
Effect of Elicitor Treatments on Quality Attributes in Blueberry: Implications of Cultivar and Environmental Conditions

Abstract
:1. Introduction
2. Results
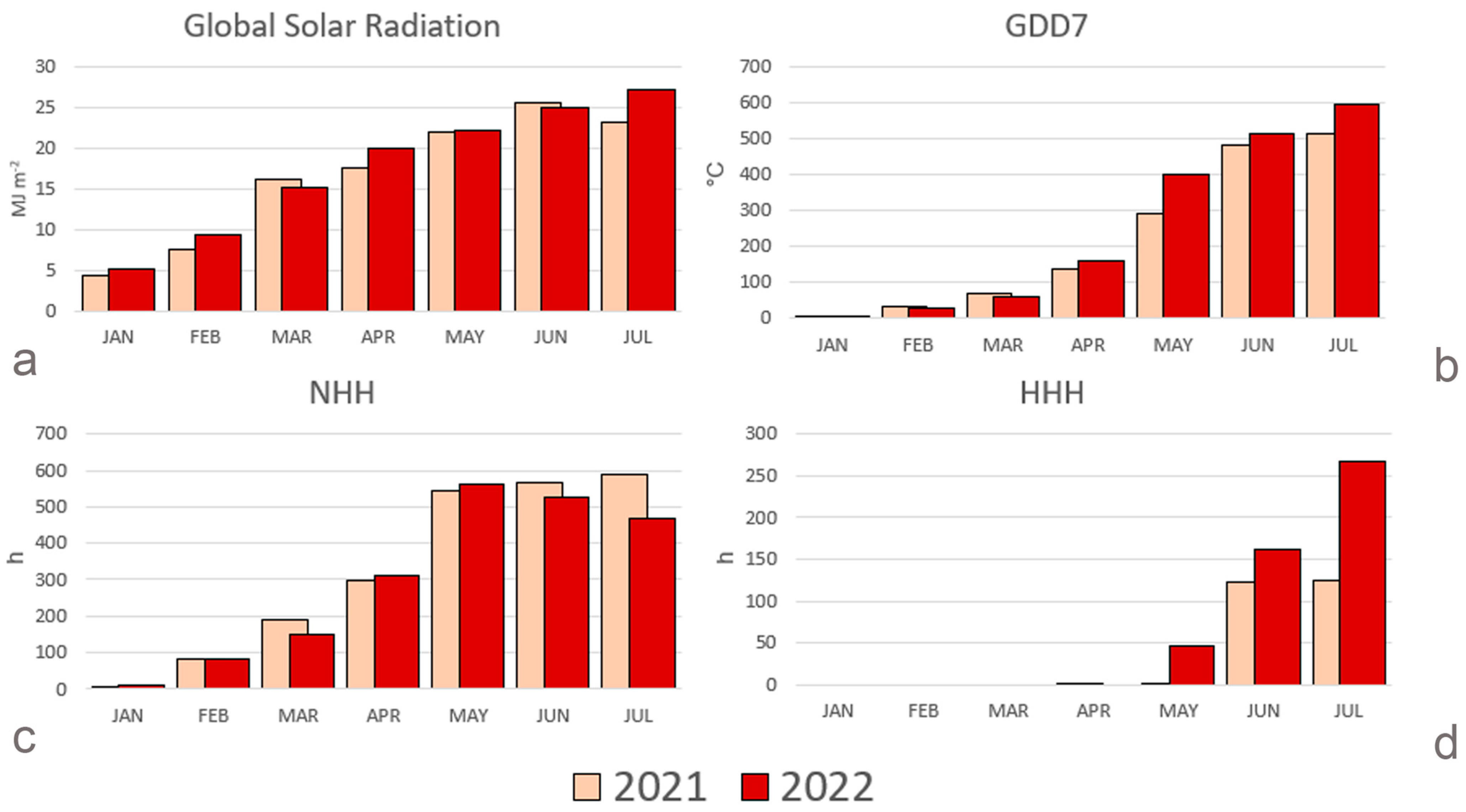
2.1. Environmental Factors
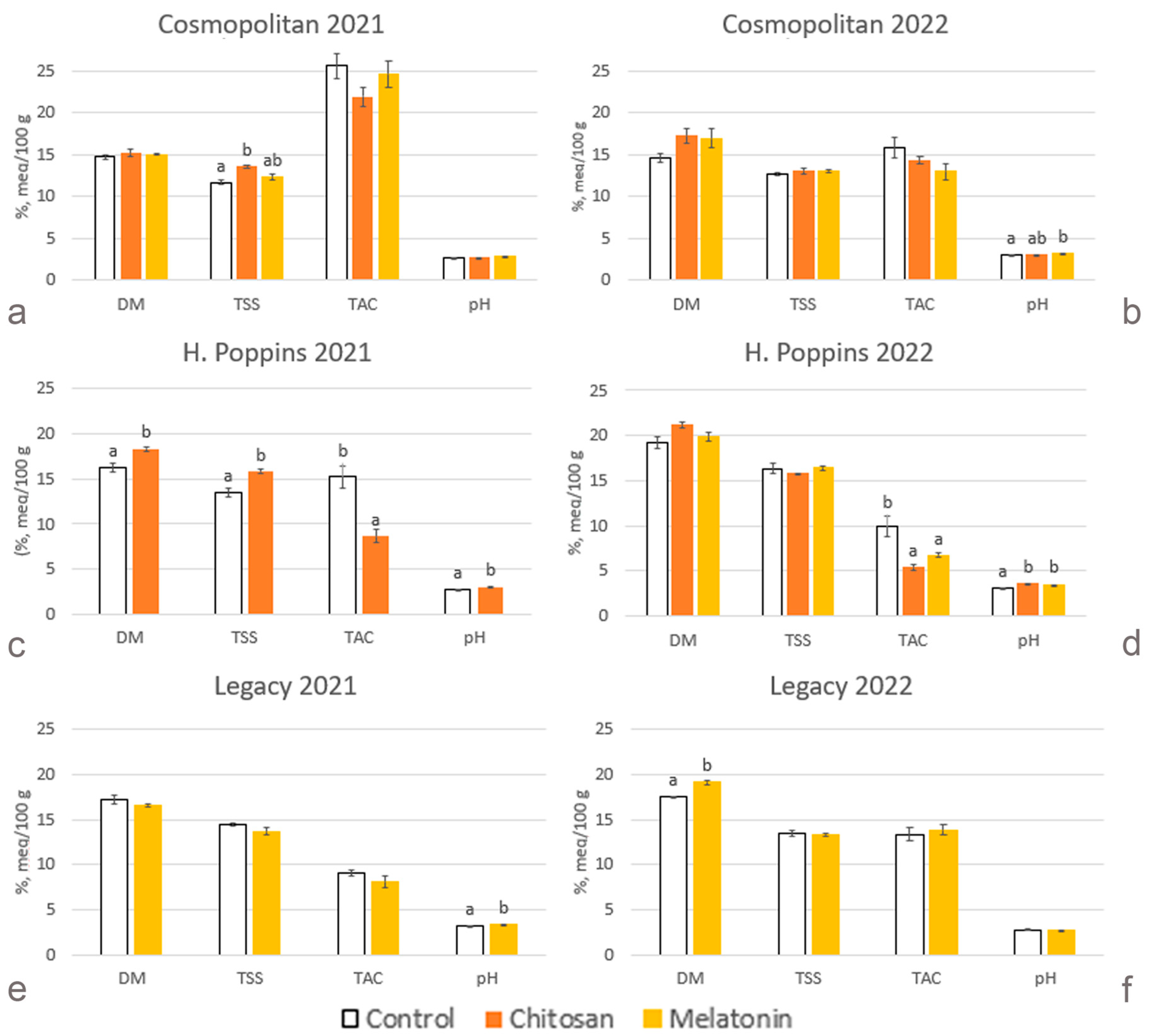
2.2. Fruit Quality Parameters
2.2.1. Dry Matter
2.2.2. Total Soluble Solids
2.2.3. Total Titratable Acidity
2.2.4. pH
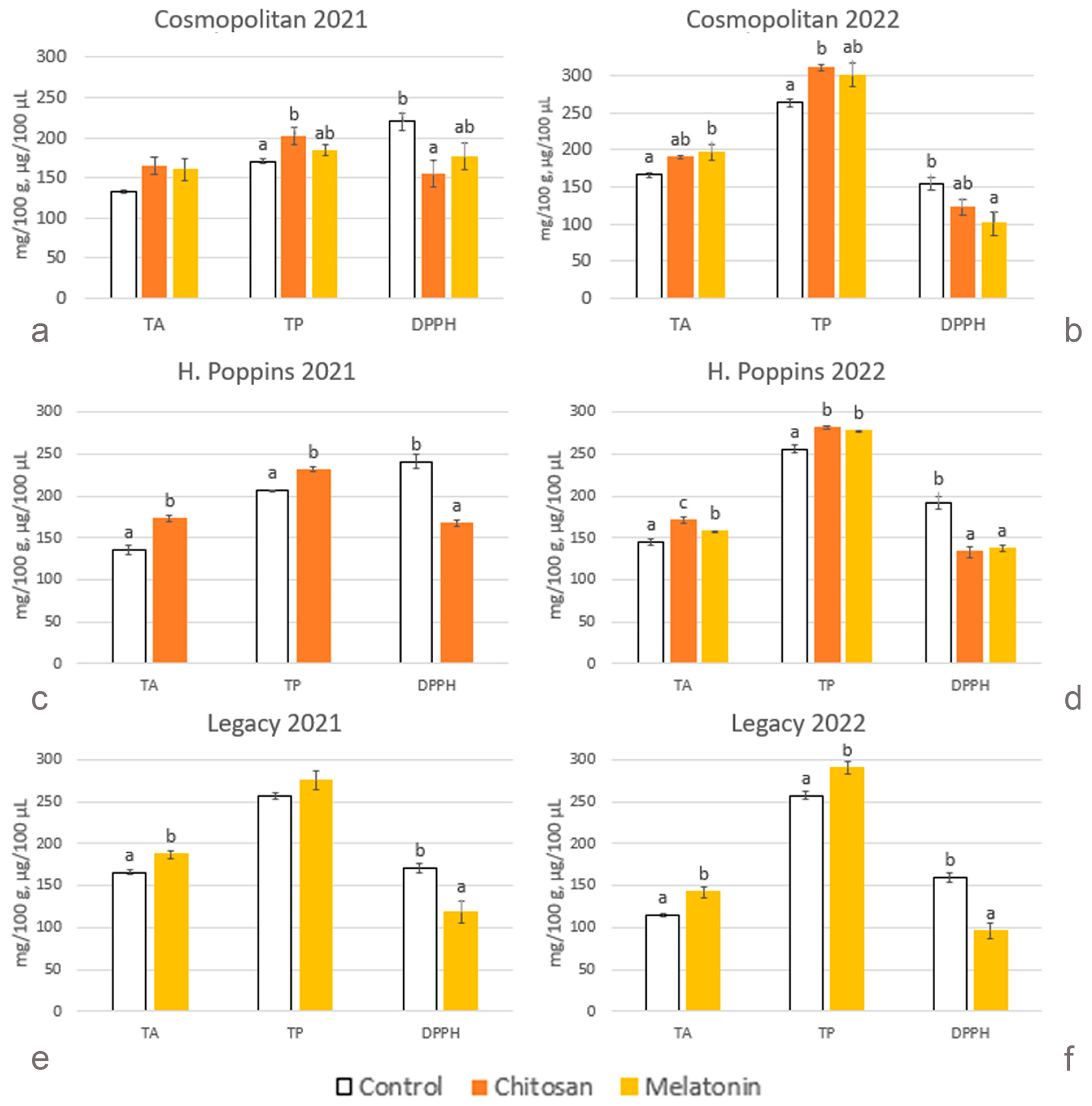
2.2.5. Total Phenolics
2.2.6. Total Anthocyanins
2.2.7. Antioxidant Activity
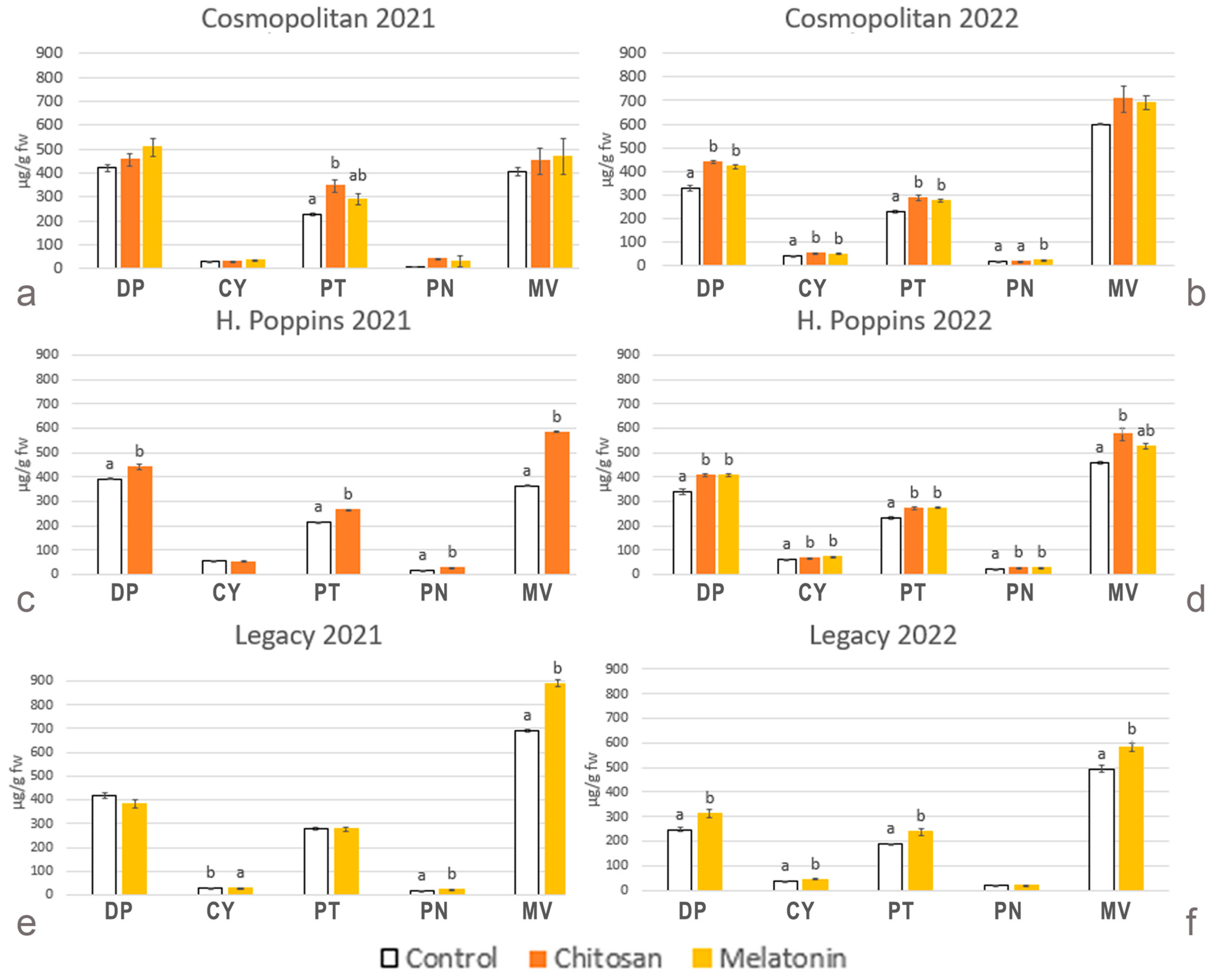
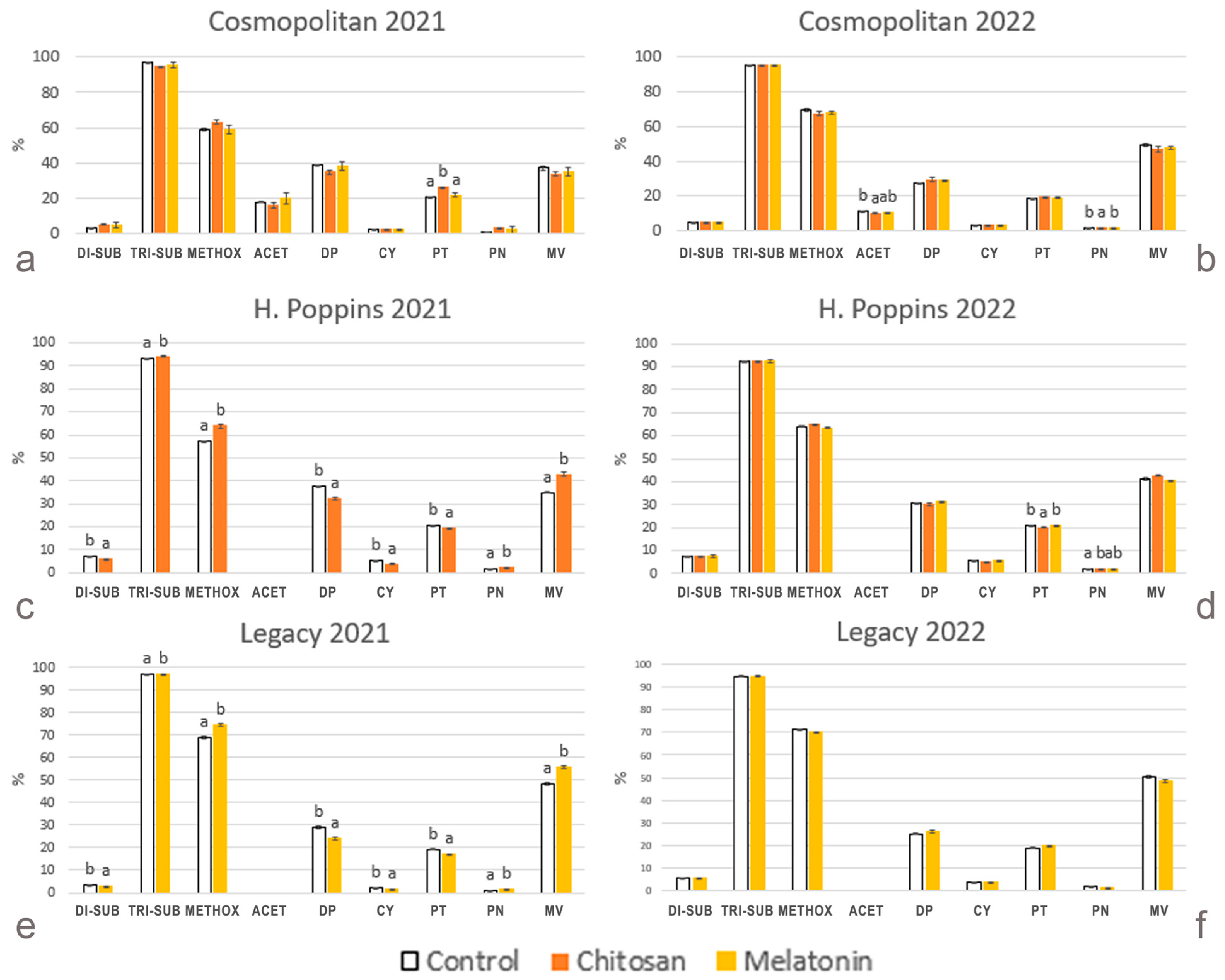
2.2.8. Anthocyanin Profile
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Treatments
4.3. Thermal and Radiative Analysis
- NAS—Strong negative anomaly: T < AVG − 2SD
- NAM—Mild negative anomaly: AVG − 2SD ≤ T < AVG − 1SD
- NORM—No anomaly: AVG − 2SD ≤ T < AVG − 1SD
- PAM—Mild positive anomaly: AVG − 1SD ≤ T < AVG + 1SD
- PAS—Strong positive anomaly: T ≥ AVG + 2SD
- GSR—Summation of daily global solar radiation [MJ m−2]
- GDD7—Accumulation of growing degree days with base temperature 7 °C [79] [GDD]. GDD represents the thermal resources that are useful for the development of blueberry. However, the absence of upper thresholds leads this index to the overestimation of the positive effect of high temperatures.
4.4. Fruit Quality Parameters
4.4.1. Dry Matter Content
4.4.2. Total Soluble Solids
4.4.3. Total Titratable Acidity and pH
4.4.4. Total Phenolics and Total Anthocyanins Analysis
4.4.5. Antioxidant Activity
4.4.6. Anthocyanin Determination by UHPLC-DAD-HR-MS
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abdul Malik, N.A.; Kumar, I.S.; Nadarajah, K. Elicitor and receptor molecules: Orchestrators of plant defense and immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef]
- Reglinski, T.; Havis, N.; Rees, H.J.; de Jong, H. The practical role of induced resistance for crop protection. Phytopathology® 2023, 113, 719–731. [Google Scholar] [CrossRef]
- Palou, L.; Ali, A.; Fallik, E.; Romanazzi, G. GRAS, plant-and animal-derived compounds as alternatives to conventional fungicides for the control of postharvest diseases of fresh horticultural produce. Postharvest. Biol. Technol. 2016, 122, 41–52. [Google Scholar] [CrossRef]
- Desmedt, W.; Kudjordjie, E.N.; Chavan, S.N.; Desmet, S.; Nicolaisen, M.; Vanholme, B.; Vestergård, M.; Kyndt, T. Distinct chemical resistance-inducing stimuli result in common transcriptional, metabolic, and nematode community signatures in rice root and rhizosphere. J. Exp. Bot. 2022, 73, 7564–7581. [Google Scholar] [CrossRef] [PubMed]
