

Ozone Treatment as an Approach to Induce Specialized Compounds in Melissa officinalis Plants


 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Visible Injury and Plant Biometric Traits
2.2. Leaf Ecophysiological Traits
2.3. Leaf Biochemical Traits
2.3.1. Hydrogen Peroxide Content and Oxidative Damage
2.3.2. Photosynthetic Pigments and Low-Molecular-Weight Antioxidants
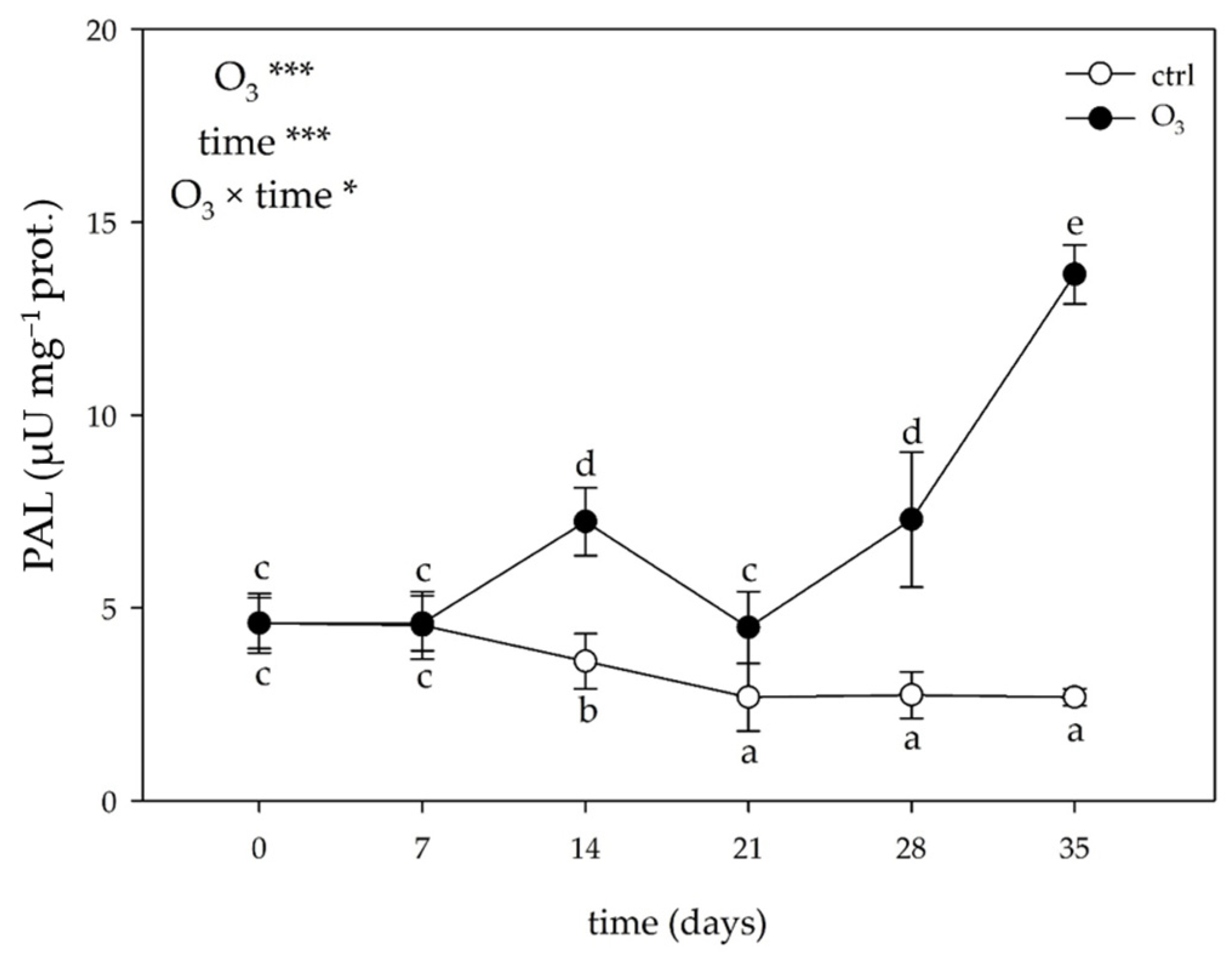
2.3.3. Activity of Phenylalanine Ammonia-Lyase
2.3.4. Total Antioxidant Activity
2.4. Leaf Molecular Traits
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Plant Biometric Traits
4.3. Leaf Physiological Traits
4.4. Leaf Biochemical Traits
4.4.1. Assessment of Hydrogen Peroxide Content and Membrane Damage
4.4.2. Determination of Photosynthetic Pigments and Phenylpropanoids
4.4.3. Activity of Phenylalanine Ammonia-Lyase
4.4.4. Total Antioxidant Activity
4.5. Leaf Molecular Traits
4.5.1. RNA Isolation and First Strand cDNA Synthesis
4.5.2. Expression Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Xu, J.; Li, R.; Ge, Y.; Li, Y.; Li, R. Plants’ responses to abiotic stress: Mechanisms and strategies. Int. J. Mol. Sci. 2023, 24, 10915. [Google Scholar] [CrossRef]
- Son, S.; Park, S.R. Climate change impedes plant immunity mechanisms. Front. Plant Sci. 2022, 13, 1032820. [Google Scholar] [CrossRef]
- Munns, R.; Millar, A.H. Seven plant capacities to adapt to abiotic stress. J. Exp. Bot. 2023, 74, 4308–4323. [Google Scholar] [CrossRef]
- Strasser, R. Protein quality control in the endoplasmic reticulum of plants. Ann. Rew. Plant Biol. 2018, 69, 147–172. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.-B.; Chu, L.-Y.; Shoa, M.-A.; Jaleel, C.A.; Mi, H.-M. Higher plant antioxidants and redox signaling under environmental stresses. Comput. Rendus Biol. 2008, 331, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.A.; Ghosh, A.; Agrawal, M.; Agrawal, S.B. Secondary metabolites responses of plants exposed to ozone: An update. Environ. Sci. Pollut. Res. 2023, 30, 88281–88312. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Mishra, P.; Mishra, A.K. Effect of climate change on plant secondary metabolism: An ecological perspective. In Evolutionary Diversity as a Source for Anticancer Molecules; Srivastava, A.K., Kumar, V., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 47–76. [Google Scholar]
- Rejeb, L.B.; Pastor, V.; Mauch-Mani, B. Plant responses to simultaneous biotic and abiotic stress: Molecular mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Calzone, A.; Tonelli, M.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E. Significance of phenylpropanoid pathways in the response of two pomegranate cultivars to salinity and ozone stress. Environ. Exp. Bot. 2023, 208, 105249. [Google Scholar] [CrossRef]
- Agathokleous, E.; Belz, R.G.; Kitao, M.; Koike, T.; Calabrese, E.J. Does the root to shoot ratio show a hormetic response to stress? An ecological and environmental perspective. J. For. Res. 2019, 30, 1569–1580. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.L.; Torres-Pacheco, I. Plant hormesis management with biostimulants of biotic origin in agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation: A tool for enriching the bioactive composition of foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef]
