
Climate Change-Induced Decline in Succulent Euphorbia in Namibia’s Arid Regions


Abstract
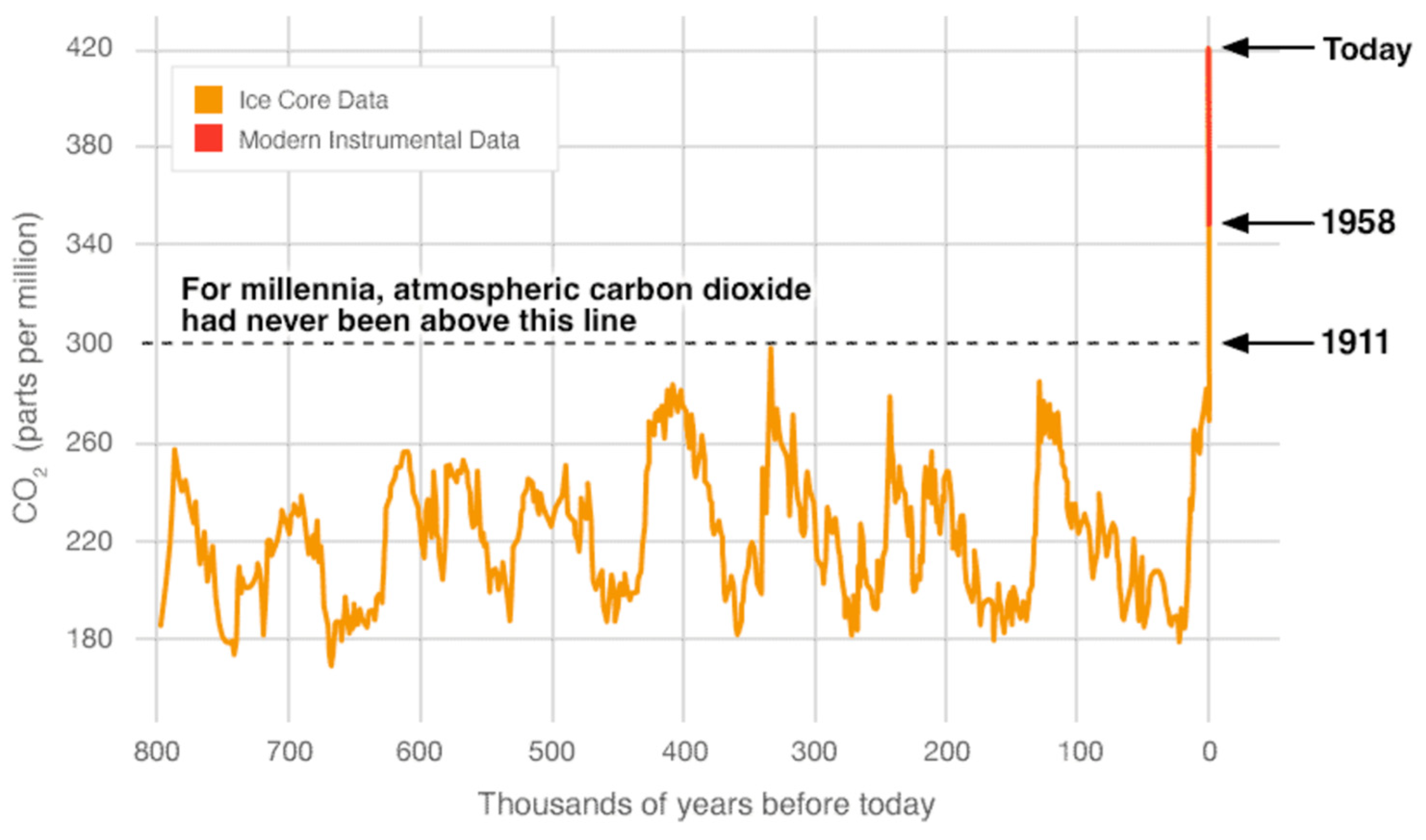
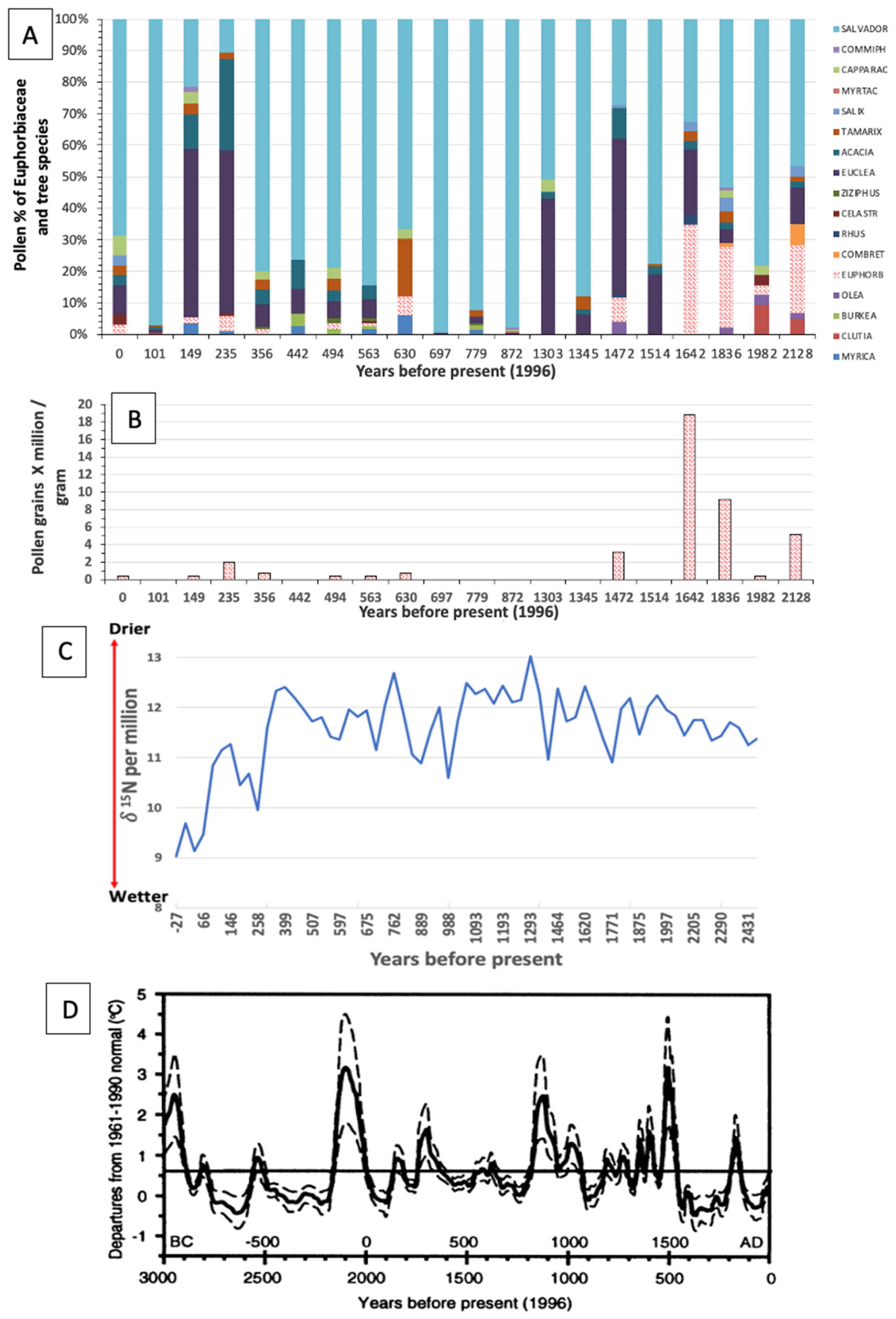
1. Introduction
