
Characterization of Browning, Chlorogenic Acid Content, and Polyphenol Oxidase Activity in Different Varietal Types of Eggplant (Solanum melongena) for Improving Visual and Nutritional Quality



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation Conditions
2.2. Fruit Collection and Browning Measurement
2.3. Fruit Processing and Composition Analyses
2.4. Statistical Analysis
3. Results
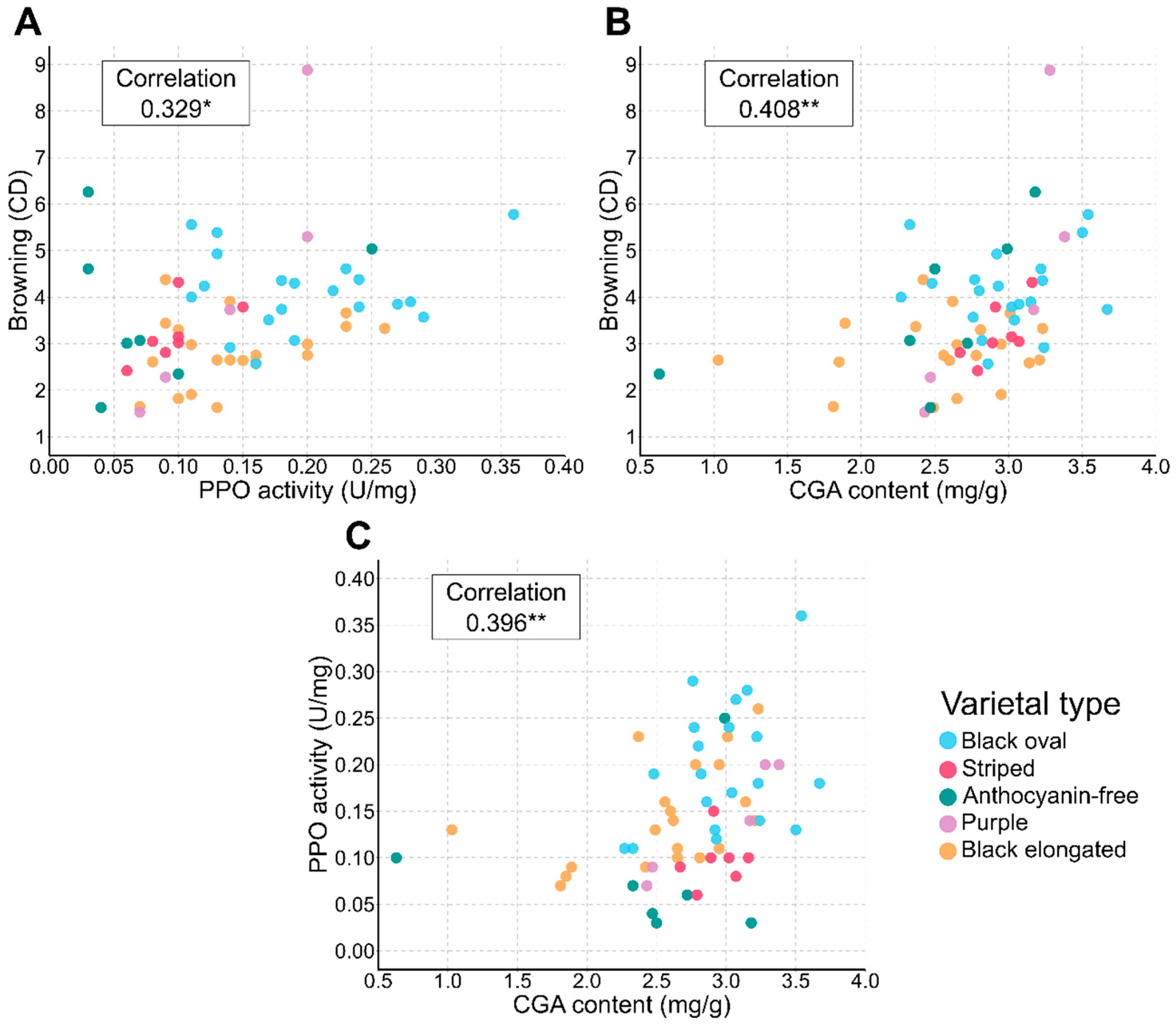
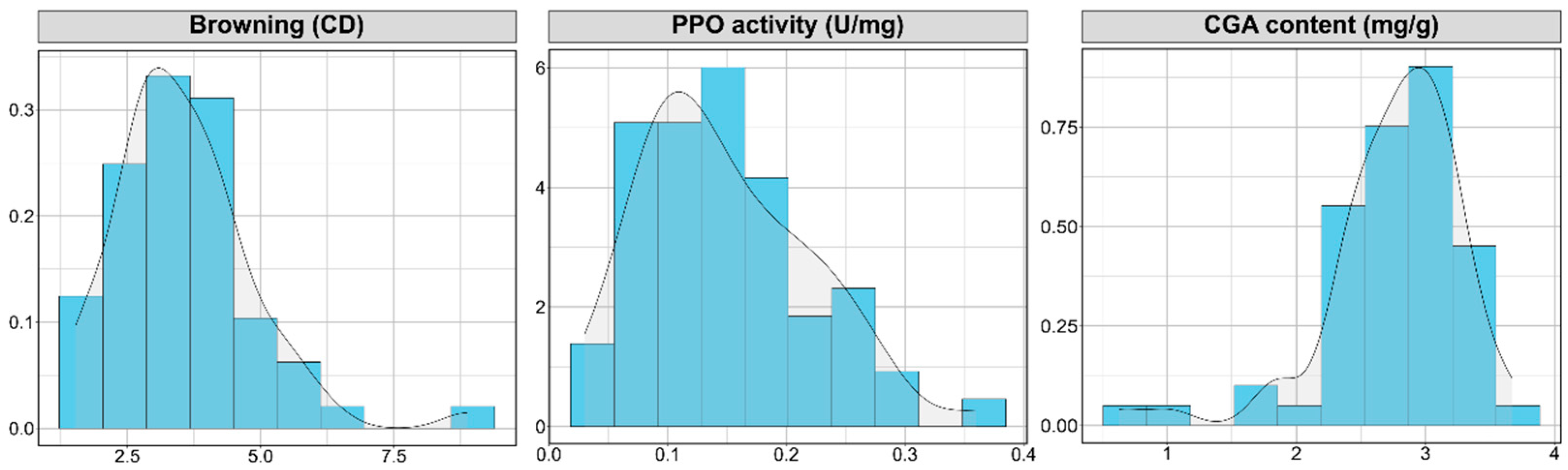
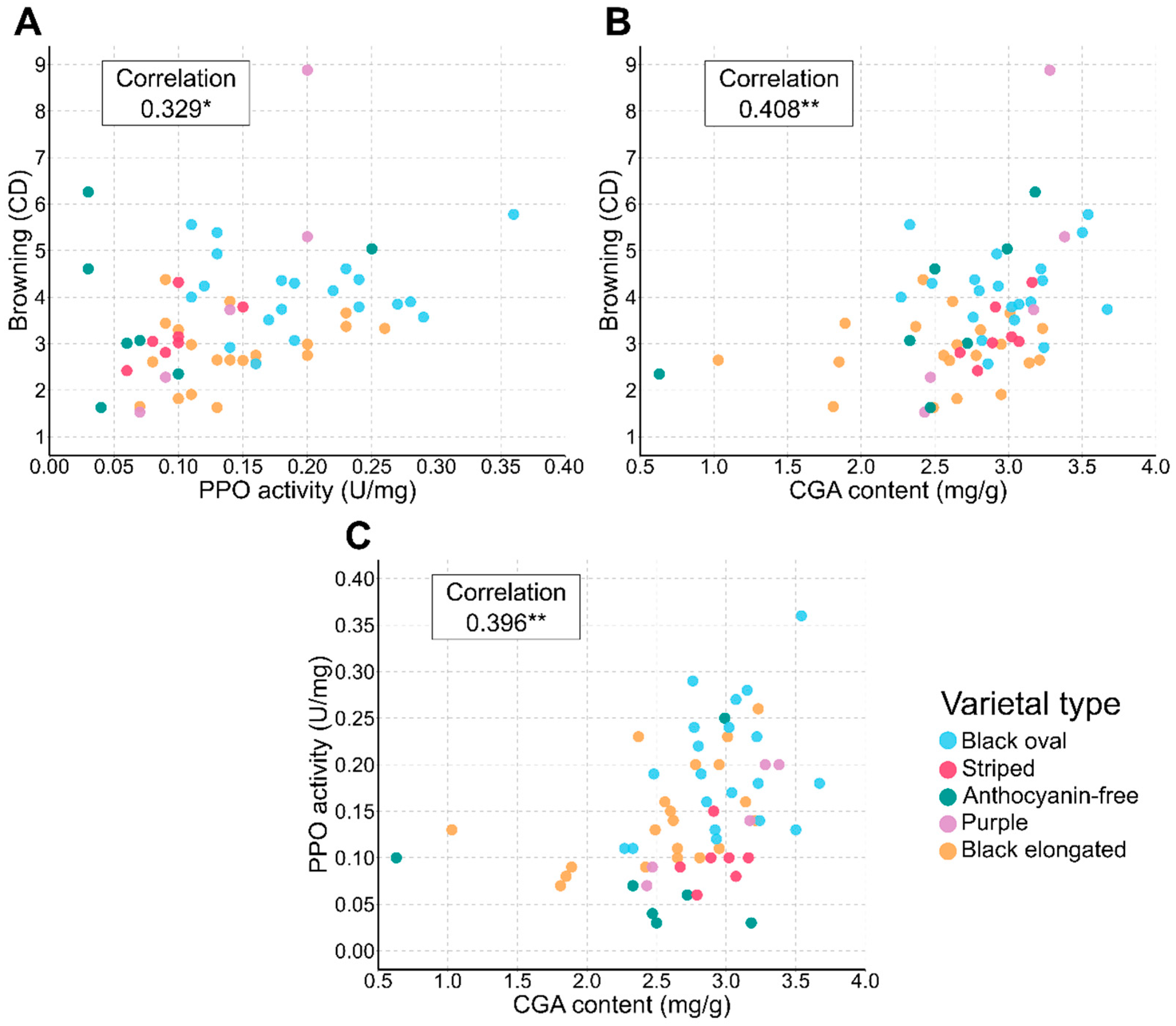
3.1. Distribution Analysis and Characterization of Varietal Types
3.2. Trends in Trait Associations across Varietal Types
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT Database Collections 2022; FAO: Rome, Italy, 2022. [Google Scholar]
- Rotino, G.L.; Sala, T.; Toppino, L. Eggplant. In Alien Gene Transfer in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2014; Volume 2, pp. 381–410. ISBN 978-1-4614-9571-0. [Google Scholar]
- Taher, D.; Solberg, S.Ø.; Prohens, J.; Chou, Y.; Rakha, M.; Wu, T. World vegetable center eggplant collection: Origin, composition, seed dissemination and utilization in breeding. Front. Plant Sci. 2017, 8, 1484. [Google Scholar] [CrossRef] [PubMed]