- Desmedt, W.; Vanholme, B.; Kyndt, T. Plant defense priming in the field: A review. In Recent Highlights in the Discovery and Optimization of Crop Protection Products; Academic Press: Cambridge, MA, USA, 2021; pp. 87–124. [Google Scholar]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing plant defense priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, V.P.; Brestič, M.; Deshmukh, R.; Vaculik, M. Priming-mediated abiotic stress management in plants: Recent avenues and future directions. Plant Stress 2022, 5, 100097. [Google Scholar] [CrossRef]
- Wisniewski, M.; Droby, S.; Norelli, J.; Liu, J.; Schena, L. Alternative management technologies for postharvest disease control: The journey from simplicity to complexity. Postharvest. Biol. Technol. 2016, 122, 3–10. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, W. UV treatment improved the quality of postharvest fruits and vegetables by inducing resistance. Trends Food Sci. Technol. 2019, 92, 71–80. [Google Scholar] [CrossRef]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- La Camera, S.; Gouzerh, G.; Dhondt, S.; Hoffmann, L.; Fritig, B.; Legrand, M.; Heitz, T. Metabolic reprogramming in plant innate immunity: The contributions of phenylpropanoid and oxylipin pathways. Immunol. Rev. 2004, 198, 267–284. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef]
- Desmedt, W.; Jonckheere, W.; Nguyen, V.H.; Ameye, M.; De Zutter, N.; De Kock, K.; Debode, J.; Van Leeuwen, T.; Audenaert, K.; Vanholme, B.; et al. The phenylpropanoid pathway inhibitor piperonylic acid induces broad-spectrum pest and disease resistance in plants. Plant Cell Environ. 2021, 44, 3122–3139. [Google Scholar] [CrossRef]
- Ruiz-García, Y.; Gómez-Plaza, E. Elicitors: A tool for improving fruit phenolic content. Agriculture 2013, 3, 33–52. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent research on the health benefits of blueberries and their anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef]
- Silva, S.; Costa, E.M.; Veiga, M.; Morais, R.M.; Calhau, C.; Pintado, M. Health promoting properties of blueberries: A review. Crit. Rev. Food Sci. 2020, 60, 181–200. [Google Scholar] [CrossRef]
- Rabea, E.I.; Badawy, M.E.T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Li, P. Chitin and chitosan fragments responsible for plant elicitor and growth stimulator. J. Agric. Food Chem. 2020, 68, 12203–12211. [Google Scholar] [CrossRef]
- Aranaz, I.; Acosta, N.; Civera, C.; Elorza, B.; Mingo, J.; Castro, C.; De los Llanos Gandìa, M.; Heras Caballero, A. Cosmetics and cosmeceutical applications of chitin, chitosan and their derivatives. Polymers 2018, 10, 213. [Google Scholar] [CrossRef]
- Swiatkiewicz, S.; Swiatkiewicz, M.; Arczewska-Wlosek, A.; Jozefiak, D. Chitosan and its oligosaccharide derivatives (chito-oligosaccharides) as feed supplements in poultry and swine nutrition. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1–12. [Google Scholar] [CrossRef]
