
Genome-Wide Analysis of the GLK Gene Family and Its Expression at Different Leaf Ages in the Citrus Cultivar Kanpei

 ,
,
Abstract
:1. Introduction
2. Results and Analysis
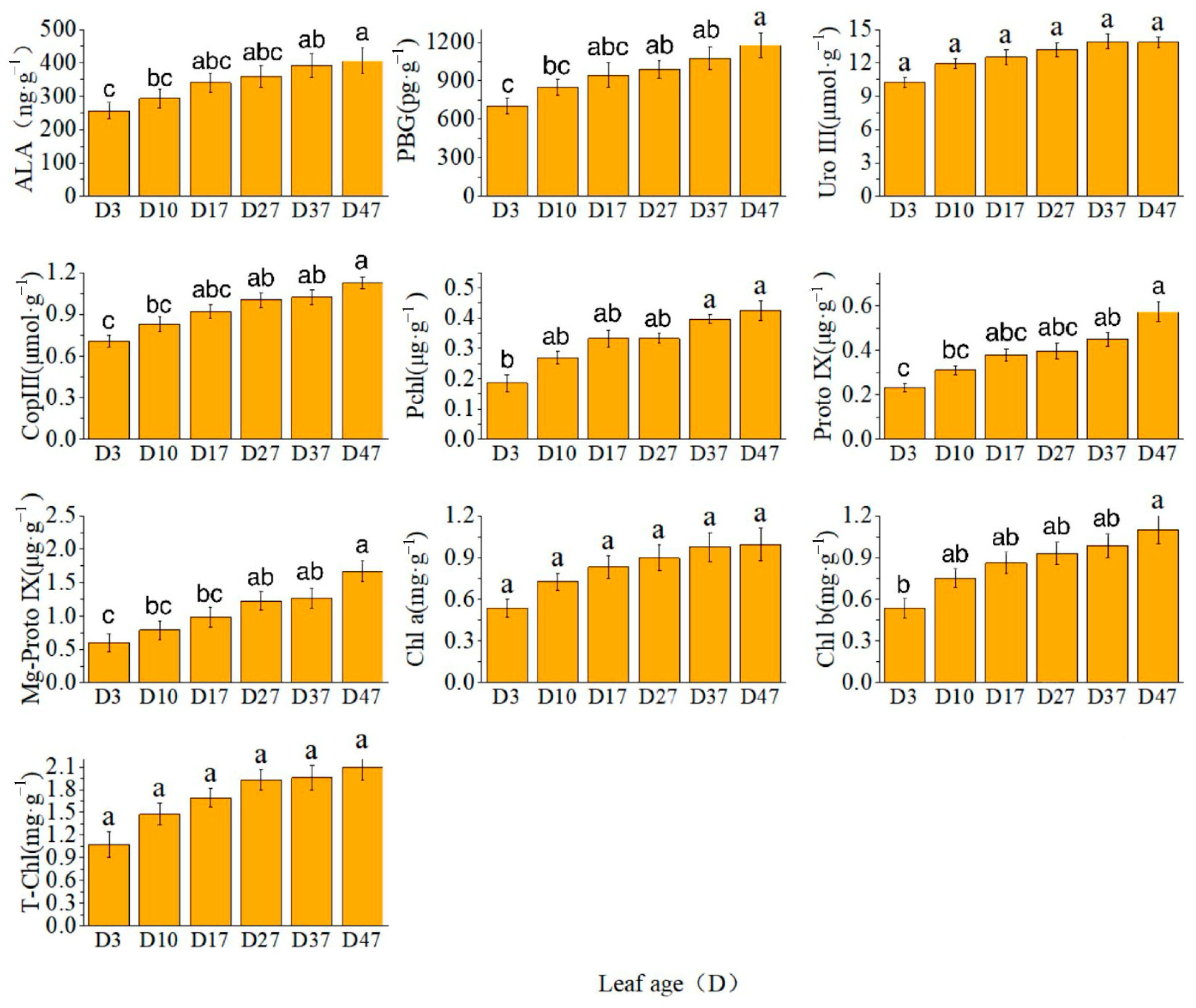
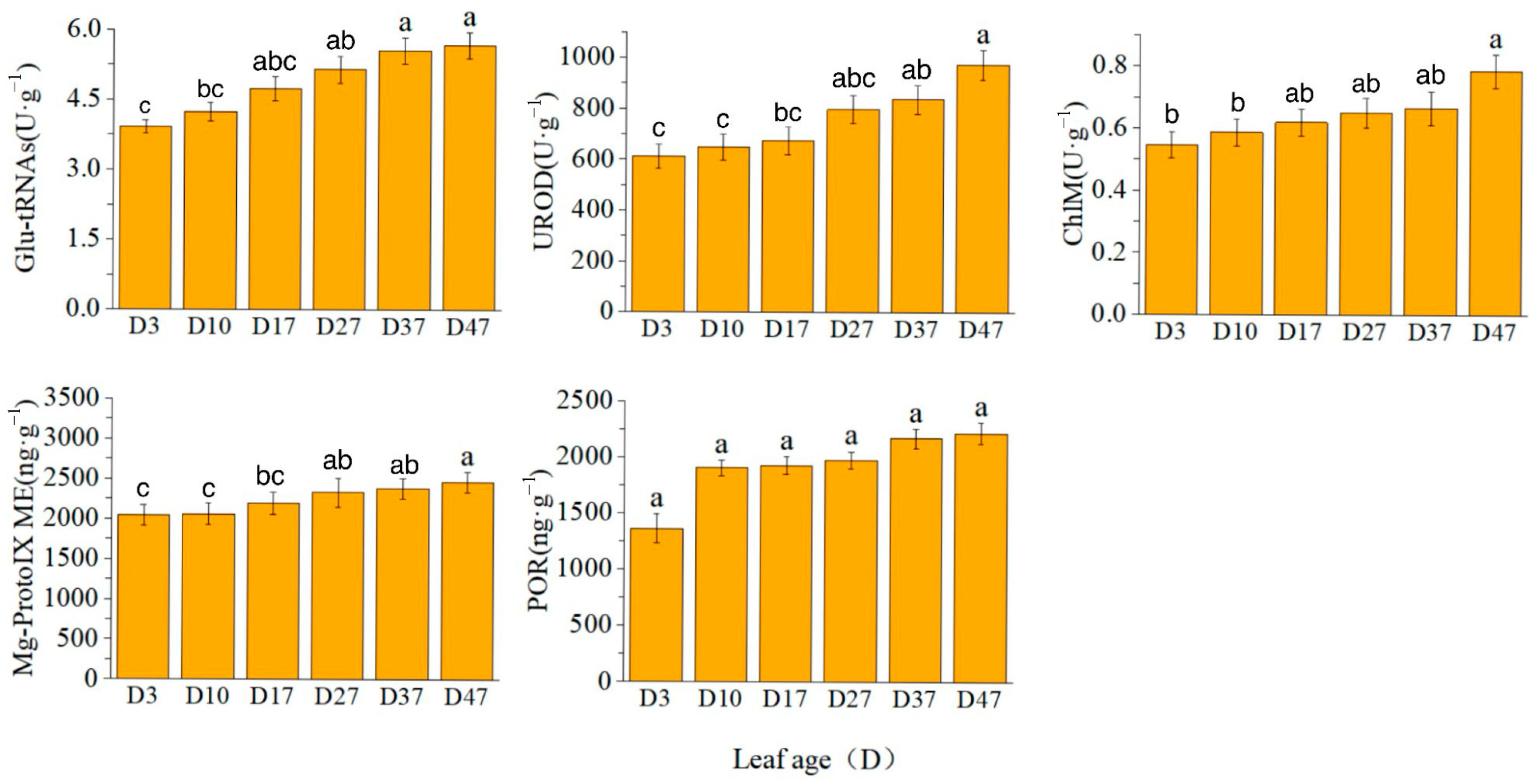
2.1. Chlorophyll Synthesis Precursor Content and Precursor Synthase Activity
2.2. Expression Levels of Key Genes for Chlorophyll Synthesis
2.3. Identification and Basic Physical and Chemical Properties of the CcGLK Proteins
2.4. Phylogeny, Conserved Motifs, and Gene Structure of CcGLKs
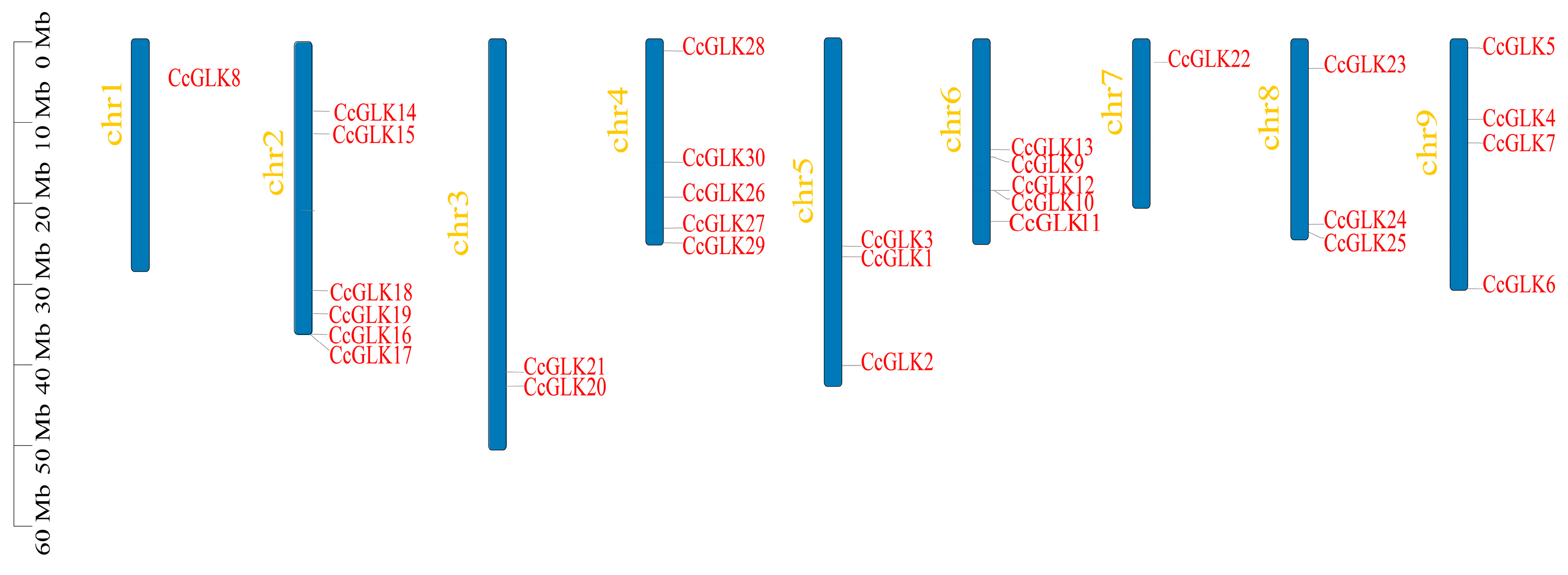
2.5. Chromosome Position, Collinearity Analysis, and Gene Duplication of CcGLKs
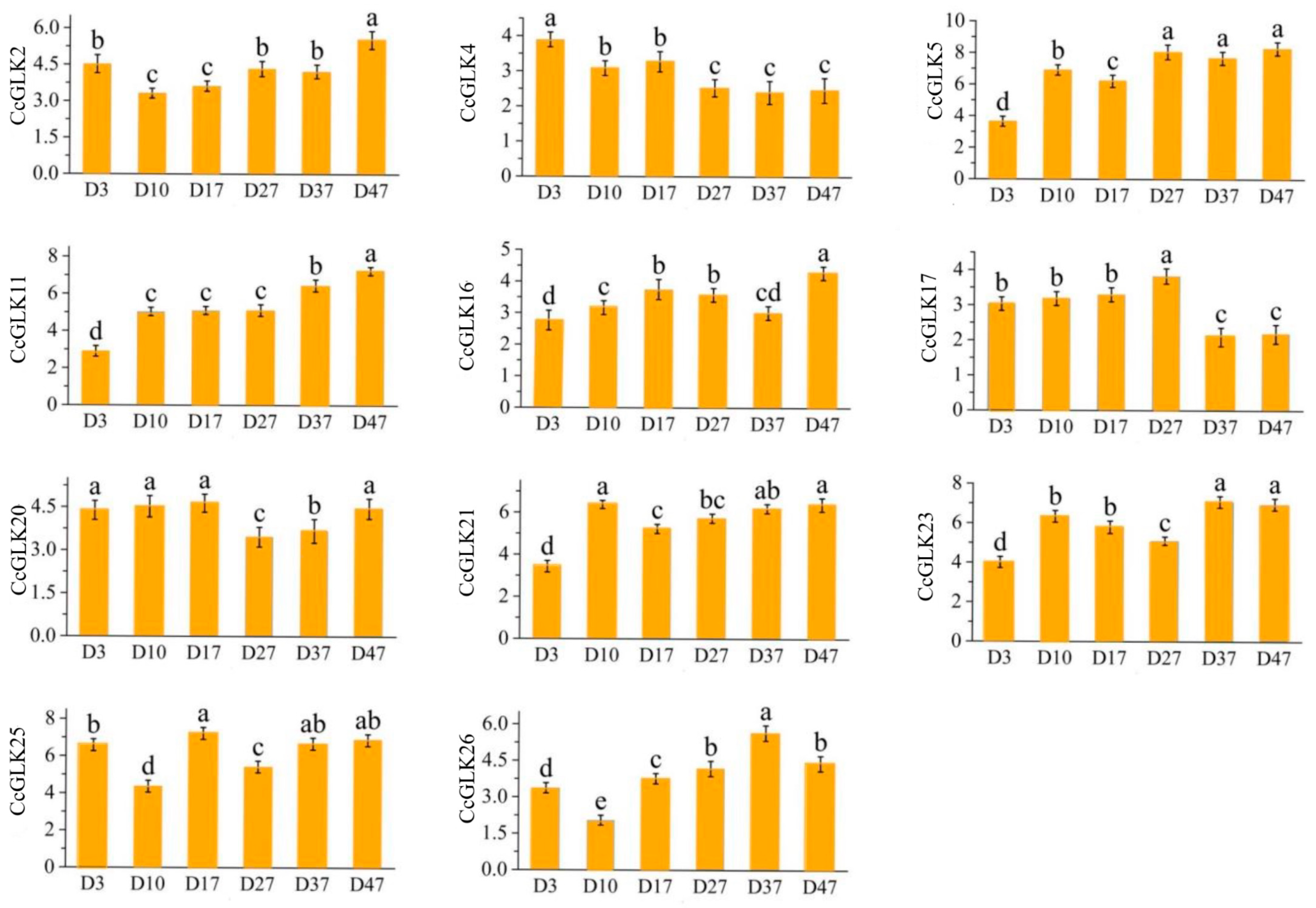
2.6. Citrus GLK Expression Pattern
2.7. Correlation Analysis
3. Materials and Methods
3.1. Test Materials
3.2. GLK Gene Family Members and Prediction of Their Physicochemical Properties
3.3. Chromosomal Distribution, Gene Duplication, and Collinearity Analysis of CcGLKs
3.4. Determination of Chlorophyll Synthesis Precursor Content and Key Precursor Synthases’ Activity
3.5. Characterization, Phylogenetic, and Physicochemical Properties Analysis of CcGLKs
3.6. The Analysis of the Expression Patterns of CcGLKs
3.7. Data Processing and Analysis
4. Discussion
4.1. Differences in Key Precursor Content and Gene Expression of Chlorophyll Synthesis
4.2. Identification and Expression Analysis of GLK Genes in Citrus
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Powell, A.L.T.; Nguyen, C.V.; Hill, T.; Cheng, K.L.; Figueroa-Balderas, R.; Aktas, H.; Ashrafi, H.; Pons, C.; Fernandez-Munoz, R.; Vicente, A.; et al. Uniform ripening Encodes a Golden 2-like Transcription Factor Regulating Tomato Fruit Chloroplast Development. Science 2012, 336, 1711–1715. [Google Scholar] [CrossRef] [PubMed]