2. Materials and Methods
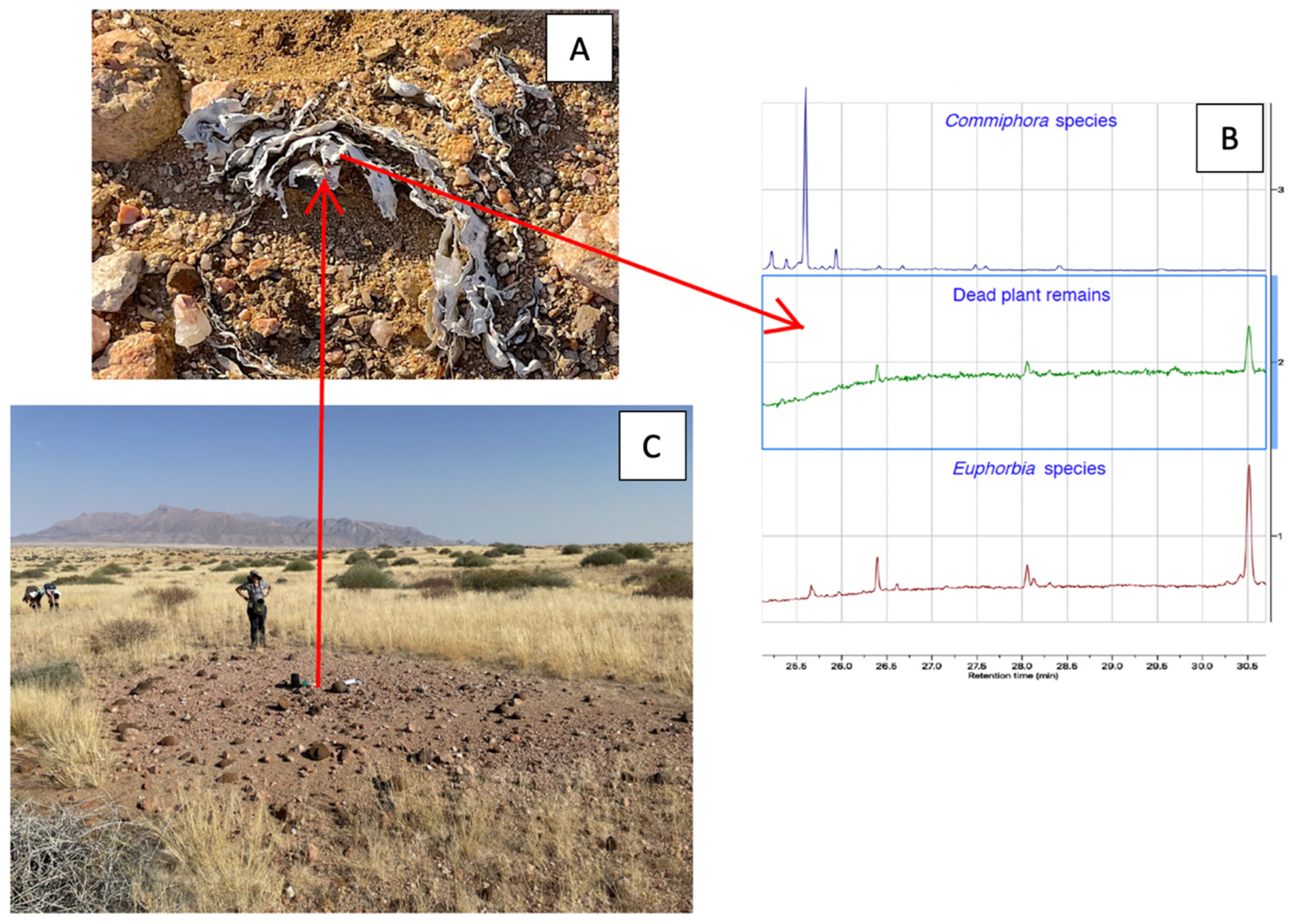
2.1. Plant Collection and Identification
2.2. Euphorbia Population Decline
2.2.1. Historical Photograph Analysis