- Marchica, A.; Ascrizzi, R.; Flamini, G.; Cotrozzi, L.; Tonelli, M.; Lorenzini, G.; Nali, C.; Pellegrini, E. Ozone as eustress for enhancing secondary metabolites and bioactive properties in Salvia officinalis. Ind. Crops Prod. 2021, 170, 113730. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef]
- Pellegrini, E.; Campanella, A.; Cotrozzi, L.; Tonelli, M.; Nali, C.; Lorenzini, G. Ozone primes changes in phytochemical parameters in the medicinal herb Hypericum perforatum (St. John’s wort). Ind. Crops Prod. 2018, 126, 119–128. [Google Scholar] [CrossRef]
- Modesti, M.; Marchica, A.; Pisuttu, C.; Risoli, S.; Pellegrini, E.; Bellincontro, A.; Mencarelli, F.; Tonutti, P.; Nali, C. Ozone-induced biochemical and molecular changes in Vitis vinifera leaves and responses to Botrytis cinerea infections. Antioxidants 2023, 12, 343. [Google Scholar] [CrossRef]
- Pisuttu, C.; Risoli, S.; Moncini, L.; Nali, C.; Pellegrini, E.; Sarrocco, S. Sustainable strategies to counteract mycotoxins contamination and cowpea weevil in chickpea seeds during post-harvest. Toxins 2023, 15, 61. [Google Scholar] [CrossRef] [PubMed]
- Nickhil, C.; Mohapatra, D.; Kar, A.; Giri, S.K.; Tripathi, M.K.; Sharma, Y. Gaseous ozone treatment of chickpea grains, part I: Effect on protein, amino acid, fatty acid, mineral content, and microstructure. Food Chem. 2021, 345, 128850. [Google Scholar] [CrossRef]
- Humbal, A.; Pathak, B. Influece of exogenous elicitors on the production of secondary metabolite in plants: A review (“VSI: Secondary metabolites”). Plant Stress 2023, 8, 100166. [Google Scholar] [CrossRef]
- Jolivet, Y.; Bagard, M.; Cabané, M.; Vaultier, M.N.; Gandin, A.; Afif, D.; Dizengremel, P.; Le Thiec, D. Deciphering the ozone-induced changes in cellular processes: A prerequisite for ozone risk assessment at the tree and forest levels. Ann. For. Sci. 2016, 73, 923–943. [Google Scholar] [CrossRef]
- Grulke, N.E.; Heath, R.L. Ozone effects on plants in natural ecosystems. Plant Biol. 2020, 22, 12–37. [Google Scholar] [CrossRef]
- Shakeri, A.; Sahebkar, A.; Javadi, B. Melissa officinalis L.–A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2016, 188, 204–228. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B.; Dresler, S.; Stasińska-Jakubas, M.; Wójciak, M.; Sowa, I.; Matraszek-Gawron, R. NaCl-induced elicitation alters physiology and increases accumulation of phenolic compounds in Melissa officinalis L. Int. J. Mol. Sci. 2021, 22, 6844. [Google Scholar] [CrossRef] [PubMed]