- Waters, M.T.; Langdale, J.A. The making of a chloroplast. Embo J. 2009, 28, 2861–2873. [Google Scholar] [CrossRef] [PubMed]
- Yasumura, Y.; Moylan, E.C.; Langdale, J.A. A conserved transcription factor mediates nuclear control of organelle biogenesis in anciently diverged land plants. Plant Cell 2005, 17, 1894–1907. [Google Scholar] [CrossRef] [PubMed]
- Waters, M.T.; Moylan, E.C.; Langdale, J.A. GLK transcription factors regulate chloroplast development in a cell-autonomous manner. Plant J. 2008, 56, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, X.; Shenghui, J.; Binbin, W. Transcriptomic and functional analyses reveal that PpGLK1 Regulates Chloroplast Development in Peach (Prunus persica). Front. Plant Sci. 2018, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Ampomah Dwamena, C.; Thrimawithana, A.H.; Dejnoprat, S.; Lewis, D.; Espley, R.V.; Allan, A.C. A kiwifruit (Actinidia deliciosa) R2R3-MYB transcription factor modulates chlorophyll and carotenoid accumulation. New Phytol. 2019, 221, 309–325. [Google Scholar] [CrossRef]
- Waters, M.T.; Wang, P.; Korkaric, M.; Capper, R.G.; Saunders, N.J.; Langdale, J.A. GLK Transcription Factors Coordinate Expression of the Photosynthetic Apparatus in Arabidopsis. Plant Cell 2009, 21, 1109–1128. [Google Scholar] [CrossRef]
- Pan, Y.; Bradley, G.; Pyke, K.; Ball, G.; Lu, C.; Fray, R.; Marshall, A.; Jayasuta, S.; Baxter, C.; van Wijk, R.; et al. Network Inference Analysis Identifies an APRR2-Like Gene Linked to Pigment Accumulation in Tomato and Pepper Fruits. Plant Physiol. 2013, 161, 1476–1485. [Google Scholar] [CrossRef]
- Adhikari, N.D.; Froehlich, J.E.; Strand, D.D.; Buck, S.M.; Kramer, D.M.; Larkin, R.M. GUN4-Porphyrin complexes bind the ChlH/GUN5 subunit of Mg-Chelatase and promote chlorophyll biosynthesis in arabidopsis. Plant Cell 2011, 23, 1449–1467. [Google Scholar] [CrossRef]