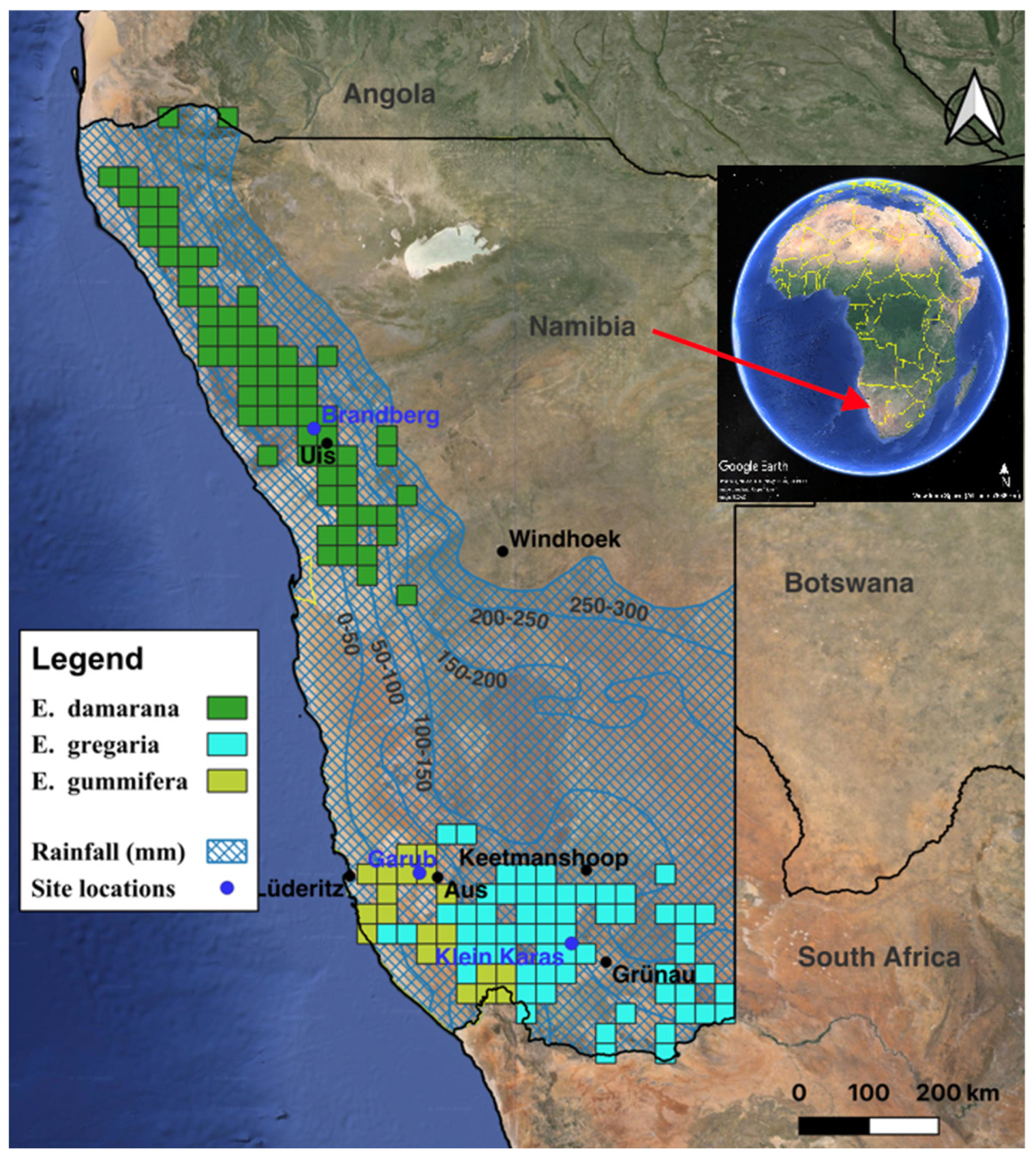
- Brandberg, north-western Namibia, E. damarana, north-west corner of site Brandberg1 (−21.0532306 N, 14.7455993 E) and site Brandberg2 (−21.0530105 N, 14.7535729 E).
- Garub, south-western Namibia, E. gummifera, north-west corner of site Garub1 (−26.628371 N, 16.054868 E) and site Garub2 (−26.6009390 N, 16.0138877 E).