- Ferri, M.; Tassoni, A. Chitosan as elicitor of health beneficial secondary metabolites in in vitro plant cell cultures. In Handbook of Chitosan Research and Applications; Mackay, R.G., Tait, J.M., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 389–414. [Google Scholar]
- Coelho, N.; Romano, A. Impact of chitosan on plant tissue culture: Recent applications. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 148, 1–13. [Google Scholar] [CrossRef]
- Gutiérrez-Martínez, P.; Bautista-Baños, S.; Berúmen-Varela, G.; Ramos-Guerrero, A.; Hernández-Ibañez, A.M. In vitro response of Colletotrichum to chitosan. Effect on incidence and quality on tropical fruit. Enzymatic expression in mango. Acta Agronómica 2017, 66, 282–289. [Google Scholar] [CrossRef]
- Bautista-Baños, S.; Hernandez-Lauzardo, A.N.; Velazquez-Del Valle, M.G.; Hernández-López, M.; Barka, E.A.; Bosquez-Molina, E.; Wilson, C.L. Chitosan as a potential natural compound to control pre and postharvest diseases of horticultural commodities. Crop Prot. 2006, 25, 108–118. [Google Scholar] [CrossRef]
- Gutiérrez-Martínez, P.; Ramos-Guerrero, A.; Rodríguez-Pereida, C.; Coronado-Partida, L.; Angulo-Parra, J.; González-Estrada, R. Chitosan for postharvest disinfection of fruits and vegetables. In Postharvest Disinfection of Fruits and Vegetables; Academic Press: Cambridge, MA, USA, 2018; pp. 231–241. [Google Scholar]
- Xoca-Orozco, L.Á.; Cuellar-Torres, E.A.; González-Morales, S.; Gutiérrez-Martínez, P.; López-García, U.; Herrera-Estrella, L.; Vega-Arreguìn, J.; Chacón-López, A. Transcriptomic analysis of avocado hass (Persea americana Mill) in the interaction system fruit-chitosan-Colletotrichum. Front. Plant Sci. 2017, 8, 956. [Google Scholar] [CrossRef]
- Yan, J.Q.; Cao, J.K.; Jiang, W.B.; Zhao, Y.M. Effects of preharvest oligochitosan sprays on portharvest fungal disease, storage quality, and defense responses in jujube (Zizyphus jujuba Mill. cv. Dongzao) fruit. Sci. Hortic. 2012, 142, 196–204. [Google Scholar] [CrossRef]
- Romanazzi, G.; Nigro, E.; Ippolito, A.; Di Venere, D. Effects of pre- and postharvest chitosan treatments to control storage grey mold of table grapes. J. Food Sci. 2002, 67, 1862–1867. [Google Scholar] [CrossRef]
- Dos Santos, N.S.T.; Athayde Aguiar, A.J.A.; de Oliveira, C.E.V.; Verissimo de Sales, C.; de Melo E Silva, S.; Sousa da Silva, R.; Stamford, T.C.M.; de Souza, E.L. Efficacy of the application of a coating composed of chitosan and Origanum vulgare L. essential oil to control Rhizopus stolonifer and Aspergillus niger in grapes (Vitis labrusca L.). Food Microbiol. 2012, 32, 345–353. [Google Scholar] [CrossRef]
- Felipini, R.B.; Boneti, J.I.; Katsurayama, Y.; Neto, A.C.R.; Veleirinho, B.; Maraschin, M.; Di Piero, R.M. Apple scab control and activation of plant defence responses using potassium phosphite and chitosan. Eur. J. Plant Pathol. 2016, 145, 929–939. [Google Scholar] [CrossRef]