- Hall, L.N.; Rossini, L.; Cribb, L.; Langdale, J.A. GOLDEN 2: A novel transcriptional regulator of cellular differentiation in the maize leaf. Plant Cell 1998, 10, 925–936. [Google Scholar] [CrossRef]
- Rossini, L.; Cribb, L.; Martin, D.J.; Langdale, J.A. The maize golden2 gene defines a novel class of transcriptional regulators in plants. Plant Cell 2001, 13, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Fouracre, J.; Kelly, S.; Karki, S.; Gowik, U.; Aubry, S.; Shaw, M.K.; Westhoff, P.; Slamet-Loedin, I.H.; Quick, W.P.; et al. Evolution of GOLDEN2-LIKE gene function in C-3 and C-4 plants. Planta 2013, 237, 481–495. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Obayashi, T.; Masuda, T. Role of the G-box element in regulation of chlorophyll biosynthesis in Arabidopsis roots. Plant Signal. Behav. 2012, 7, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Moran, R.; Porath, D. Chlorophyll determination in intact tissues using n,n-dimethylformamide. Plant Physiol. 1980, 65, 478–479. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, J.; Yao, X.; Zhang, Y.; Li, J.; Wang, X.; Xu, Z.; Chen, W. Characterization and fine mapping of thermo-sensitive chlorophyll deficit mutant1 in rice (Oryza sativa L.). Breed Sci. 2015, 65, 161–169. [Google Scholar] [CrossRef]
- Westbeek, M.; Pons, T.L.; Atkin, C. Analysis of differences in photosynthetic nitrogen use efficiency of alpine and lowland Poa species. Oecologia 1999, 120, 19–26. [Google Scholar] [CrossRef]
- Kumar, A.M.; Soll, D. Antisense HEMA1 RNA expression inhibits heme and chlorophyll biosynthesis in arabidopsis. Plant Physiol. 2000, 122, 49–56. [Google Scholar] [CrossRef]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole Metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef]