- Klein Karas, south-eastern Namibia, E. gregaria north-west corner of site Klein Karas1 (−27.493063 N, 17.944561 E) and site Klein Karas2 (−27.496656 N, 17.943603 E).
2.2.2. Determination of the Number of Seedlings and Dead Plants Not Detectable Through Remote Sensing
2.3. Normalized Difference Vegetation Index (NDVI) Analysis
2.4. Temperature Data
3. Results
3.1. Historical Photograph Analysis
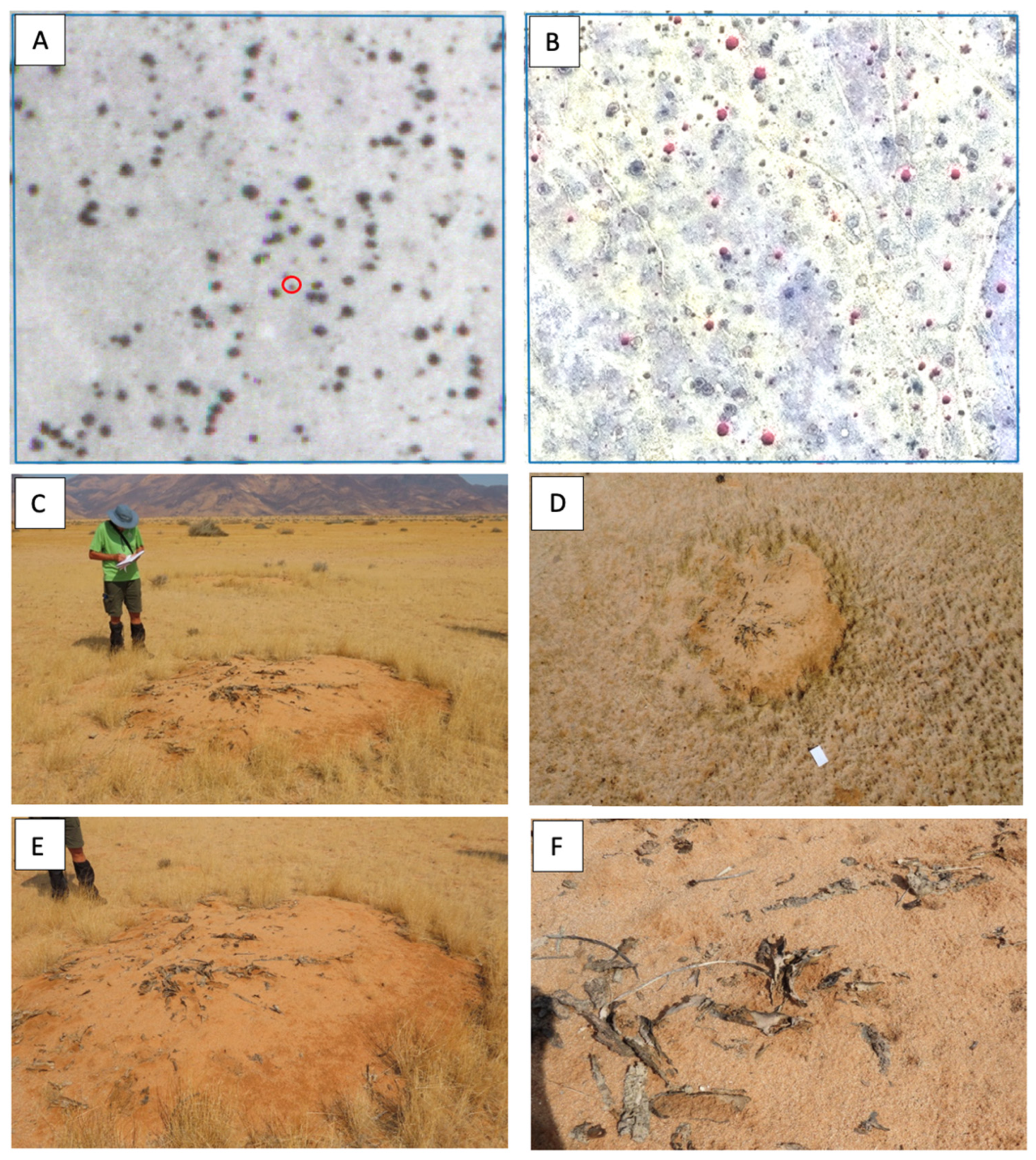
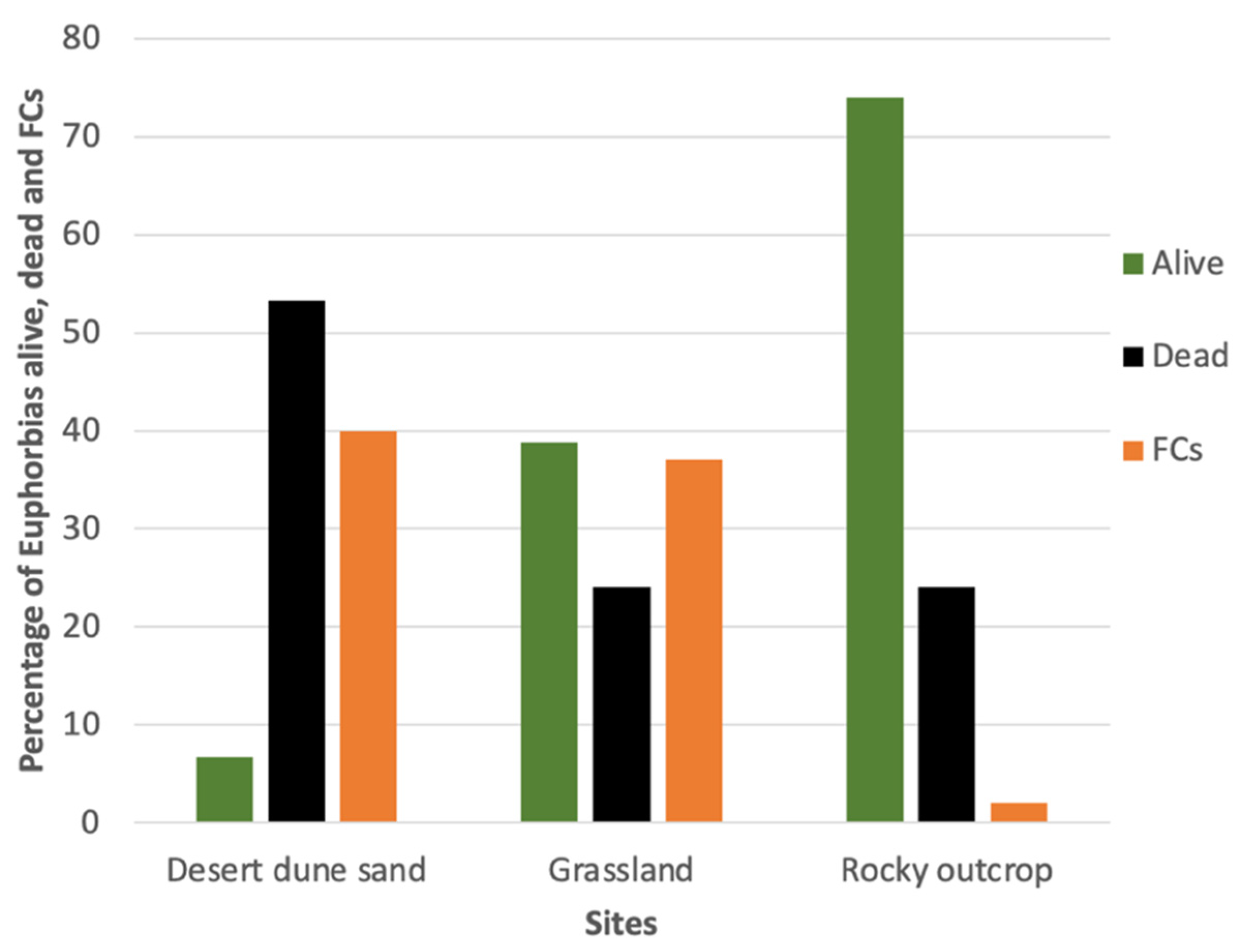
3.2. Presence of Seedlings