- Giacalone, G.; Chiabrando, V. Effect of preharvest and postharvest application of chitosan coating on storage quality of nectarines. Acta Hortic. 2015, 1084, 675–680. [Google Scholar] [CrossRef]
- Gayed, A.A.N.A.; Shaarawi, S.A.M.A.; Elkhishen, M.A.; Elsherbini, N.R.M. Pre-harvest application of calcium chloride and chitosan on fruit quality and storability of ‘Early Swelling’peach during cold storage. Cienc. Agrotec. 2017, 41, 220–231. [Google Scholar] [CrossRef]
- Feliziani, E.; Santini, M.; Landi, L.; Romanazzi, G. Pre-and postharvest treatment with alternatives to synthetic fungicides to control postharvest decay of sweet cherry. Postharvest. Biol. Technol. 2013, 78, 133–138. [Google Scholar] [CrossRef]
- Ahmed, Z.F.; Alblooshi, S.S.; Kaur, N.; Maqsood, S.; Schmeda-Hirschmann, G. Synergistic effect of preharvest spray application of natural elicitors on storage life and bioactive compounds of date palm (Phoenix dactylifera L.; cv. Khesab). Horticulturae 2021, 7, 145. [Google Scholar] [CrossRef]
- Cui, K.; Shu, C.; Zhao, H.; Fan, X.; Cao, J.; Jiang, W. Preharvest chitosan oligochitosan and salicylic acid treatments enhance phenol metabolism and maintain the postharvest quality of apricots (Prunus armeniaca L.). Sci. Hortic. 2020, 267, 109334. [Google Scholar] [CrossRef]
- Migliori, C.A.; Salvati, L.; Di Cesare, L.F.; Scalzo, R.L.; Parisi, M. Effects of preharvest applications of natural antimicrobial products on tomato fruit decay and quality during long-term storage. Sci. Hortic. 2017, 222, 193–202. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, H.; Wang, T.; Mustafa, G.; Liu, L.; Wang, Q.; Shao, Z. Quality improvement of tomato fruits by preharvest application of chitosan oligosaccharide. Horticulturae 2023, 9, 300. [Google Scholar] [CrossRef]
- Li, Z.; Xue, S.; Xu, X.; Wang, B.; Zheng, X.; Li, B.; Xie, P.; Bi, Y.; Prusky, D. Preharvest multiple sprays with chitosan accelerate the deposition of suberin poly phenolic at wound sites of harvested muskmelons. Postharvest. Biol. Technol. 2021, 179, 111565. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2021, 63, 577–597. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xing, Q.; Ahammed, G.J.; Zhou, J. Functions and prospects of melatonin in plant growth, yield, and quality. J. Exp. Bot. 2022, 73, 5928–5946. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic stress in crop species: Improving tolerance by applying plant metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the defense mechanisms during plant oxidative stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef]