- Oster, U.; Tanaka, R.; Tanaka, A.; Rudiger, W. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2000, 21, 305–310. [Google Scholar] [CrossRef]
- Rose, A.; Meier, I.; Wienand, U. The tomato I-box binding factor LeMYBI is a member of a novel class of myb-like proteins. Plant J. Cell Mol. Biol. 1999, 20, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef]
- Massari, M.E.; Murre, C. Helix-Loop-Helix Proteins: Regulators of Transcription in Eucaryotic Organisms. Mol. Cell. Biol. 2000, 20, 429–440. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, Y.; Wang, X.; Gu, S.; Yu, J.; Liang, G.; Yan, C.; Xu, C. Genomewide comparative phylogenetic and molecular evolutionary analysis of tubby-like protein family in Arabidopsis, rice, and poplar. Genomics 2008, 92, 246–253. [Google Scholar] [CrossRef]
- Hosoda, K.; Imamura, A.; Katoh, E.; Hatta, T.; Tachiki, M.; Yamada, H.; Mizuno, T.; Yamazaki, T. Molecular structure of the GARP family of plant Myb-related DNA binding motifs of the Arabidopsis response regulators. Plant Cell 2002, 14, 2015–2029. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Han, G.; Zhou, L.; Ali, A.; Zhu, S.; Li, X. Molecular Evolution and Genetic Variation of G2-Like Transcription Factor Genes in Maize. PLoS ONE 2016, 11, e0161763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef]
- Passardi, F.; Longet, D.; Penel, C.; Dunand, C. The class III peroxidase multigenic family in rice and its evolution in land plants. Phytochemistry 2004, 65, 1879–1893. [Google Scholar] [CrossRef] [PubMed]
- Rogozin, I.B.; Carmel, L.; Csuros, M.; Koonin, E.V. Origin and evolution of spliceosomal introns. Biol. Direct 2012, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Zhu, W.; Silva, J.C.; Gu, X.; Buell, C.R. Intron gain and loss in segmentally duplicated genes in rice. Genome Biol. 2006, 7, R41. [Google Scholar] [CrossRef]
