
Jurassic Osmundaceous Landscapes in Patagonia: Exploring the Concept of Ecological Stasis in the Deseado Massif, Argentina

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jablonski, D.; Gould, S.J.; Raup, D.M. The Nature of the Fossil Record: A Biological Perspective. In Patterns and Processes in the History of Life; Raup, D.M., Jablonski, D., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; pp. 7–22. ISBN 978-3-642-70833-6. [Google Scholar]
- Greenwood, D.R. The Taphonomy of Plant Macrofossils. In The Processes of Fossilization; Donovan, S.K., Ed.; Belhaven Press: London, UK, 1991; pp. 141–169. [Google Scholar]
- Spicer, R.A. Plant Taphonomic Processes. In Taphonomy; Allison, P.A., Briggs, D.E.G., Eds.; Topics in Geobiology; Springer: Boston, MA, USA, 1991; Volume 9, pp. 71–113. ISBN 978-1-4899-5036-9. [Google Scholar]
- Taylor, T.N.; Krings, M.; Taylor, E.L. Fossil Fungi; Academic Press: London, UK, 2015. [Google Scholar]
- Taylor, T.N.; Taylor, E.L.; Krings, M. Paleobotany: The Biology and Evolution of Fossil Plants, 2nd ed.; Academic Press: New York, NY, USA, 2009. [Google Scholar]
- Bomfleur, B.; McLoughlin, S.; Vajda, V. Fossilized Nuclei and Chromosomes Reveal 180 Million Years of Genomic Stasis in Royal Ferns. Science 2014, 343, 1376–1377. [Google Scholar] [CrossRef] [PubMed]
- Bomfleur, B.; Decombeix, A.L.; Escapa, I.H.; Schwendemann, A.B.; Axsmith, B. Whole-Plant Concept and Environment Reconstruction of a Telemachus Conifer (Voltziales) from the Triassic of Antarctica. Int. J. Plant Sci. 2013, 174, 425–444. [Google Scholar] [CrossRef]
- Miller, C.N. Evolution of the Fern Family Osmundaceae Based on Anatomical Studies. Contrib. Mus. Paleontol. Univ. Mich. 1971, 23, 105–169. [Google Scholar]
- Phipps, C.J.; Taylor, T.N.; Taylor, E.L.; Cúneo, N.R.; Boucher, L.D.; Yao, X.L. Osmunda (Osmundaceae) from the Triassic of Antarctica: An Example of Evolutionary Stasis. Am. J. Bot. 1998, 85, 888. [Google Scholar] [CrossRef] [PubMed]
- Schwendemann, A.B.; Taylor, T.N.; Taylor, E.L.; Krings, M. Organization, Anatomy, and Fungal Endophytes of a Triassic Conifer Embryo. Am. J. Bot. 2010, 97, 1873–1883. [Google Scholar] [CrossRef]
- DiMichele, W.A.; Behrensmeyer, A.K.; Olszewski, T.D.; Labandeira, C.C.; Pandolfi, J.M.; Wing, S.L.; Bobe, R. Long-Term Stasis in Ecological Assemblages: Evidence from the Fossil Record. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 285–322. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Lehnert, M.; Monjau, T.; Rosche, C. Synopsis of Osmunda (Royal Ferns; Osmundaceae): Towards Reconciliation of Genetic and Biogeographic Patterns with Morphologic Variation. Bot. J. Linn. Soc. 2024, 205, 341–364. [Google Scholar] [CrossRef]
- Escapa, I.H.; Cúneo, N.R. Fertile Osmundaceae from the Early Jurassic of Patagonia, Argentina. Int. J. Plant Sci. 2012, 173, 54–66. [Google Scholar] [CrossRef]
- Qu, Y.; McLoughlin, N.; Van Zuilen, M.A.; Whitehouse, M.; Engdahl, A.; Vajda, V. Evidence for Molecular Structural Variations in the Cytoarchitectures of a Jurassic Plant. Geology 2019, 47, 325–329. [Google Scholar] [CrossRef]
- Schneider, H.; Liu, H.; Clark, J.; Hidalgo, O.; Pellicer, J.; Zhang, S.; Kelly, L.J.; Fay, M.F.; Leitch, I.J. Are the Genomes of Royal Ferns Really Frozen in Time? Evidence for Coinciding Genome Stability and Limited Evolvability in the Royal Ferns. New Phytol. 2015, 207, 10–13. [Google Scholar] [CrossRef]
- Tidwell, W.D.; Ash, S.R. A Review of Selected Triassic to Early Cretaceous Ferns. J. Plant Res. 1994, 107, 417–442. [Google Scholar] [CrossRef]
- Sagasti, A.J.; Massini, J.G.; Escapa, I.H.; Guido, D.M.; Channing, A. Millerocaulis zamunerae Sp. Nov. (Osmundaceae) from Jurassic, Geothermally Influenced, Wetland Environments of Patagonia, Argentina. Alcheringa Australas. J. Palaeontol. 2016, 40, 456–474. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, F.; Yang, X.; Sun, T. Two New Species of Mesozoic Tree Ferns (Osmundaceae: Osmundacaulis) in Eurasia as Evidence of Long-Term Geographic Isolation. Geosci. Front. 2020, 11, 1875–1888. [Google Scholar] [CrossRef]