- Sun, H.; Cao, X.; Wang, X.; Zhang, W.; Li, W.; Wang, X.; Liu, S.; Lyu, D. RBOH-dependent hydrogen peroxide signaling mediates melatonin-induced anthocyanin biosynthesis in red pear fruit. Plant Sci. 2021, 313, 111093. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.F.; Yu, Y.; Shi, T.C.; Fu, Y.S.; Zhao, T.; Zhang, Z.W. Melatonin treatment of pre-veraison grape berries modifies phenolic components and antioxidant activity of grapes and wine. Food Sci. Technol. 2018, 39, 35–42. [Google Scholar] [CrossRef]
- Sun, H.L.; Wang, X.Y.; Shang, Y.; Wang, X.Q.; Du, G.D.; Lü, D.G. Preharvest application of melatonin induces anthocyanin accumulation and related gene upregulation in red pear (Pyrus ussuriensis). J. Integr. Agric. 2021, 20, 2126–2137. [Google Scholar] [CrossRef]
- Michailidis, M.; Tanou, G.; Sarrou, E.; Karagiannis, E.; Ganopoulos, I.; Martens, S.; Molassiotis, A. Pre- and post-harvest melatonin application boosted phenolic compounds accumulation and altered respiratory characters in sweet cherry fruit. Front. Nutr. 2021, 8, 695061. [Google Scholar] [CrossRef] [PubMed]
- Carrión-Antolí, A.; Lorente-Mento, J.M.; Valverde, J.M.; Castillo, S.; Valero, D.; Serrano, M. Effects of melatonin treatment on sweet cherry tree yield and fruit quality. Agronomy 2021, 12, 3. [Google Scholar] [CrossRef]
- Fekry, W.M.; Rashad, Y.M.; Alaraidh, I.A.; Mehany, T. Exogenous application of melatonin and methyl jasmonate as a pre-harvest treatment enhances growth of barhi date palm trees, prolongs storability, and maintains quality of their fruits under storage conditions. Plants 2021, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Medina-Santamarina, J.; Zapata, P.J.; Valverde, J.M.; Valero, D.; Serrano, M.; Guillén, F. Melatonin treatment of apricot trees leads to maintenance of fruit quality attributes during storage at chilling and non-chilling temperatures. Agronomy 2021, 11, 917. [Google Scholar] [CrossRef]
- Medina-Santamarina, J.; Serrano, M.; Lorente-Mento, J.M.; García-Pastor, M.E.; Zapata, P.J.; Valero, D.; Guillén, F. Melatonin treatment of pomegranate trees increases crop yield and quality parameters at harvest and during storage. Agronomy 2021, 11, 861. [Google Scholar] [CrossRef]
- Lorente-Mento, J.M.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valverde, J.M.; Valero, D.; Serrano, M. Melatonin treatment to pomegranate trees enhances fruit bioactive compounds and quality traits at harvest and during postharvest storage. Antioxidants 2021, 10, 820. [Google Scholar] [CrossRef]
- Cortés-Montaña, D.; Bernalte-García, M.J.; Serradilla, M.J.; Velardo-Micharet, B. Optimal Preharvest Melatonin Applications to Enhance Endogenous Melatonin Content, Harvest and Postharvest Quality of Japanese Plum. Agriculture 2023, 13, 1318. [Google Scholar] [CrossRef]