- Prince, V.E.; Pickett, F.B. Splitting pairs: The diverging fates of duplicated genes. Nat. Rev. Genet. 2002, 3, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhu, B.; Fu, D.; Luo, Y. RIN transcription factor plays an important role in ethylene biosynthesis of tomato fruit ripening. J. Sci. Food Agric. 2011, 91, 2308–2314. [Google Scholar] [CrossRef]
- Li, G.; Chen, D.; Tang, X.; Liu, Y. Heterologous expression of kiwifruit (Actinidia chinensis) GOLDEN2-LIKE homolog elevates chloroplast level and nutritional quality in tomato (Solanum lycopersicum). Planta 2018, 247, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, F.; Obayashi, T.; Sasaki-Sekimoto, Y.; Ohta, H.; Takamiya, K.-i.; Masuda, T. Gene expression profiling of the tetrapyrrole metabolic pathway in Arabidopsis with a mini-array system. Plant Physiol. 2004, 135, 2379–2391. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, P.G.; Terry, M.J. Light signalling pathways regulating the Mg-chelatase branchpoint of chlorophyll synthesis during de-etiolation in Arabidopsis thaliana. Photochem. Photobiol. Sci. 2008, 7, 1243–1252. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of aa | MolWt | pI | II | GRAVY | Number of Introns |
|---|---|---|---|---|---|---|
| I | 250–496 | 28,464.01–54,724.06 | 5.57–8.84 | 27.46–71.22 | −0.972–−0.476 | 5–8 |
| II | 122–419 | 14,435.59–46,634.53 | 7.23–10.17 | 41.32–62.37 | −1.057–−0.578 | 2–5 |
| III | 189–494 | 20,919.32–54,020.85 | 6.01–10.46 | 43.87–70.95 | −1.032–−0.246 | 0–5 |
| Sequence 1 | Sequence 2 | Ka | Ks | Ka/Ks | Copy Type |
|---|---|---|---|---|---|
| CcGLK15 | CcGLK18 | 0.326772 | 1.449596 | 0.225422539 | tandem duplication |
| CcGLK28 | CcGLK9 | 0.358406 | 1.893917 | 0.189240602 | tandem duplication |