- Bomfleur, B.; Grimm, G.W.; McLoughlin, S. The Fossil Osmundales (Royal Ferns)—A Phylogenetic Network Analysis, Revised Taxonomy, and Evolutionary Classification of Anatomically Preserved Trunks and Rhizomes. PeerJ 2017, 5, e3433. [Google Scholar] [CrossRef] [PubMed]
- Tryon, R.M.; Tryon, A.F. Ferns and Allied Plants: With Special Reference to Tropical America; Springer: New York, NY, USA, 1982; ISBN 978-1-4613-8164-8. [Google Scholar]
- Vera, E. A New Specimen of Millerocaulis (Osmundales: Osmundaceae) from the Cerro Negro Formation (Lower Cretaceous), Antarctica. Rev. Mus. Argent. Cienc. Nat. Nueva Ser. 2010, 12, 41–46. [Google Scholar] [CrossRef]
- Grimm, G.W.; Kapli, P.; Bomfleur, B.; McLoughlin, S.; Renner, S.S. Using More Than the Oldest Fossils: Dating Osmundaceae with Three Bayesian Clock Approaches. Syst. Biol. 2015, 64, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Guido, D.M.; Campbell, K.A. Jurassic Hot Spring Deposits of the Deseado Massif (Patagonia, Argentina): Characteristics and Controls on Regional Distribution. J. Volcanol. Geotherm. Res. 2011, 203, 35–47. [Google Scholar] [CrossRef]
- García Massini, J.; Escapa, I.H.; Guido, D.M.; Channing, A. First Glimpse of the Silicified Hot Spring Biota from a New Jurassic Chert Deposit in the Deseado Massif, Patagonia, Argentina. Ameghiniana 2016, 53, 205–230. [Google Scholar] [CrossRef]
- Pankhurst, R.J.; Riley, T.R.; Fanning, C.M.; Kelley, S.P. Episodic Silicic Volcanism in Patagonia and the Antarctic Peninsula: Chronology of Magmatism Associated with the Break-up of Gondwana. J. Petrol. 2000, 41, 605–625. [Google Scholar] [CrossRef]
- Guido, D.M. Subdivisión litofacial e interpretación del volcanismo jurásico (Grupo Bahía Laura) en el este del Macizo del Deseado, provincia de Santa Cruz. Rev. Asoc. Geológica Argent. 2004, 59, 727–742. [Google Scholar]
- Pankhurst, R.J.; Leat, P.T.; Sruoga, P.; Rapela, C.W.; Márquez, M.; Storey, B.C.; Riley, T.R. The Chon Aike Province of Patagonia and Related Rocks in West Antarctica: A Silicic Large Igneous Province. J. Volcanol. Geotherm. Res. 1998, 81, 113–136. [Google Scholar] [CrossRef]
- Riley, T.R.; Leat, P.T.; Pankhurst, R.J.; Harris, C. Origins of Large Volume Rhyolitic Volcanism in the Antarctic Peninsula and Patagonia by Crustal Melting. J. Petrol. 2001, 42, 1043–1065. [Google Scholar] [CrossRef]
- Richardson, N.J.; Underhill, J.R. Controls on the Structural Architecture and Sedimentary Character of Syn-Rift Sequences, North Falkland Basin, South Atlantic. Mar. Pet. Geol. 2002, 19, 417–443. [Google Scholar] [CrossRef]
- Guido, D.M.; Campbell, K.A. A Large and Complete Jurassic Geothermal Field at Claudia, Deseado Massif, Santa Cruz, Argentina. J. Volcanol. Geotherm. Res. 2014, 275, 61–70. [Google Scholar] [CrossRef]
- Nunes, C.I.; García Massini, J.; Escapa, I.H.; Guido, D.M.; Campbell, K. Conifer Root Nodules Colonized by Arbuscular Mycorrhizal Fungi in Jurassic Geothermal Settings from Patagonia, Argentina. Int. J. Plant Sci. 2020, 181, 196–209. [Google Scholar] [CrossRef]
- Nunes, C.I.; García Massini, J.L.; Escapa, I.H.; Guido, D.M.; Campbell, K.A. Sooty Molds from the Jurassic of Patagonia, Argentina. Am. J. Bot. 2021, 108, 1464–1482. [Google Scholar] [CrossRef] [PubMed]
- Garcia Massini, J.L.; Guido, D.M.; Campbell, K.C.; Sagasti, A.J.; Krings, M. Filamentous Cyanobacteria and Associated Microorganisms, Structurally Preserved in a Late Jurassic Chert from Patagonia, Argentina. J. S. Am. Earth Sci. 2021, 107, 103111. [Google Scholar] [CrossRef]
- Jones, T.P.; Rowe, N.P. Fossil Plants and Spores: Modern Techniques; Geological Society of London: London, UK, 1999; ISBN 978-1-86239-035-5. [Google Scholar]
- Hass, H.; Rowe, N.P. Thin Section and Wafering. In Fossil Plant and Spores: Modern Techniques; Jones, T.P., Rowe, N.P., Eds.; Geological Society of London: London, UK, 1999; pp. 76–81. [Google Scholar]