- Oladosu, Y.; Rafii, M.Y.; Arolu, F.; Chukwu, S.C.; Salisu, M.A.; Olaniyan, B.A.; Fagbohun, I.K.; Muftaudeen, T.K. Genetic diversity and utilization of cultivated eggplant germplasm in varietal improvement. Plants 2021, 10, 1714. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Tiwari, S.K.; Tiwari, K.N. Genetic improvement of eggplant: Perspectives and challenges. In Genetic Engineering of Crop Plants for Food and Health; Springer: Berlin/Heidelberg, Germany, 2023; pp. 123–149. [Google Scholar]
- Gürbüz, N.; Uluişik, S.; Frary, A.; Frary, A.; Doğanlar, S. Health benefits and bioactive compounds of eggplant. Food Chem. 2018, 268, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Stommel, J.R.; Whitaker, B.D. Phenolic Acid Content and composition of eggplant fruit in a germplasm core subset. J. Am. Soc. Hortic. Sci. 2003, 128, 704–710. [Google Scholar] [CrossRef]
- Saha, P.; Singh, J.; Bhanushree, N.; Harisha, S.M.; Tomar, B.S.; Rathinasabapathi, B. Eggplant (Solanum melongena L.) nutritional and health promoting phytochemicals. In Compendium of Crop Genome Designing for Nutraceuticals; Springer Nature Singapore Pte Ltd.: Singapore, 2023; pp. 1463–1493. ISBN 9789811941696. [Google Scholar]
- Plazas, M.; Andújar, I.; Vilanova, S.; Hurtado, M.; Gramazio, P.; Herraiz, F.J.; Prohens, J. Breeding for chlorogenic acid content in eggplant: Interest and prospects. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 26–35. [Google Scholar] [CrossRef]