| Chromosomes | Arabidopsis Thaliana | Location | Citrus C Lementina | Location | |
|---|---|---|---|---|---|
| chr1 | AT1G32240.1 | == | chr5 | CcGLK1 | 27065813…27070133 |
| chr5 | AT5G45580.2 | == | chr5 | CcGLK3 | 25793408…25795877 |
| chr1 | AT1G79430.2 | == | chr9 | CcGLK6 | 31030857…31033422 |
| chr3 | AT3G12730.1 | == | chr9 | CcGLK6 | 31030857…31033422 |
| chr2 | AT2G02060.2 | == | chr9 | CcGLK7 | 12119033…12122259 |
| chr2 | AT2G38300.1 | == | chr6 | CcGLK9 | 16820952…16822532 |
| chr2 | AT2G40970.1 | == | chr6 | CcGLK11 | 21920357…21922631 |
| chr5 | AT5G06800.2 | == | chr6 | CcGLK12 | 18766399…18767741 |
| chr2 | AT2G20570.2 | == | chr2 | CcGLK14 | 10399864…10403935 |
| chr5 | AT5G44190.1 | == | chr2 | CcGLK14 | 10399864…10403935 |
| chr2 | AT2G42660.1 | == | chr2 | CcGLK15 | 11280731…11282377 |
| chr2 | AT2G42660.1 | == | chr2 | CcGLK18 | 34240828…34242711 |
| chr4 | AT4G04555.1 | == | chr2 | CcGLK19 | 35451063…35452097 |
| chr2 | AT2G20400.2 | == | chr3 | CcGLK20 | 42990251…42995979 |
| chr3 | AT3G04450.1 | == | chr3 | CcGLK20 | 42990251…42995979 |
| chr4 | AT4G28610.1 | == | chr3 | CcGLK20 | 42990251…42995979 |
| chr3 | AT3G04030.3 | == | chr3 | CcGLK21 | 41238803…41241110 |
| chr5 | AT5G18240.4 | == | chr3 | CcGLK21 | 41238803…41241110 |
| chr1 | AT1G49560.1 | == | chr7 | CcGLK22 | 2881144…2882746 |
| chr4 | AT4G37180.2 | == | chr7 | CcGLK22 | 2881144…2882746 |
| chr1 | AT1G49560.1 | == | chr8 | CcGLK23 | 3625136…3628623 |
| chr2 | AT2G40260.1 | == | chr8 | CcGLK24 | 24682989…24684558 |
| chr3 | AT3G13040.1 | == | chr8 | CcGLK25 | 24976315…24979483 |
| chr1 | AT1G25550.1 | == | chr4 | CcGLK26 | 19605473…19608059 |
| chr1 | AT1G68670.1 | == | chr4 | CcGLK26 | 19605473…19608059 |
| chr1 | AT1G13300.1 | == | chr4 | CcGLK26 | 19605473…19608059 |
| chr3 | AT3G25790.1 | == | chr4 | CcGLK26 | 19605473…19608059 |