- Döring, A.S.; Pellegrini, E.; Campanella, A.; Trivellini, A.; Gennai, C.; Petersen, M.; Nali, C.; Lorenzini, G. How sensitive is Melissa officinalis to realistic ozone concentrations? Plant Physiol. Biochem. 2014, 74, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Döring, A.S.; Pellegrini, E.; Della Bartola, M.; Nali, C.; Lorenzini, G.; Petersen, M. How do background ozone concentrations affect the biosynthesis of rosmarinic acid in Melissa officinalis? J. Plant Physiol. 2014, 171, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Döring, A.S.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Petersen, M.; Pellegrini, E. Deciphering the role of low molecular weight antioxidants in the sensitivity of Melissa officinalis L. to realistic ozone concentrations. Ind. Crops Prod. 2020, 150, 112369. [Google Scholar] [CrossRef]
- Pellegrini, E.; Carucci, M.G.; Campanella, A.; Lorenzini, G.; Nali, C. Ozone stress in Melissa officinalis plants assessed by photosynthetic function. Environ. Exp. Bot. 2011, 73, 94–101. [Google Scholar] [CrossRef]
- Pellegrini, E.; Trivellini, A.; Campanella, A.; Francini, A.; Lorenzini, G.; Nali, C.; Vernieri, P. Signaling molecules and cell death in Melissa officinalis plants exposed to ozone. Plant Cell Rep. 2013, 32, 1965–1980. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, M.; Pellegrini, E.; D’Angiolillo, F.; Petersen, M.; Nali, C.; Pistelli, L.; Lorenzini, G. Ozone-elicited secondary metabolites in shoot cultures of Melissa officinalis L. Plant Cell Tissue Organ Cult. 2015, 120, 617–629. [Google Scholar] [CrossRef]
- Hoshika, Y.; Fares, S.; Pellegrini, E.; Conte, A.; Paoletti, E. Water use strategy affects avoidance of ozone stress by stomatal closure in Mediterranean trees—A modelling analysis. Plant Cell Environ. 2020, 43, 611–623. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Stasińska-Jakubas, M.; Hawrylak-Nowak, B.; Dresler, S.; Wójciak, M.; Rubinowska, K. Application of chitosan lactate, selenite, and salicylic acid as an approach to induce biological responses and enhance secondary metabolism in Melissa officinalis L. Ind. Crops Prod. 2023, 205, 117571. [Google Scholar] [CrossRef]
- Guo, C.; Wang, X.; Wang, Q.; Zhao, Z.; Xie, B.; Xu, L.; Zhang, R. Plant defense mechanisms against ozone stress: Insights from secondary metabolism. Environ. Exp. Bot. 2023, 2023, 105553. [Google Scholar] [CrossRef]
- Nowroz, F.; Hasanuzzaman, M.; Siddika, A.; Parvin, K.; Caparros, P.G.; Nahar, K.; Prasad, P.V. Elevated tropospheric ozone and crop production: Potential negative effects and plant defense mechanisms. Front. Plant Sci. 2024, 14, 1244515. [Google Scholar] [CrossRef]
- Godínez-Mendoza, P.L.; Rico-Chávez, A.K.; Ferrusquía-Jimenez, N.I.; Carbajal-Valenzuela, I.A.; Villagómez-Aranda, A.L.; Torres-Pacheco, I.; Guevara-González, R.G. Plant hormesis: Revising of the concepts of biostimulation, elicitation and their application in a sustainable agricultural production. Sci. Total Environ. 2023, 894, 164883. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, E.; Grulke, N.E. Ozone exposure and stomatal sluggishness in different plant physiognomic classes. Environ. Pollut. 2010, 158, 2664–2671. [Google Scholar] [CrossRef] [PubMed]