- Plazas, M.; López-Gresa, M.P.; Vilanova, S.; Torres, C.; Hurtado, M.; Gramazio, P.; Andújar, I.; Herráiz, F.J.; Bellés, J.M.; Prohens, J. Diversity and relationships in key traits for functional and apparent quality in a collection of eggplant: Fruit phenolics content, antioxidant activity, polyphenol oxidase activity, and browning. J. Agric. Food Chem. 2013, 61, 8871–8879. [Google Scholar] [CrossRef]
- Mishra, B.B.; Gautam, S.; Sharma, A. Free phenolics and polyphenol oxidase (PPO): The factors affecting post-cut browning in eggplant (Solanum melongena). Food Chem. 2013, 139, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.M.; Chandrashekar, A.; Venkatesh, Y.P. Eggplant polyphenol oxidase multigene family: Cloning, phylogeny, expression analyses and immunolocalization in response to wounding. Phytochemistry 2011, 72, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, X. Recent advances in polyphenol oxidase-mediated plant stress responses. Phytochemistry 2021, 181, 112588. [Google Scholar] [CrossRef]
- Taranto, F.; Pasqualone, A.; Mangini, G.; Tripodi, P.; Miazzi, M.M.; Pavan, S.; Montemurro, C.; Selles, S.; Burke, A. Polyphenol oxidases in crops: Biochemical, physiological and genetic aspects. Int. J. Mol. Sci. 2017, 18, 377. [Google Scholar] [CrossRef]
- CIE Recommendations on uniform color spaces, color-difference equations, and psychometric color terms. Color Res. Appl. 1977, 2, 5–6. [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, F.A.; Demir, O.; Torun, H.; Kolcuoglu, Y.; Colak, A. Characterization of polyphenoloxidase (PPO) and total phenolic contents in medlar (Mespilus germanica L.) fruit during ripening and over ripening. Food Chem. 2008, 106, 291–298. [Google Scholar] [CrossRef]
