]
- He, M.D.; Jiang, Y.J.; Liu, L.L.; Zhong, X.B.; Zhao, Y.; Ma, W.J.; Tang, G.X. Benefits of High Nitrogen Fertilizer on Nitrogen Metabolism, Nitrogen Transfer Rate, Root System Architecture and Grain Yield of Wheat (Triticum aestivum L.) under Water Deficit at Heading Stage. Acta. Physiol. Plant. 2022, 44, 121. [Google Scholar] [CrossRef]
- Amako, K.; Chen, G.X.; Asada, K. Separate Assays Specific for Ascorbate Peroxidase and Guaiacol Peroxidase and for the Chloroplastic and Cytosolic Isozymes of Ascorbate Peroxidase in Plants. Plant Cell Physiol. 1994, 35, 497–504. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen-Peroxide is Scavenged by Ascorbate-Specific Peroxidase in Spinach-Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hayzer, D.J.; Leisinger, T. The Gene-Enzyme Relationships of Proline Riosynthesis in Escherichia coli. J. General. Micro. 1979, 118, 287–293. [Google Scholar]
- Lutts, S.; Majerus, V.; Kinet, J.M. NaCl Effects on Proline Metabolism in Rice (Oryza sativa L.) Seedlings. Physiol. Plant. 1999, 105, 450–458. [Google Scholar] [CrossRef]
- Vogel, R.H.; Kopac, M.J. Some Properties of Ornithine--Transaminase from Neurospore. Biochem. Biphys. Acta. 1960, 37, 539–540. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (bp) | Tm (°C) | Gene Identity |
|---|---|---|---|
| δ-OAT | F: AGGGTTTGCAGAGGAAGTAGG | 60.0 | DQ224372.1 |
| R: CAGAGGTTCCCTTTGCCTGA | 60.0 | ||
| P5CR | F: GGGTTCCGTGGAACACTGAT | 59.0 | X16352.1 |
| R: AGCTCGAAAAGACTGTTATGGC | 59.0 | ||
| ProDH | F: GGTGTCGACAAAGAGGCTG | 60.0 | AY492003.1 |
| R: GCGTCTTCCACACCGTACA | 60.0 | ||
| P5CS | F: ATGGCAAGGCGGATTGTACT | 59.74 | NM_001251224.1 |
| R: TTCAACTGTGCATGCCAACG | 59.97 | ||
| β-Tubulin | F: GCTCCAACACAGGGGAAAATG | 59.73 | NM_001252709.2 |
| R: ACTTCCCCGTCGGATCTATG | 58.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.; Zhang, W.; Wang, G.; Hu, Y.; Zhong, X.; Tang, G. Physiological Regulation of Photosynthetic-Related Indices, Antioxidant Defense, and Proline Anabolism on Drought Tolerance of Wild Soybean (Glycine soja L.). Plants 2024, 13, 880. https://doi.org/10.3390/plants13060880
Lin S, Zhang W, Wang G, Hu Y, Zhong X, Tang G. Physiological Regulation of Photosynthetic-Related Indices, Antioxidant Defense, and Proline Anabolism on Drought Tolerance of Wild Soybean (Glycine soja L.). Plants. 2024; 13(6):880. https://doi.org/10.3390/plants13060880
Chicago/Turabian StyleLin, Song, Weimei Zhang, Guifeng Wang, Yunxiang Hu, Xuanbo Zhong, and Guixiang Tang. 2024. "Physiological Regulation of Photosynthetic-Related Indices, Antioxidant Defense, and Proline Anabolism on Drought Tolerance of Wild Soybean (Glycine soja L.)" Plants 13, no. 6: 880. https://doi.org/10.3390/plants13060880
APA StyleLin, S., Zhang, W., Wang, G., Hu, Y., Zhong, X., & Tang, G. (2024). Physiological Regulation of Photosynthetic-Related Indices, Antioxidant Defense, and Proline Anabolism on Drought Tolerance of Wild Soybean (Glycine soja L.). Plants, 13(6), 880. https://doi.org/10.3390/plants13060880