- Bercovici, A.; Hadley, A.; Villanueva-Amadoz, U. Improving Depth of Field Resolution for Palynological Photomicrography. Palaeontol. Electron. 2009, 12, 12. [Google Scholar]
- Zarlavsky, G.E. Histología Vegetal: Técnicas Simples y Complejas, 1st ed.; Sociedad Argentina de Botánica: Buenos Aires, Argentina, 2014. [Google Scholar]
- Morris, P.J.; Ivany, L.C.; Schopf, K.M.; Brett, C.E. The Challenge of Paleoecological Stasis: Reassessing Sources of Evolutionary Stability. Proc. Natl. Acad. Sci. USA 1995, 92, 11269–11273. [Google Scholar] [CrossRef]
- Rothwell, G.W.; Stockey, R.A. Onoclea Sensibilis in the Paleocene of North America, a Dramatic Example of Structural and Ecological Stasis. Rev. Palaeobot. Palynol. 1991, 70, 113–124. [Google Scholar] [CrossRef]
- Landi, M.; Angiolini, C. Population Structure of Osmunda Regalis in Relation to Environment and Vegetation: An Example in the Mediterranean Area. Folia Geobot. 2011, 46, 49–68. [Google Scholar] [CrossRef]
- Landi, M.; Angiolini, C. Ecological Responses of Osmunda Regalis to Forest Canopy Cover and Grazing. Am. Fern J. 2010, 100, 137–147. [Google Scholar] [CrossRef]
- Klimešová, J.; Ottaviani, G.; Charles-Dominique, T.; Campetella, G.; Canullo, R.; Chelli, S.; Janovský, Z.; Lubbe, F.C.; Martínková, J.; Herben, T. Incorporating Clonality into the Plant Ecology Research Agenda. Trends Plant Sci. 2021, 26, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Faust, A.; Petersen, R.L. Longevity of Interrupted Fern Colonies. Southeast. Nat. 2015, 14, 203–209. [Google Scholar] [CrossRef]
- Tsakalos, J.L.; Ottaviani, G.; Chelli, S.; Rea, A.; Elder, S.; Dobrowolski, M.P.; Mucina, L. Plant Clonality in a Soil-Impoverished Open Ecosystem: Insights from Southwest Australian Shrublands. Ann. Bot. 2022, 130, 981–990. [Google Scholar] [CrossRef]
- Burr, M.D.; Botero, L.M.; Young, M.J.; Inskeep, W.P.; McDermott, T.R. Observations Concerning Nitrogen Cycling in a Yellowstone Thermal Soil Environment. In Geothermal Biology and Geochemistry in Yellowstone National Park: Proceeding of the Thermal Biology Institute Workshop, Yellowstone National Park, WY; Inskeep, W.P., Ed.; Montana State University Publications: Bozeman, MT, USA, 2005; pp. 171–182. [Google Scholar]
- Price, E.A.C.; Marshall, C. Clonal Plants and Environmental Heterogeneity—An Introduction to the Proceedings. Plant Ecol. 1999, 141, 3–7. [Google Scholar] [CrossRef]
- Escapa, I.H.; Elgorriaga, A.; Nunes, C.; Scasso, R.; Cúneo, N.R. Megafloras del Jurásico en la Cuenca de Cañadón Asfalto: Biomas en Transformación. In Geología y Recursos Naturales de la Provincia de Chubut, 21º Congreso Geológico Argentino, Relatorio; Giacosa, R., Ed.; Asociación Geológica Argentina: Puerto Madryn, Argentina, 2022; pp. 878–901. [Google Scholar]
- Iglesias, A.; Artabe, A.E.; Morel, E.M. The Evolution of Patagonian Climate and Vegetation from the Mesozoic to the Present. Biol. J. Linn. Soc. 2011, 103, 409–422. [Google Scholar] [CrossRef]
- Benzaquén, L.; Blanco, D.E.; Bo, R.; Kandus, P.; Lingua, G.; Minotti, P.; Quintana, R. (Eds.) Regiones de Humedales de la Argentina, 1st ed.; Ministerio de Ambiente y Desarrollo Sustentable, Fundación Humedales: Buenos Aires, Argentina, 2017; ISBN 978-987-29811-6-7. [Google Scholar]
- Guido, D.G.; Channing, A.; Campbell, K.A.; Zamuner, A. Jurassic Geothermal Landscapes and Fossil Ecosystems at San Agustín, Patagonia, Argentina. J. Geol. Soc. Lond. 2010, 167, 11–20. [Google Scholar] [CrossRef]
- Guido, D.M.; Campbell, K.A. Diverse Subaerial and Sublacustrine Hot Spring Settings of the Cerro Negro Epithermal System (Jurassic, Deseado Massif), Patagonia, Argentina. J. Volcanol. Geotherm. Res. 2012, 229–230, 1–12. [Google Scholar] [CrossRef]
- Channing, A. A Review of Active Hot-Spring Analogues of Rhynie: Environments, Habitats and Ecosystems. Philos. Trans. R. Soc. B Biol. Sci. 2017, 373, 20160490. [Google Scholar] [CrossRef] [PubMed]
- van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.-A.; Sanders, I.R. Mycorrhizal Ecology and Evolution: The Past, the Present, and the Future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Schüßler, A.; Walker, C. The Glomeromycota: A Species List with New Families and New Genera; Createspace Independent Pub: Gloucester, UK, 2010. [Google Scholar]