- Song, L.; Tan, Z.; Zhang, W.; Li, Q.; Jiang, Z.; Shen, S.; Luo, S.; Chen, X. Exogenous melatonin improves the chilling tolerance and preharvest fruit shelf life in eggplant by affecting ROS-and senescence-related processes. Hortic. Plant J. 2023, 9, 523–540. [Google Scholar] [CrossRef]
- Li, S.; Cheng, Y.; Yan, R.; Liu, Y.; Huan, C.; Zheng, X. Preharvest spray with melatonin improves postharvest disease resistance in cherry tomato fruit. Postharvest. Biol Technol. 2022, 193, 112055. [Google Scholar] [CrossRef]
- Mariani, L.; Parisi, S.G.; Cola, G.; Failla, O. Climate change in Europe and effects on thermal resources for crops. Int. J. Biometeorol. 2012, 56, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Camacho, J.E.; Guevara-González, R.G.; Rico-García, E.; Tovar-Pérez, E.G.; Torres-Pacheco, I. Delayed senescence and marketability index preservation of blackberry fruit by preharvest application of chitosan and salicylic acid. Front. Plant Sci. 2022, 13, 796393. [Google Scholar] [CrossRef] [PubMed]
- Chiabrando, V.; Giacalone, G. Quality evaluation of blueberries coated with chitosan and sodium alginate during postharvest storage. Int. Food Res. J. 2017, 24, 1553–1561. [Google Scholar]
- Ruiz-García, Y.; Romero-Cascales, I.; Bautista-Ortín, A.B.; Gil-Muñoz, R.; Martínez-Cutillas, A.; Gómez-Plaza, E. Increasing bioactive phenolic compounds in grapes: Response of six monastrell grape clones to benzothiadiazole and methyl jasmonate treatments. Am. J. Enol. Viticult. 2013, 64, 459–465. [Google Scholar] [CrossRef]
- Pan, Y.G.; Liu, X.H. Effect of benzo-thiadiazole-7-carbothioic acid S-methyl ester (BTH) treatment on the resistant substance in postharvest mango fruits of different varieties. Afr. J. Biotechnol. 2011, 10, 15521. [Google Scholar] [CrossRef]
- Smart, L.E.; Martin, J.L.; Limpalaër, M.; Bruce, T.J.; Pickett, J.A. Responses of herbivore and predatory mites to tomato plants exposed to jasmonic acid seed treatment. J. Chem. Ecol. 2013, 39, 1297–1300. [Google Scholar] [CrossRef]
- Jiang, B.; Fang, X.; Fu, D.; Wu, W.; Han, Y.; Chen, H.; Liu, R.; Gao, H. Exogenous salicylic acid regulates organic acids metabolism in postharvest blueberry fruit. Front. Plant Sci. 2022, 13, 1024909. [Google Scholar] [CrossRef]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine responses to heat stress and global warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, S.; Dinis, L.T.; Machado, N.; Moutinho-Pereira, J. Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron. Sustain. 2018, 38, 66. [Google Scholar] [CrossRef]
- Campa, A.; Ferreira, J.J. Genetic diversity assessed by genotyping by sequencing (GBS) and for phenological traits in blueberry cultivars. PLoS ONE 2018, 13, e0206361. [Google Scholar] [CrossRef] [PubMed]