- Helmja, K.; Vaher, M.; Püssa, T.; Raudsepp, P.; Kaljurand, M. Evaluation of antioxidative capability of the tomato (Solanum lycopersicum) skin constituents by capillary electrophoresis and high-performance liquid chromatography. Electrophoresis 2008, 29, 3980–3988. [Google Scholar] [CrossRef] [PubMed]
- Luthria, D.L.; Mukhopadhyay, S. Influence of sample preparation on assay of phenolic acids from eggplant. J. Agric. Food Chem. 2006, 54, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Plazas, M.; Prohens, J.; Cuñat, A.N.; Vilanova, S.; Gramazio, P.; Herraiz, F.J.; Andújar, I. Reducing capacity, chlorogenic acid content and biological activity in a collection of scarlet (Solanum aethiopicum) and Gboma (S. macrocarpon) eggplants. Int. J. Mol. Sci. 2014, 15, 17221–17241. [Google Scholar] [CrossRef] [PubMed]
- Schloerke, B.; Cook, D.; Larmarange, J.; Briatte, F.; Marbach, M.; Thoen, E.; Elberg, A.; Crowley, J. GGally: Extension to “ggplot2”. 2024. Available online: https://ggobi.github.io/ggally/ (accessed on 5 April 2024).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Salguero, P. RColorConesa: Conesa Colors Palette. 2022. Available online: https://github.com/ConesaLab/RColorConesa (accessed on 5 April 2024).