- Morton, J.B.; Redecker, D. Two New Families of Glomales, Archaeosporaceae and Paraglomaceae, with Two New Genera Archaeospora and Paraglomus, Based on Concordant Molecular and Morphological Characters. Mycologia 2001, 93, 181–195. [Google Scholar] [CrossRef]
- Brundrett, M.C. Coevolution of Roots and Mycorrhizas of Land Plants. New Phytol. 2002, 154, 275–304. [Google Scholar] [CrossRef]
- Iqbal, S.H.; Yousaf, M.; Younus, M. A Field Survey of Mycorrhizal Associations in Ferns of Pakistan. New Phytol. 1981, 87, 69–79. [Google Scholar] [CrossRef]
- Brundrett, M.C. Mycorrhizal Associations and Other Means of Nutrition of Vascular Plants: Understanding the Global Diversity of Host Plants by Resolving Conflicting Information and Developing Reliable Means of Diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Brundrett, M.C. Diversity and classification of mycorrhizal associations. Biol. Rev. 2004, 79, 473–495. [Google Scholar] [CrossRef]
- Lehnert, M.; Kottke, I.; Setaro, S.; Pazmiño, L.F.; Suárez, J.P.; Kessler, M. Mycorrhizal Associations in Ferns from Southern Ecuador. Am. Fern J. 2009, 99, 292–306. [Google Scholar] [CrossRef]
- Huey, C.J.; Gopinath, S.C.B.; Uda, M.N.A.; Zulhaimi, H.I.; Jaafar, M.N.; Kasim, F.H.; Yaakub, A.R.W. Mycorrhiza: A Natural Resource Assists Plant Growth under Varied Soil Conditions. 3 Biotech 2020, 10, 204. [Google Scholar] [CrossRef]
- Wright, D.P.; Scholes, J.D.; Read, D.J. Effects of VA Mycorrhizal Colonization on Photosynthesis and Biomass Production of Trifolium repens L. Plant Cell Environ. 1998, 21, 209–216. [Google Scholar] [CrossRef]
- Du, J.; Yu, F.-H.; Alpert, P.; Dong, M. Arbuscular Mycorrhizal Fungi Reduce Effects of Physiological Integration in Trifolium repens. Ann. Bot. 2009, 104, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Dominiak, M.; Olejniczak, P.; Lembicz, M. Diversified Impact of Mycorrhizal Inoculation on Mother Plants and Daughter Ramets in the Clonally Spreading Plant Hieracium pilosella L. (Asteraceae). Plant Ecol. 2019, 220, 757–763. [Google Scholar] [CrossRef]
- Bittebiere, A.-K.; Benot, M.-L.; Mony, C. Clonality as a Key but Overlooked Driver of Biotic Interactions in Plants. Perspect. Plant Ecol. Evol. Syst. 2020, 43, 125510. [Google Scholar] [CrossRef]
- Streitwolf-Engel, R.; Van der Heijden, M.G.A.; Wiemken, A.; Sanders, I.R. The Ecological Significance of Arbuscular Mycorrhizal Fungal Effects on Clonal Reproduction in Plants. Ecology 2001, 82, 2846–2859. [Google Scholar] [CrossRef]
- Lehnert, M.; Kessler, M. Review Mycorrhizal Relationships in Lycophytes and Ferns. Fern Gaz. 2016, 20, 101–116. [Google Scholar]
- Redman, R.S.; Litvintseva, A.; Sheehan, K.B.; Henson, J.M.; Rodriguez, R.J. Fungi from Geothermal Soils in Yellowstone National Park | Applied and Environmental Microbiology. Appl. Environ. Microbiol. 1999, 65, 5193–5197. [Google Scholar] [CrossRef]
- Redman, R.S.; Sheehan, K.B.; Stout, R.G.; Rodriguez, R.J.; Henson, J.M. Thermotolerance Generated by Plant/Fungal Symbiosis. Science 2002, 298, 1581. [Google Scholar] [CrossRef] [PubMed]
- Channing, A.; Edwards, D. Yellowstone Hot Spring Environments and the Palaeo-Ecophysiology of Rhynie Chert Plants: Towards a Synthesis. Plant Ecol. Divers. 2009, 2, 111–143. [Google Scholar] [CrossRef]
- Sparrow, F.K. Aquatic Phycomycetes, 2nd ed.; University of Michigan Press: Ann Arbor, MI, USA, 1960. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; APS Press: St. Paul, MN, USA, 1998. [Google Scholar]
- Prescott, G.W. How to Know Freshwater Algae. Pictured Key Nature Series, 3rd ed.; Wm. C. Brown Company Publishers: Dubuque, IN, USA, 1980. [Google Scholar]
- Patterson, R.T.; Baker, T.; Burbridge, S.M. Arcellaceans (Thecamoebians) as Proxies of Arsenic and Mercury Contamination in Northeastern Ontario Lakes. J. Foraminifer. Res. 1996, 26, 172–183. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. A Revised Six-Kingdom System of Life. Biol. Rev. 1998, 73, 203–266. [Google Scholar] [CrossRef]