| chr1 | AT1G69580.2 | == | chr4 | CcGLK27 | 23957038…23959284 |
| chr2 | AT2G40260.1 | == | chr4 | CcGLK28 | 1461989…1463323 |
| chr2 | AT2G38300.1 | == | chr4 | CcGLK28 | 1461989…1463323 |
| Chromosomes | Oryza sativa | Location | Citrus Clementina | Location | |
|---|---|---|---|---|---|
| chr1 | LOC_Os01g08160.1 | == | chr1 | CcGLK8 | 5067104…5069631 |
| chr1 | LOC_Os01g13740.1 | == | chr2 | CcGLK14 | 10399864…10403935 |
| chr2 | LOC_Os02g14490.1 | == | chr2 | CcGLK18 | 34240828…34242711 |
| chr2 | LOC_Os02g14490.1 | == | chr2 | CcGLK15 | 11280731…11282377 |
| chr6 | LOC_Os06g35140.1 | == | chr2 | CcGLK18 | 34240828…34242711 |
| chr6 | LOC_Os06g35140.1 | == | chr2 | CcGLK15 | 11280731…11282377 |
| chr11 | LOC_Os11g01480.1 | == | chr4 | CcGLK28 | 1461989…1463323 |
| chr12 | LOC_Os12g01490.1 | == | chr4 | CcGLK28 | 1461989…1463323 |
| chr3 | LOC_Os03g20900.1 | == | chr4 | CcGLK27 | 23957038…23959284 |
| chr8 | LOC_Os08g33750.1 | == | chr4 | CcGLK27 | 23957038…23959284 |
| chr3 | LOC_Os03g55760.1 | == | chr5 | CcGLK2 | 40569723..40573126 |
| chr11 | LOC_Os11g01480.1 | == | chr6 | CcGLK9 | 16820952…16822532 |
| chr12 | LOC_Os12g01490.1 | == | chr6 | CcGLK9 | 16820952…16822532 |
| chr5 | LOC_Os05g34110.1 | == | chr6 | CcGLK11 | 21920357…21922631 |
| chr7 | LOC_Os07g02800.3 | == | chr7 | CcGLK22 | 2881144…2882746 |
| chr11 | LOC_Os11g01480.1 | == | chr8 | CcGLK24 | 24682989…24684558 |
| chr12 | LOC_Os12g01490.1 | == | chr8 | CcGLK24 | 24682989…24684558 |
| chr2 | LOC_Os02g07770.1 | == | chr9 | CcGLK6 | 31030857…31033422 |
| Gen Name | Forward Primer | Reverse Primer |
|---|---|---|
| CcGLK2 | F: CCAGGGACACAGAGCAGAAC | R: TGGTGGAGGCTGTTGGTTTT |
| CcGLK4 | F: CCCCGTAATCAAGGTAGT | R: GACCCTGTCTTCTGTAGCAC |
| CcGLK5 | F: ATGACGAAGGCTCGCTGAAA | R: CTGGTGCTTCCCCTCCATTT |