- Tarara, J.M.; Lee, J.; Spayd, S.E.; Scagel, C.F. Berry temperature and solar radiation alter acylation, proportion, and concentration of anthocyanin in Merlot grapes. Am. J. Enol. Viticult. 2008, 59, 235–247. [Google Scholar] [CrossRef]
- Timmers, M.A.; Grace, M.H.; Yousef, G.G.; Lila, M.A. Inter-and intra-seasonal changes in anthocyanin accumulation and global metabolite profiling of six blueberry genotypes. J. Food Compos. Anal. 2017, 59, 105–110. [Google Scholar] [CrossRef]
- Zhu, L.; Huang, Y.; Zhang, Y.; Xu, C.; Lu, J.; Wang, Y. The growing season impacts the accumulation and composition of flavonoids in grape skins in two-crop-a-year viticulture. J. Food Sci. Technol. 2017, 54, 2861–2870. [Google Scholar] [CrossRef]
- Spinardi, A.; Cola, G.; Gardana, C.S.; Mignani, I. Variation of anthocyanin content and profile throughout fruit development and ripening of highbush blueberry cultivars grown at two different altitudes. Front. Plant Sci. 2019, 10, 1045. [Google Scholar] [CrossRef]
- Cheng, G.; Zhou, S.; Liu, J.; Feng, Q.; Wei, R.; Yu, H.; Wang, B.; Zhang, Y.; Bai, X. Widely targeted metabolomics provides new insights into the flavonoid metabolism in ‘Kyoho’grapes under a two-crop-a-year cultivation system. Sci. Hortic. 2023, 9, 154. [Google Scholar]
- Carrión-Antolí, A.; Martínez-Romero, D.; Guillén, F.; Zapata, P.J.; Serrano, M.; Valero, D. Melatonin pre-harvest treatments leads to maintenance of sweet cherry quality during storage by increasing antioxidant systems. Front. Plant Sci. 2022, 13, 863467. [Google Scholar] [CrossRef]
- Shah, H.M.S.; Singh, Z.; Hasan, M.U.; Afrifa-Yamoah, E.; Woodward, A. Preharvest melatonin application alleviates red drupelet reversion, improves antioxidant potential and maintains postharvest quality of ‘Elvira’blackberry. Postharvest. Biol. Technol. 2023, 203, 112418. [Google Scholar] [CrossRef]
- He, Y.; Bose, S.K.; Wang, W.; Jia, X.; Lu, H.; Yin, H. Pre-harvest treatment of chitosan oligosaccharides improved strawberry fruit quality. Int. J. Mol. Sci. 2018, 19, 2194. [Google Scholar] [CrossRef] [PubMed]
- Lo Piccolo, E.; Quattrocelli, P.; Becagli, M.; Cardelli, R.; El Horri, H.; Guidi, L.; Landi, M.; Pecchia, S. Can Chitosan Applications in Pre-and Post-Harvest Affect the Quality and Antioxidant Contents of Red Raspberries? Sci. Hortic. 2023, 9, 1135. [Google Scholar] [CrossRef]
- Griñán, I.; Morales, D.; Collado-González, J.; Falcón-Rodríguez, A.B.; Torrecillas, A.; Martín-Palomo, M.J.; Galindo, A. Reducing incidence of peel physiopathies and increasing antioxidant activity in pomegranate fruit under different irrigation conditions by preharvest application of chitosan. Sci. Hortic. 2019, 247, 247–253. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- ARPA Lombardia. 2023. Available online: https://www.arpalombardia.it/temi-ambientali/meteo-e-clima/form-richiesta-dati/ (accessed on 1 September 2023).