- Dick, M.W. Straminipilous Fungi: Systematics of the Peronosporomycetes Including Accounts of the Marine Straminipilous Protists, the Plasmodiophorids and Similar Organisms; Kluwer Academic Publishers: London, UK, 2001. [Google Scholar]
- Lynn, D.H.; Small, E.B. Phylum Ciliophora. In The Illustrated Guide to the Protozoa; Lee, J.J., Leedale, G.F., Bradbury, P., Eds.; Society of Protozoologists, Allen Press Inc.: Lawrence, MA, USA, 2000; Volume 2, pp. 371–656. [Google Scholar]
- Mesterfield, R. Order Arcellinida. In The Illustrated Guide to the Protozoa; Lee, J.J., Leedale, G.F., Eds.; Society of Protozoologists, Allen Press Inc.: Lawrence, MA, USA, 2000; Volume 2, pp. 827–859. [Google Scholar]
- Patterson, D.J. Free-Living Freshwater Protozoa. A Color Guide; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Karling, J.S. The Plasmodiophorales, 1st ed.; The Author: New York, NY, USA, 1942; pp. 1–168. [Google Scholar]
- Dick, M.W. Morphology and Taxonomy of the Oomycetes, with Special Reference to Saprolegniaceae, Leptomitaceae and Pythiaceae. New Phytol. 1969, 68, 751–775. [Google Scholar] [CrossRef]
- Huann-Ju, H.; Chang, H.-S. Five Species of Pythium, Two Species of Pythiogeton New for Taiwan and Pythium Afertile. Bot. Bull. Acad. Sin. 1976, 17, 141–150. [Google Scholar]
- Kalgutkar, R.M.; Jansonius, J. Synopsis of Fossil Fungal Spores, Mycelia and Fructifications; Palynologists, Contribution Series; American Association of Stratigraphic: Dallas, TX, USA, 2000. [Google Scholar]
- Karling, J.S. Chytridiomycetarum Iconographia; Lubrecht & Cramer Ltd.: Monticello, NY, USA, 1977. [Google Scholar]
- Longcore, J.E. Chytridiomycete Taxonomy since 1960. Mycotaxon 1996, 60, 149–174. [Google Scholar]
- Longcore, J.E. Chytridiomycota. In Systematics and Evolution of Fungi; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Gleason, F.H.; Scholz, B.; Jephcott, T.G.; van Ogtrop, F.F.; Henderson, L.; Lilje, O.; Kittelmann, S.; Macarthur, D.J. Key ecological roles for zoosporic true fungi in aquatic habitats. Microbiol. Spectrum. 2017, 5, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- DiLeo, K.; Donat, K.; Min-Venditti, A.; Dighton, J. A Correlation between Chytrid Abundance and Ecological Integrity in New Jersey Pine Barrens Waters. Fungal Ecol. 2010, 3, 295–301. [Google Scholar] [CrossRef]
- Lund, J.W.G. The Ecology of the Freshwater Algae Potamogeton in the English Lakes. J. Ecol. 1954, 42, 366–385. [Google Scholar]
- Fisher, P.J.; Petrini, O. Fungal Endophytes in Phragmites Australis. Mycol. Res. 1992, 96, 1–5. [Google Scholar]
- Channing, A.; Wujek, D.E. Preservation of Protists within Decaying Plants from Geothermally Influenced Wetlands of Yellowstone National Park, Wyoming, United States. PALAIOS 2010, 25, 347–355. [Google Scholar] [CrossRef]
- Krings, M.; Harper, C.J.; White, J.F.; Barthel, M.; Heinrichs, J.; Taylor, E.L.; Taylor, T.N. Fungi in a Psaronius Root Mantle from the Rotliegend (Asselian, Lower Permian/Cisuralian) of Thuringia, Germany. Rev. Palaeobot. Palynol. 2017, 239, 14–30. [Google Scholar] [CrossRef]
- Ellis, M.B. British Fungi, Part 2; Jarrold Publishing: Norwich, UK, 1976. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; Kew Botanical Garden: Surrey, UK, 1971. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi, 2nd ed.; IHW-Verlag: Eching, Germany, 2007. [Google Scholar]
- Osono, T. Ecology of Ligninolytic Fungi Associated with Leaf Litter Decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Mishra, R.R. Fungi Associated with the Fronds of Pteris vittata L. (Pteridaceae). Trop. Ecol. 1994, 35, 29–33. [Google Scholar]
- Kumar, D.S.; Hyde, K.D. Endophytic Fungal Assemblages in Fern Species: A Preliminary Study. Mycoscience 2004, 45, 334–338. [Google Scholar]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanism of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol. 1996, 34, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W. Dictionary of the Fungi, 3rd ed.; CABI Europe: Wallingford, UK, 2008. [Google Scholar]