- Penella, C.; Landi, M.; Guidi, L.; Nebauer, S.G.; Pellegrini, E.; San Bautista, A.; Remorini, D.; Nali, C.; López-Galarza, S.; Calatayud, A. Salt-tolerant rootstock increases yield of pepper under salinity through maintenance of photosynthetic performance and sinks strength. J. Plant Physiol. 2016, 193, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vougeleka, V.; Risoli, S.; Saitanis, C.; Agathokleous, E.; Ntatsi, G.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Pisuttu, C. Exogenous application of melatonin protects bean and tobacco plants against ozone damage by improving antioxidant enzyme activities, enhancing photosynthetic performance, and preventing membrane damage. Environ. Pollut. 2024, 343, 123180. [Google Scholar] [CrossRef] [PubMed]
- Czarnocka, W.; Karpińsky, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Hoshika, Y.; Dusart, N.; Cotrozzi, L.; Gérard, J.; Nali, C.; Vaultier, M.-N.; Jolivet, Y.; Lorenzini, G.; Paoletti, E. Antioxidative responses of three oak species under ozone and water stress conditions. Sci. Total Environ. 2019, 647, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, C.; Guidi, L.; Sebastiani, F.; Tattini, M. Isoprenoids and phenylpropanoids are key components of the antioxidant defense system of plants facing severe excess light stress. Environ. Exp. Bot. 2015, 119, 54–62. [Google Scholar] [CrossRef]
- Havaux, M.; Eymery, F.; Porfirovo, S.; Rey, P.; Dörmann, P. Vitamin E protects against photoinhibition and photoxidative stress in Arabidopsis thaliana. Plant Cell 2005, 17, 3451–3469. [Google Scholar] [CrossRef]
- Saleem, A.; Lopnen, J.; Pihlaja, K.; Oksanen, E. Effects of long-term open-field ozone exposure on leaf phenolics of European silver birch (Betula pendula ROTH). J. Chem. Ecol. 2001, 27, 1049–1062. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Campanella, A.; Pellegrini, E.; Lorenzini, G.; Nali, C.; Paoletti, E. Phenylpropanoids are key players in the antioxidant defense to ozone of European ash, Fraxinus excelsior. Environ. Sci. Pollut. Res. 2018, 25, 8137–8147. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Agathokleous, E.; Araminiene, V.; Belz, R.G.; Calatayud, V.; De Marco, A.; Domingos, M.; Feng, Z.; Hoshika, Y.; Kitao, M.; Koike, T.; et al. A quantitative assessment of hormetic responses of plants to ozone. Environ. Res. 2019, 176, 108527. [Google Scholar] [CrossRef]
- Li, Z.; Yang, J.; Shang, B.; Agathokleous, E.; Rubert-Nason, K.F.; Xu, Y.; Feng, Z. Nonlinear responses of foliar phenylpropanoids to increasing O3 exposure: Ecological implications in a Populus model system. Sci. Tot. Environ. 2021, 767, 144358. [Google Scholar] [CrossRef] [PubMed]
- Marchica, A.; Cotrozzi, L.; Detti, R.; Lorenzini, G.; Pellegrini, E.; Petersen, M.; Nali, C. The biosynthesis of phenolic compounds is an integrated defence mechanism to prevent ozone injury in Salvia officinalis. Antioxidants 2020, 9, 1274. [Google Scholar] [CrossRef]
- Petersen, M.; Abdullah, Y.; Benner, J.; Eberle, D.; Gehlen, K.; Hücherig, S.; Janiak, V.; Kim, K.H.; Sander, M.; Weitzel, C.; et al. Evolution of rosmarinic acid biosynthesis. Phytochemistry 2009, 70, 1663–1679. [Google Scholar] [CrossRef]
- Asensi-Fabado, A.; Garcia-Breijo, F.J.; Reig-Arminana, J. Ozone-induced reductions in below-ground biomass: An anatomical approach in potato. Plant Cell Environ. 2010, 33, 1070–1083. [Google Scholar] [CrossRef]