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Rosa-Martínez, E.; Bovy, A.; Plazas, M.; Tikunov, Y.; Prohens, J.; Pereira-Dias, L. Genetics and breeding of phenolic content in tomato, eggplant and pepper fruits. Front. Plant Sci. 2023, 14, 1135237. [Google Scholar] [CrossRef]
- Toppino, L.; Barchi, L.; Lo Scalzo, R.; Palazzolo, E.; Francese, G.; Fibiani, M.; D’Alessandro, A.; Papa, V.; Laudicina, V.A.; Sabatino, L.; et al. Mapping quantitative trait loci affecting biochemical and morphological fruit properties in eggplant (Solanum melongena L.). Front. Plant Sci. 2016, 7, 256. [Google Scholar] [CrossRef]
- Docimo, T.; Francese, G.; Ruggiero, A.; Batelli, G.; De Palma, M.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Mennella, G.; Tucci, M. Phenylpropanoids accumulation in eggplant fruit: Characterization of biosynthetic genes and regulation by a MYB transcription factor. Front. Plant Sci. 2016, 6, 1233. [Google Scholar] [CrossRef]
- Ballester, A.R.; Tikunov, Y.; Molthoff, J.; Grandillo, S.; Viquez-Zamora, M.; de Vos, R.; de Maagd, R.A.; van Heusden, S.; Bovy, A.G. Identification of loci affecting accumulation of secondary metabolites in tomato fruit of a Solanum lycopersicum × Solanum chmielewskii introgression line population. Front. Plant Sci. 2016, 7, 1428. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, P.; Gramazio, P.; Vilanova, S.; Raigón, M.D.; Prohens, J.; Plazas, M. Phenolics content, fruit flesh colour and browning in cultivated eggplant, wild relatives and interspecific hybrids and implications for fruit quality breeding. Food Res. Int. 2017, 102, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Mennella, G.; Rotino, G.L.; Fibiani, M.; D’Alessandro, A.; Franceses, G.; Toppino, L.; Cavallanti, F.; Acciarri, N.; Lo Scalzo, R. Characterization of health-related compounds in eggplant (Solanum melongena L.) lines derived from introgression of allied species. J. Agric. Food Chem. 2010, 58, 7597–7603. [Google Scholar] [CrossRef] [PubMed]
- Mennella, G.; Lo Scalzo, R.; Fibiani, M.; DAlessandro, A.; Francese, G.; Toppino, L.; Acciarri, N.; De Almeida, A.E.; Rotino, G.L. Chemical and bioactive quality traits during fruit ripening in eggplant (S. melongena L.) and allied species. J. Agric. Food Chem. 2012, 60, 11821–11831. [Google Scholar] [CrossRef] [PubMed]
- Todaro, A.; Cavallaro, R.; Argento, S.; Branca, F.; Spagna, G. Study and characterization of polyphenol oxidase from eggplant (Solanum melongena L.). J. Agric. Food Chem. 2011, 59, 11244–11248. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, D.; Yang, Z.; Chen, F.; Hu, X.; Xiao, Z.; Ji, J.; Ma, L. Non-enzymatic browning induced by chlorogenic acid quinone mediated catechin oxidation. Food Res. Int. 2022, 156, 111297. [Google Scholar] [CrossRef] [PubMed]
- Croguennec, T. Non-enzymatic browning. In Handbook of Food Science and Technology 1: Food Alteration and Food Quality; Jeantet, R., Croguennec, T., Schuck, P., Brulé, G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 1–248. ISBN 9781119268659. [Google Scholar]