- Chandra, G.; Chater, K. Developmental Biology of Streptomyces from the Perspective of 100 Actinobacterial Genome Sequences. FEMS Microbiol. Rev. 2014, 38, 345–379. [Google Scholar] [CrossRef]
- Giovannetti, M.; Sbrana, C.; Avio, L.; Citernesi, A.S.; Logi, C. Differential Hyphal Morphogenesis in Arbuscular Mycorrhizal Fungi during Pre-Infection Stages. New Phytol. 1993, 125, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Vannier, N.; Bittebiere, A.-K.; Vandenkoornhuyse, P.; Mony, C. AM Fungi Patchiness and the Clonal Growth of Glechoma Hederacea in Heterogeneous Environments. Sci. Rep. 2016, 6, 37852. [Google Scholar] [CrossRef]
- Vannier, N.; Mony, C.; Bittebiere, A.-K.; Michon-Coudouel, S.; Biget, M.; Vandenkoornhuyse, P. A Microorganisms’ Journey between Plant Generations. Microbiome 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, F.W.M.R. Wood Decay under the Microscope. Fungal Biol. Rev. 2007, 21, 133–170. [Google Scholar] [CrossRef]
- Christiansen, E.; Krokene, P.; Berryman, A.A.; Franceschi, V.R.; Krekling, T.; Lieutier, F.; Lönneborg, A.; Solheim, H. Mechanical Injury and Fungal Infection Induce Acquired Resistance in Norway Spruce. Tree Physiol. 1999, 19, 399–403. [Google Scholar] [CrossRef]
- Krokene, P.; Nagy, N.E.; Krekling, T. Traumatic Resin Ducts and Polyphenolic Parenchyma Cells in Conifers. In Induced Plant Resistance to Herbivory; Schaller, A., Ed.; Springer Science: Berlin/Heidelberg, Germany, 2008; pp. 147–169. [Google Scholar]
- Shain, L. Dynamic Responses of Differentiated Sapwood to Injury and Infection. Phytopathology 1979, 69, 1143–1147. [Google Scholar] [CrossRef]
- Singh, A.P.; Nilsson, N.; Daniel, D. Variable Resistance of Pinus Sylvestris Wood Components to Attack by Wood Degrading Bacteria. In Recent Advances in Wood Anatomy; Donaldson, L.A., Singh, A.P., Butterfield, B.G., Whitehouse, L., Eds.; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; pp. 408–416. [Google Scholar]
- Singh, A.P.; Kim, Y.S.; Singh, T. Bacterial Degradation of Wood. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier Inc.: Boston, MA, USA, 2016; pp. 169–190. [Google Scholar]
- Labandeira, C.C.; Lucas, S.G.; Kirkland, J.I.; Estep, J.W. The Role of Insects in Late Jurassic to Middle Cretaceous Ecosystems. In Lower and Middle Cretaceous Terrestrial Ecosystems; New Mexico Museum of Natural History and Science Bulletin: Albuquerque, NM, USA, 1998. [Google Scholar]
- Kellogg, D.W.; Taylor, E.L. Evidence of Oribatid Mite Detritivory in Antarctica during the Late Paleozoic and Mesozoic. J. Paleontol. 2004, 78, 1146–1153. [Google Scholar] [CrossRef]
- Haq, M.A. Potential of Oribatid Mites in Biodegradation and Mineralization for Enhancing Plant Productivity. Acarol. Stud. 2019, 1, 101–122. [Google Scholar]
- Tidwell, W.D.; Clifford, H.T. Three New Species of Millerocaulis (Osmundaceae) from Queensland, Australia. Aust. Syst. Bot. 1995, 8, 667–685. [Google Scholar] [CrossRef]
- D’Rozario, A.; Labandeira, C.; Guo, W.-Y.; Yao, Y.-F.; Li, C.-S. Spatiotemporal Extension of the Euramerican Psaronius Component Community to the Late Permian of Cathaysia: In Situ Coprolites in a P. housuoensis Stem from Yunnan Province, Southwest China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 306, 127–133. [Google Scholar] [CrossRef]
- Solomon, J.D. Frass Characteristics for Identifying Insect Borers (Lepidoptera: Cossidae and Sesiidae; Coleoptera: Cerambycidae) in Living Hardwoods. Can. Entomol. 1977, 109, 295–303. [Google Scholar] [CrossRef]
- Solomon, J.D. Guide to Insect Borers in North American Broadleaf Trees and Shrubs; United States Department of Agriculture Forest Service: New Orleans, LA, USA, 1995. [Google Scholar]
- Greppi, C.D.; García Massini, J.L.; Pujana, R.R. Saproxylic Arthropod Borings in Nothofagoxylon Woods from the Miocene of Patagonia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 571, 110369. [Google Scholar] [CrossRef]