| CcGLK11 | F: CCGGCTACTGACCACTTGTT | R: ACAGGCACATACGGCAAGAA |
| CcGLK16 | F: TGACTCCTCTTCGGATGGGAAA | R: TGCAGTCGTTTTTGCACCTC |
| CcGLK17 | F: CGTCACTCAACTAGGCGGTC | R: GAGTCACCCATTTCTCTCCCC |
| CcGLK20 | F: GGCCAGATTCATCAGAAGGGT | R: GCAAATTTCTTTGTATCTCAAGCTG |
| CcGLK21 | F: GTCATCTGGTGTAGGTCCAGT | R: GCTCTTGTTTGGTTGGGGTC |
| CcGLK23 | F: CCACCTCAGCAACAGACCAA | R: CTGCAAGGCACTGACAAACC |
| CcGLK25 | F: GCAACCAGCCGAGTCAAAAA | R: AGGCGAATCCTTTTCAGGCA |
| CcGLK26 | F: GGGGAAAAGAAGGACCAGCA | R: TGCAATTCAGGCGACCAAGA |
| Actin | F: CATCCCTCAGCACCTTCC | R: CCAACCTTAGCACTTCTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, B.; Chen, H.; Ma, Q.; Yao, J.; Wang, J.; Wu, W.; Liao, L.; Wang, X.; Zhang, M.; He, S.; et al. Genome-Wide Analysis of the GLK Gene Family and Its Expression at Different Leaf Ages in the Citrus Cultivar Kanpei. Plants 2024, 13, 936. https://doi.org/10.3390/plants13070936
Xiong B, Chen H, Ma Q, Yao J, Wang J, Wu W, Liao L, Wang X, Zhang M, He S, et al. Genome-Wide Analysis of the GLK Gene Family and Its Expression at Different Leaf Ages in the Citrus Cultivar Kanpei. Plants. 2024; 13(7):936. https://doi.org/10.3390/plants13070936
Chicago/Turabian StyleXiong, Bo, Hongzhen Chen, Qingqing Ma, Junfei Yao, Jialu Wang, Wenjia Wu, Ling Liao, Xun Wang, Mingfei Zhang, Siya He, and et al. 2024. "Genome-Wide Analysis of the GLK Gene Family and Its Expression at Different Leaf Ages in the Citrus Cultivar Kanpei" Plants 13, no. 7: 936. https://doi.org/10.3390/plants13070936
APA StyleXiong, B., Chen, H., Ma, Q., Yao, J., Wang, J., Wu, W., Liao, L., Wang, X., Zhang, M., He, S., He, J., Sun, G., & Wang, Z. (2024). Genome-Wide Analysis of the GLK Gene Family and Its Expression at Different Leaf Ages in the Citrus Cultivar Kanpei. Plants, 13(7), 936. https://doi.org/10.3390/plants13070936