- Kulak, M.; Gul, F.; Sekeroglu, N. Changes in growth parameter and essential oil composition of sage (Salvia officinalis L.) leaves in response to various salt stresses. Ind. Crops Prod. 2020, 145, 112078. [Google Scholar] [CrossRef]
- Pellegrini, E. PSII photochemistry is the primary target of oxidative stress imposed by ozone in Tilia americana. Urban For. Urban Green. 2014, 13, 94–102. [Google Scholar] [CrossRef]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive oxygen species and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Bensadoun, A.; Weinsteinm, D. Assay of proteins in the presence of interfering materials. Analyt. Biochem. 1976, 70, 241–251. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Deemer, E.K.; Prior, R.L.; Huang, D. Novel fluorometric assay for hydroxyl radical prevention capacity using fluorescein as the probe. J. Agric. Food Chem. 2002, 50, 2772–2777. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G.; Bartley, G.E.; Scolnik, P.A. Regulation of carotenoid biosynthesis during tomato development. Plant Cell 1993, 5, 379–387. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | O3 | p | |
|---|---|---|---|
| Height (cm) | 39.1 ± 3.23 | 36.8 ± 2.64 | * |
| Total FW (g) | 115.4 ± 13.06 | 102.4 ± 10.26 | * |
| Total DW (g) | 13.1 ± 2.07 | 10.9 ± 1.51 | ** |
| Total foliar area (cm2) | 3769 ± 1037.5 | 2470 ± 301.8 | *** |
| Root to shoot ratio | 0.32 ± 0.054 | 0.43 ± 0.059 | ** |
| Primers | Sequences |
|---|---|
| β-Actin | F: 5-GGATGATATGGAGAAGATYTGTGGC-3′ |
| R: 5′-AGATCACGMCCAGCRAGATC-3′ | |
| PAL | F: 5′-ATACATATGGCAGAGAACGGTCATCATGATTCC-3′ |
| R: 5′-ATACATATGCTAGCAGATAGGCAGAGGTCCACCATT-3′ | |
| 4CL | F: 5′-ATGGAGAACCCGGCAGGCCAAG-3′ |
| R: 5-GACTGCAGCTGCTAATCTTGATCT-3′ | |
| RAS | F: 5′ATGAGGATCGATATCAAGGAC-3′ |
| R: 5′TCAAATCTCATAAAACAACTTCTCAA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scimone, G.; Carucci, M.G.; Risoli, S.; Pisuttu, C.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Petersen, M. Ozone Treatment as an Approach to Induce Specialized Compounds in Melissa officinalis Plants. Plants 2024, 13, 933. https://doi.org/10.3390/plants13070933
Scimone G, Carucci MG, Risoli S, Pisuttu C, Cotrozzi L, Lorenzini G, Nali C, Pellegrini E, Petersen M. Ozone Treatment as an Approach to Induce Specialized Compounds in Melissa officinalis Plants. Plants. 2024; 13(7):933. https://doi.org/10.3390/plants13070933
Chicago/Turabian StyleScimone, Giulia, Maria Giovanna Carucci, Samuele Risoli, Claudia Pisuttu, Lorenzo Cotrozzi, Giacomo Lorenzini, Cristina Nali, Elisa Pellegrini, and Maike Petersen. 2024. "Ozone Treatment as an Approach to Induce Specialized Compounds in Melissa officinalis Plants" Plants 13, no. 7: 933. https://doi.org/10.3390/plants13070933
APA StyleScimone, G., Carucci, M. G., Risoli, S., Pisuttu, C., Cotrozzi, L., Lorenzini, G., Nali, C., Pellegrini, E., & Petersen, M. (2024). Ozone Treatment as an Approach to Induce Specialized Compounds in Melissa officinalis Plants. Plants, 13(7), 933. https://doi.org/10.3390/plants13070933