- Havrylenko, D.; Winterhalter, J.J. Insectos del Parque Nacional Nahuel Huapí; Administración General de Parques Nacionales y Turismo: Berkeley, CA, USA, 1949. [Google Scholar]
- Yee, M.; Grove, S.J.; Richardson, A.M.M.; Mohammed, C.L. Brown Rot in Inner Heartwood: Why Large Logs Support Characteristic Saproxylic Beetle Assemblages of Conservation Concern. In Insect Biodiversity and Dead Wood: Proceedings of a Symposium for the 22nd International Congress of Entomology; Grove, G.J., Simon, J., Hanula, J.L., Eds.; Department of Agriculture Forest Service, SouthernResearch Station: Asheville, NC, USA, 2006; pp. 42–53. [Google Scholar]
- Kukor, J.J.; Martin, M.M. Cellulose Digestion inMonochamus Marmorator Kby. (Coleoptera: Cerambycidae): Role of Acquired Fungal Enzymes. J. Chem. Ecol. 1986, 12, 1057–1070. [Google Scholar] [CrossRef]
- Wotton, R.S.; Malmqvist, B.; Muotka, T.; Larsson, K. Fecal Pellets from a Dense Aggregation of Suspension-Feeders in a Stream: An Example of Ecosystem Engineering. Limnol. Oceanogr. 1998, 43, 719–725. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and Chemical Defenses of Conifer Bark against Bark Beetles and Other Pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed]
- Shigo, A.L.; Marx, H.G. Compartimentalization of Decay in Trees. Bull. US Dep. Agric. 1977, 405, 73. [Google Scholar]
- Esteban, L.G.; Guindeo, A.; Peraza, C.; de Palacios, P. La Madera y Su Anatomía; Fundacion Conde del Valle de Salazar, Mundi-prensa y AiTiM: Madrid, Spain, 2003. [Google Scholar]
- Hudgins, J.W.; Christiansen, E.; Franceschi, V.R. Induction of Anatomically Based Defense Responses in Stems of Diverse Conifers by Methyl Jasmonate: A Phylogenetic Perspective. Tree Physiol. 2004, 24, 251–264. [Google Scholar] [CrossRef]
- Rombola, C.F.; Greppi, C.D.; Pujana, R.R.; García Massini, J.L.; Bellosi, E.S.; Marenssi, S.A. Brachyoxylon Fossil Woods with Traumatic Resin Canals from the Upper Cretaceous Cerro Fortaleza Formation, Southern Patagonia (Santa Cruz Province, Argentina). Cretac. Res. 2022, 130, 105065. [Google Scholar] [CrossRef]
- Bonello, P.; Blodgett, J.T. Pinus Nigra–Sphaeropsis Sapinea as a Model Pathosystem to Investigate Local and Systemic Effects of Fungal Infection of Pines. Physiol. Mol. Plant Pathol. 2003, 63, 249–261. [Google Scholar] [CrossRef]
- Shortle, W.C. Mechanisms of Compartmentalization of Decay in Living Trees. Phytopathology 1979, 69, 1147–1151. [Google Scholar] [CrossRef]
- Christiansen, E.; Franceschi, V.R.; Nagy, N.E.; Krekling, T.; Berryman, A.A.; Krokene, P.; Solheim, H. Traumatic Resin Duct Formation in Norway Spruce (Picea abies (L.) Karst.) after Wounding or Infection with a Bark Beetle-Associated Blue-Stain Fungus, Ceratocystis Polonica. In Physiology and Genetics of Tree-Phytophage Interactions; Lieutier, F., Mattson, W.J., Wagner, M.R., Eds.; Les Colloques de I’INRA: Versailles, France, 1999; pp. 79–89. [Google Scholar]
- Dighton, J. Fungi in Ecosystem Processes, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; ISBN 978-1-315-37152-8. [Google Scholar]
- Lehnert, M.; Krug, M.; Kessler, M. A Review of Symbiotic Fungal Endophytes in Lycophytes and Ferns—A Global Phylogenetic and Ecological Perspective. Symbiosis 2017, 71, 77–89. [Google Scholar] [CrossRef]
- Ogura-Tsujita, Y.; Hirayama, Y.; Sakoda, A.; Suzuki, A.; Ebihara, A.; Morita, N.; Imaichi, R. Arbuscular Mycorrhizal Colonization in Field-Collected Terrestrial Cordate Gametophytes of Pre-Polypod Leptosporangiate Ferns (Osmundaceae, Gleicheniaceae, Plagiogyriaceae, Cyatheaceae) | Mycorrhiza. Mycorrhiza 2016, 26, 87–97. [Google Scholar] [CrossRef]
- Ohgushi, T. Herbivore-Induced Indirect Interaction Webs on Terrestrial Plants: The Importance of Non-Trophic, Indirect, and Facilitative Interactions. Entomol. Exp. Appl. 2008, 128, 217–229. [Google Scholar] [CrossRef]
- McLoughlin, S.; Bomfleur, B. Biotic Interactions in an Exceptionally Well Preserved Osmundaceous Fern Rhizome from the Early Jurassic of Sweden. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 464, 86–96. [Google Scholar] [CrossRef]