- Lemos, M.L.; Gutiérrez, D.R.; Farías, M.J.; Rodríguez, S.d.C. Effect of UV-C treatments on quality and browning-related enzyme activity of fresh-cut eggplant (Solanum melongena L.) during cold storage. J. Food Process. Preserv. 2022, 46, e16986. [Google Scholar] [CrossRef]
- Uscanga-Sosa, D.P.; Pérez-Gago, M.B.; Gómez-Merino, F.C.; Herrera-Corredor, J.A.; Hernández-Cázares, A.S.; Contreras-Oliva, A. Effect of antioxidants and pH on browning and firmness of minimally processed eggplant. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 79–89. [Google Scholar] [CrossRef]
- Ghidelli, C.; Mateos, M.; Rojas-Argudo, C.; Pérez-Gago, M.B. Effect of antioxidants on enzymatic browning of eggplant extract and fresh-cut tissue. J. Food Process. Preserv. 2014, 38, 1501–1510. [Google Scholar] [CrossRef]
- Šilarová, P.; Boulekbache-Makhlouf, L.; Pellati, F.; Česlová, L. Monitoring of chlorogenic acid and antioxidant capacity of Solanum melongena L. (eggplant) under different heat and storage treatments. Antioxidants 2019, 8, 234. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Trait | Black Oval (n = 20) | Striped (n = 7) | Anthocyanin-Free (n = 7) | Purple (n = 5) | Black Elongated (n = 20) |
|---|---|---|---|---|---|
| Browning (CD) | 4.13 b | 3.22 ab | 3.71 ab | 4.34 ab | 2.85 a |
| (2.57–5.78) | (2.42–4.32) | (1.63–6.26) | (1.53–8.88) | (1.63–4.38) | |
| PPO Activity (U/mg) | 0.20 b | 0.10 a | 0.08 a | 0.14 ab | 0.14 ab |
| (0.11–0.36) | (0.06–0.15) | (0.03–0.25) | (0.07–0.20) | (0.07–0.26) | |
| CGA content (mg/g) | 2.98 | 2.93 | 2.40 | 2.95 | 2.55 |
| (2.27–3.67) | (2.67–3.16) | (0.63–3.18) | (2.43–3.38) | (1.03–3.23) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villanueva, G.; Vilanova, S.; Plazas, M. Characterization of Browning, Chlorogenic Acid Content, and Polyphenol Oxidase Activity in Different Varietal Types of Eggplant (Solanum melongena) for Improving Visual and Nutritional Quality. Plants 2024, 13, 1059. https://doi.org/10.3390/plants13081059
Villanueva G, Vilanova S, Plazas M. Characterization of Browning, Chlorogenic Acid Content, and Polyphenol Oxidase Activity in Different Varietal Types of Eggplant (Solanum melongena) for Improving Visual and Nutritional Quality. Plants. 2024; 13(8):1059. https://doi.org/10.3390/plants13081059
Chicago/Turabian StyleVillanueva, Gloria, Santiago Vilanova, and Mariola Plazas. 2024. "Characterization of Browning, Chlorogenic Acid Content, and Polyphenol Oxidase Activity in Different Varietal Types of Eggplant (Solanum melongena) for Improving Visual and Nutritional Quality" Plants 13, no. 8: 1059. https://doi.org/10.3390/plants13081059
APA StyleVillanueva, G., Vilanova, S., & Plazas, M. (2024). Characterization of Browning, Chlorogenic Acid Content, and Polyphenol Oxidase Activity in Different Varietal Types of Eggplant (Solanum melongena) for Improving Visual and Nutritional Quality. Plants, 13(8), 1059. https://doi.org/10.3390/plants13081059