- Nesmith, D.S. Fruit Development Period of Several Southern Highbush Blueberry Cultivars. Int. J. Fruit Sci. 2012, 12, 249–255. [Google Scholar] [CrossRef]
- Cola, G.; Mariani, L.; Maghradze, D.; Failla, O. Changes in thermal resources and limitations for Georgian viticulture. Aust. J. Grape Wine Res. 2020, 26, 29–40. [Google Scholar] [CrossRef]
- Gough, R.E. Climatic Requirements. In The Highbush Blueberry and Its Management; CRC Press: New York, NY, USA, 1993; pp. 67–74. [Google Scholar]
- Yang, F.H.; Bryla, D.R.; Strik, B.C. Critical temperatures and heating times for fruit damage in northern highbush blueberry. Hortic. Sci. 2019, 54, 2231–2239. [Google Scholar] [CrossRef]
- Schmidt, B.M.; Erdman, J.W., Jr.; Lila, M.A. Effects of food processing on blueberry antiproliferation and antioxidant activity. J. Food Sci. 2005, 70, s389–s394. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2021 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Month | Minimum Daily Temperature | Maximum Daily Temperature | ||||||||
| Nas | Nam | Norm | Pam | Pas | Nas | Nam | Norm | Pam | Pas | |
| January | 0 (0.7) | 2 (4.0) | 22 (21.7) | 7 (3.9) | 0 (0.7) | 0 (0.2) | 2 (5) | 25 (21.1) | 4 (3.9) | 0 (0.8) |
| February | 0 (1.0) | 2 (3.1) | 8 (19.5) | 13 (4.0) | 5 (0.4) | 0 (0.7) | 1 (3.9) | 22 (19.1) | 2 (3.8) | 3 (0.5) |
| March | 1 (0.6) | 6 (4.3) | 19 (20.9) | 5 (4.9) | 0 (0.3) | 0 (0.9) | 4 (4.1) | 22 (21.1) | 5 (4.5) | 0 (0.4) |
| April | 2 (1.2) | 6 (3.6) | 19 (20.6) | 3 (4.3) | 0 (0.3) | 0 (0.4) | 10 (4.9) | 17 (19.6) | 1 (4.8) | 2 (0.3) |
| May | 0 (0.8) | 6 (4.1) | 25 (21.2) | 0 (4.8) | 0 (0.1) | 1 (0.9) | 3 (4.2) | 27 (21.3) | 0 (4.3) | 0 (0.3) |
| June | 0 (0.7) | 0 (3.8) | 21 (20.9) | 8 (4.2) | 1 (0.4) | 0 (0.9) | 0 (3.8) | 28 (20.4) | 2 (4.7) | 0 (0.2) |
| July | 0 (0.9) | 2 (4.4) | 26 (20.9) | 3 (4.5) | 0 (0.3) | 1 (1.1) | 4 (3.7) | 26 (21.8) | 0 (4.1) | 0 (0.3) |
| August | 0 (0.8) | 6 (4.3) | 22 (20.7) | 1 (5.0) | 2 (0.2) | 2 (0.9) | 4 (3.6) | 21 (22.3) | 3 (3.7) | 1 (0.5) |
| September | 0 (0.8) | 0 (4.3) | 21 (20.0) | 8 (4.8) | 1 (0.1) | 0 (0.8) | 1 (3.9) | 26 (20.4) | 3 (4.7) | 0 (0.3) |
| October | 0 (0.7) | 10 (4.1) | 16 (21.4) | 3 (4.6) | 2 (0.2) | 0 (0.7) | 3 (4.1) | 27 (21.3) | 1 (4.3) | 0 (0.6) |
| November | 0 (0.4) | 5 (4.8) | 16 (19.4) | 9 (5.0) | 0 (0.4) | 0 (0.7) | 4 (3.8) | 23 (21) | 3 (3.9) | 0 (0.6) |
| December | 0 (0.4) | 3 (4.4) | 23 (21.2) | 4 (4.1) | 1 (0.9) | 0 (0.7) | 7 (4.1) | 23 (21.2) | 1 (4.4) | 0 (0.6) |
| 2022 | ||||||||||
| Month | Minimum daily temperature | Maximum daily temperature | ||||||||
| Nas | Nam | Norm | Pam | Pas | Nas | Nam | Norm | Pam | Pas | |
| January | 0 (0.7) | 4 (4.0) | 24 (21.7) | 1 (3.9) | 2 (0.7) | 0 (0.2) | 4 (5) | 24 (21.1) | 1 (3.9) | 2 (0.8) |
| February | 0 (1.0) | 2 (3.1) | 18 (19.5) | 5 (4.0) | 3 (0.4) | 0 (0.7) | 2 (3.9) | 18 (19.1) | 5 (3.8) | 3 (0.5) |
| March | 2 (0.6) | 9 (4.3) | 16 (20.9) | 2 (4.9) | 2 (0.3) | 2 (0.9) | 9 (4.1) | 16 (21.1) | 2 (4.5) | 2 (0.4) |
| April | 1 (1.2) | 6 (3.6) | 22 (20.6) | 1 (4.3) | 0 (0.3) | 1 (0.4) | 6 (4.9) | 22 (19.6) | 1 (4.8) | 0 (0.3) |
| May | 0 (0.8) | 0 (4.1) | 12 (21.2) | 13 (4.8) | 6 (0.1) | 0 (0.9) | 0 (4.2) | 12 (21.3) | 13 (4.3) | 6 (0.3) |
| June | 0 (0.7) | 1 (3.8) | 11 (20.9) | 17 (4.2) | 1 (0.4) | 0 (0.9) | 1 (3.8) | 11 (20.4) | 17 (4.7) | 1 (0.2) |
| July | 0 (0.9) | 1 (4.4) | 16 (20.9) | 12 (4.5) | 2 (0.3) | 0 (1.1) | 1 (3.7) | 16 (21.8) | 12 (4.1) | 2 (0.3) |
| August | 0 (0.8) | 0 (4.3) | 19 (20.7) | 12 (5.0) | 0 (0.2) | 0 (0.9) | 0 (3.6) | 19 (22.3) | 12 (3.7) | 0 (0.5) |
| September | 1 (0.8) | 5 (4.3) | 13 (20.0) | 8 (4.8) | 3 (0.1) | 1 (0.8) | 5 (3.9) | 13 (20.4) | 8 (4.7) | 3 (0.3) |
| October | 0 (0.7) | 0 (4.1) | 16 (21.4) | 9 (4.6) | 6 (0.2) | 0 (0.7) | 0 (4.1) | 11 (21.3) | 11 (4.3) | 9 (0.6) |
| November | 0 (0.4) | 0 (4.8) | 23 (19.4) | 5 (5.0) | 2 (0.4) | 0 (0.7) | 1 (3.8) | 19 (21) | 9 (3.9) | 1 (0.6) |
| December | 0 (0.4) | 0 (4.4) | 15 (21.2) | 8 (4.1) | 8 (0.9) | 1 (0.7) | 3 (4.1) | 20 (21.2) | 7 (4.4) | 0 (0.6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cola, G.; Cavenago, B.; Gardana, C.S.; Spinardi, A. Effect of Elicitor Treatments on Quality Attributes in Blueberry: Implications of Cultivar and Environmental Conditions. Plants 2024, 13, 1105. https://doi.org/10.3390/plants13081105
Cola G, Cavenago B, Gardana CS, Spinardi A. Effect of Elicitor Treatments on Quality Attributes in Blueberry: Implications of Cultivar and Environmental Conditions. Plants. 2024; 13(8):1105. https://doi.org/10.3390/plants13081105
Chicago/Turabian StyleCola, Gabriele, Beatrice Cavenago, Claudio Sebastiano Gardana, and Anna Spinardi. 2024. "Effect of Elicitor Treatments on Quality Attributes in Blueberry: Implications of Cultivar and Environmental Conditions" Plants 13, no. 8: 1105. https://doi.org/10.3390/plants13081105