- Bippus, A.C.; Escapa, I.H.; Wilf, P.; Tomescu, A.M.F. Fossil Fern Rhizomes as a Model System for Biotic Interactions Across Geologic Time: Evidence from Patagonia. PeerJ Prepr. 2017, 5, e3422v1. [Google Scholar] [CrossRef]
- Lara-Pérez, L.A.; Zulueta-Rodríguez, R.; Andrade-Torres, A. Micorriza arbuscular, Mucoromycotina y hongos septados oscuros en helechos y licófitas con distribución en México: Una revisión global. Rev. Biol. Trop. 2017, 65, 1062–1081. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, L.-D.; Liu, R.-J. Arbuscular Mycorrhizal Fungi Associated with Common Pteridophytes in Dujiangyan, Southwest China. Mycorrhiza 2004, 14, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Nakashima, C.; Crous, P.W. Cercosporoid Fungi (Mycosphaerellaceae) 1. Species on Other Fungi, Pteridophyta and Gymnospermae. IMA Fungus 2013, 4, 265–345. [Google Scholar] [CrossRef] [PubMed]
- Brunner, F.S.; Deere, J.A.; Egas, M.; Eizaguirre, C.; Raeymaekers, J.A.M. The Diversity of Eco-Evolutionary Dynamics: Comparing the Feedbacks between Ecology and Evolution across Scales. Funct. Ecol. 2019, 33, 7–12. [Google Scholar] [CrossRef]
- Vislobokova, I.A. The Concept of Macroevolution in View of Modern Data. Paleontol. J. 2017, 51, 799–898. [Google Scholar] [CrossRef]







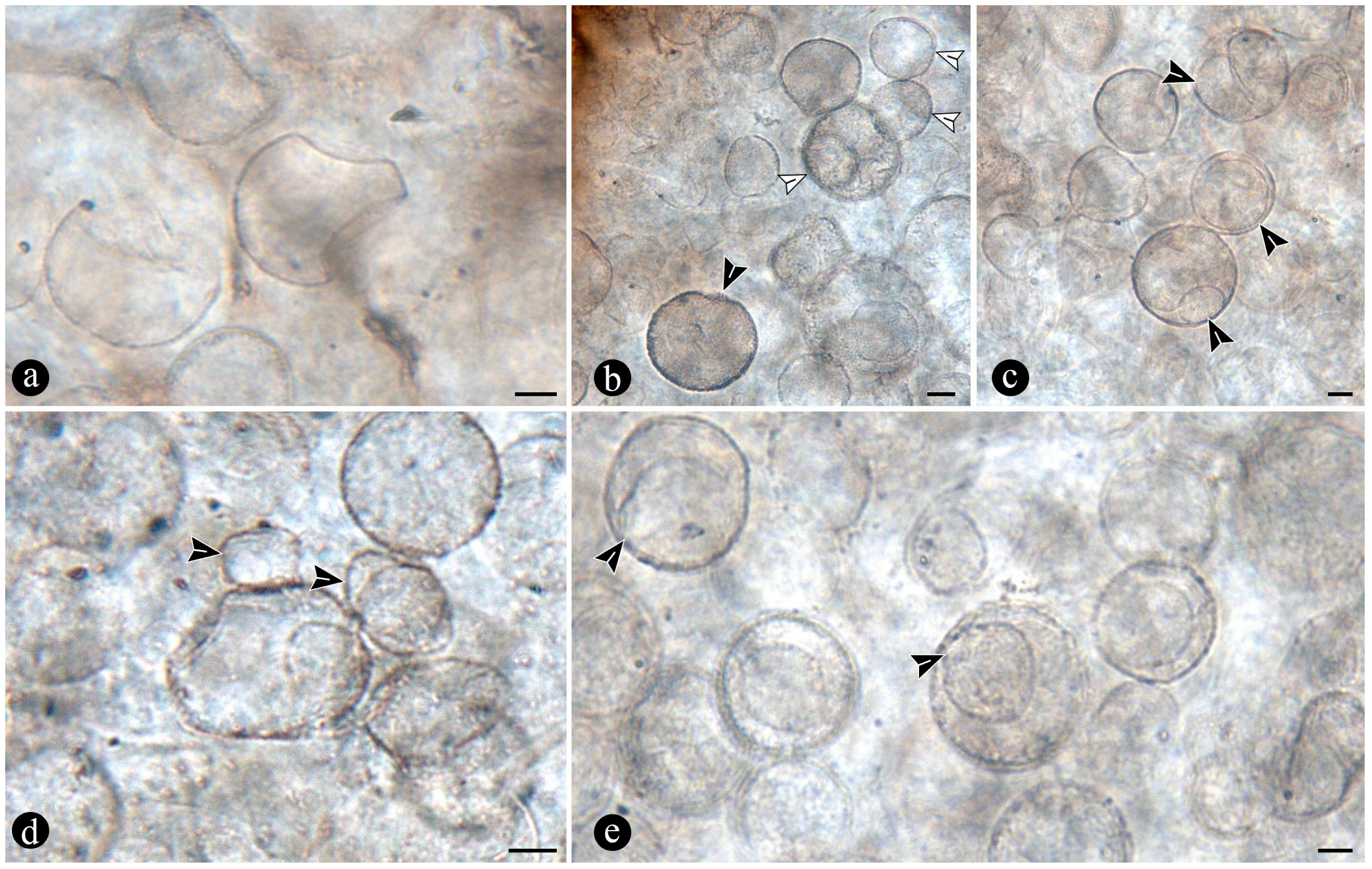







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Distance Between Individuals from Center to Center (cm) | Diameter (cm) | Number of Petiole Cycles | Ramifications Measurements (Smaller Individuals) | Petioles Diameter (cm) | Roots Diameter (cm) | |||
|---|---|---|---|---|---|---|---|---|---|
| Ramification | Diameter | Number of Petiole Cycles | |||||||
| MPM-Pb 16096 | OSLB-08 Big block 1 | 15 | 7 | 7 | - | - | - | - | - |
| 12 | 9 | - | - | - | - | - | |||
| MPM-Pb 16097 | OSLB-08 Big block 2 | 20 | 13 | 15 | 1 | 3 | 5 | - | - |
| - | - | - | 2 | 3 | 5 | - | - | ||
| - | - | - | 3 | 4 | 6 | - | - | ||
| - | 7 | 10 | 1 | - | - | - | - | ||
| - | 6 | 8 | 1 | - | - | - | - | ||
| MPM-Pb 16086 | OSLB-06-2 | - | 5 | 5 | - | - | - | 1.5–10 | 0.6–1.8 |
| MPM-Pb 16087 | OSLB-06-6 | - | 13 | 10 | 1 | - | - | - | - |
| MPM-Pb 16088 | OSLB-06-8 | 10 | 10 | - | - | - | 1.8–73 | - | |
| MPM-Pb 16089 | OSLB-08-6 | - | 2.5 | 5 | - | - | - | - | - |
| - | 1.5 | 3 | - | - | - | - | - | ||
| MPM-Pb 16090 | OSLB-07-2 | - | 8 | 9 | - | - | - | - | - |
| MPM-Pb 16091 | OSLB-10-4 | - | 14 | 13 | - | - | - | - | - |
| MPM-Pb-16084-85 | OSLB-01 Big Block | 14 | 10 | ||||||
| MPM-Pb 16092 | OSLB-06-12 | - | - | - | 1 | 1.5 | 3 | - | - |
| MPM-Pb 16093 | OSLB-07-7 | - | 16 | 12 | - | - | - | - | - |
| MPM-Pb 16094 | OSLB-07-6 | - | 11 | 10 | - | - | - | - | - |
| MPM-Pb 16095 | OSLB-06-7 | - | 12 | 9 | - | - | - | - | - |
| Yañez et al. 596 | 6.2 | 4.6–7 | 5–7 | 1 | 1.2 | 2 | 4–10.2 | 0.35–3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García Massini, J.L.; Nunes, G.C.; Yañez, A.; Escapa, I.H.; Guido, D. Jurassic Osmundaceous Landscapes in Patagonia: Exploring the Concept of Ecological Stasis in the Deseado Massif, Argentina. Plants 2025, 14, 165. https://doi.org/10.3390/plants14020165
García Massini JL, Nunes GC, Yañez A, Escapa IH, Guido D. Jurassic Osmundaceous Landscapes in Patagonia: Exploring the Concept of Ecological Stasis in the Deseado Massif, Argentina. Plants. 2025; 14(2):165. https://doi.org/10.3390/plants14020165
Chicago/Turabian StyleGarcía Massini, Juan L., Giovanni C. Nunes, Agustina Yañez, Ignacio H. Escapa, and Diego Guido. 2025. "Jurassic Osmundaceous Landscapes in Patagonia: Exploring the Concept of Ecological Stasis in the Deseado Massif, Argentina" Plants 14, no. 2: 165. https://doi.org/10.3390/plants14020165
APA StyleGarcía Massini, J. L., Nunes, G. C., Yañez, A., Escapa, I. H., & Guido, D. (2025). Jurassic Osmundaceous Landscapes in Patagonia: Exploring the Concept of Ecological Stasis in the Deseado Massif, Argentina. Plants, 14(2), 165. https://doi.org/10.3390/plants14020165