and the Mortality of Euphorbias Not Detectable Through Remote Sensing
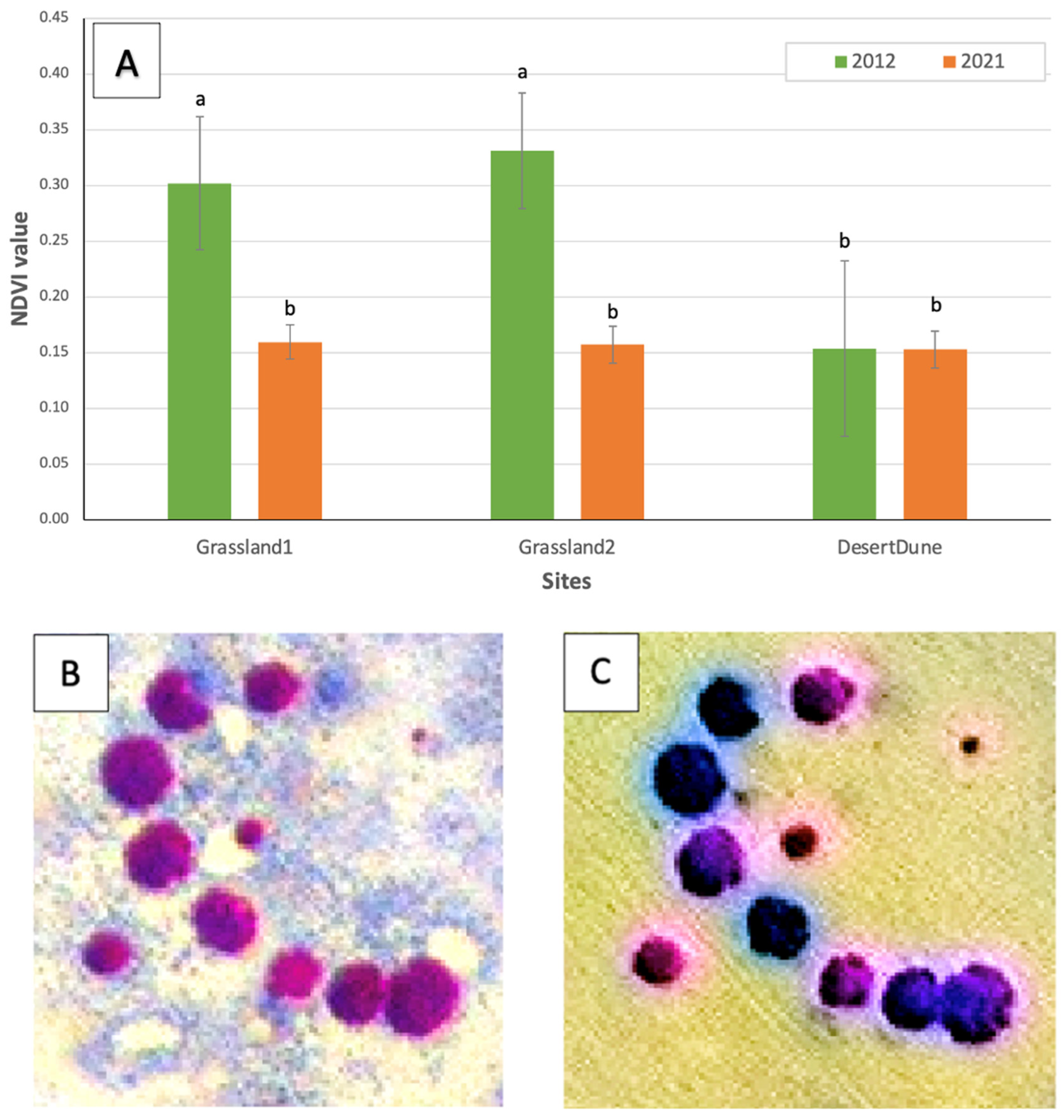
3.3. NDVI Analysis
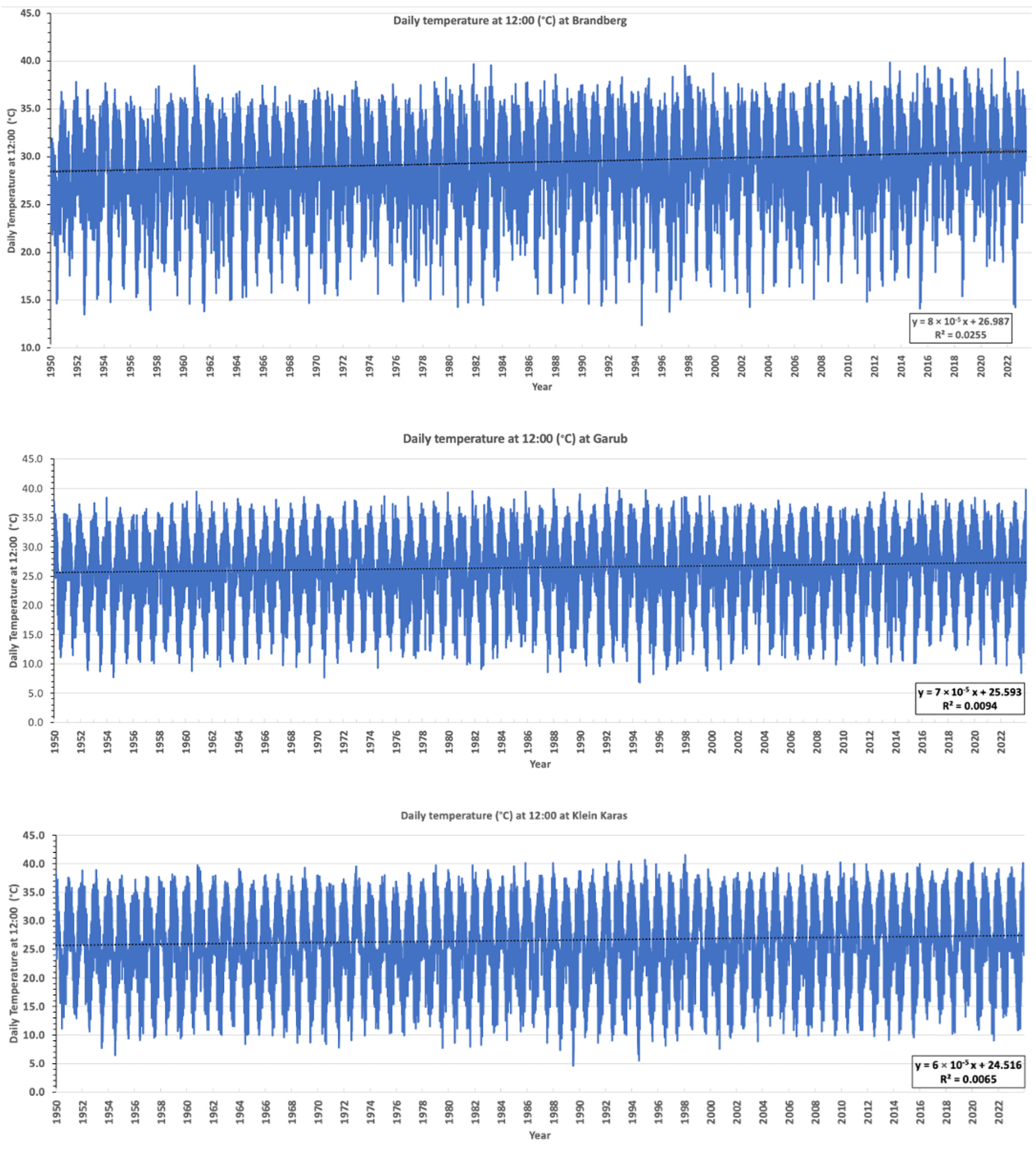
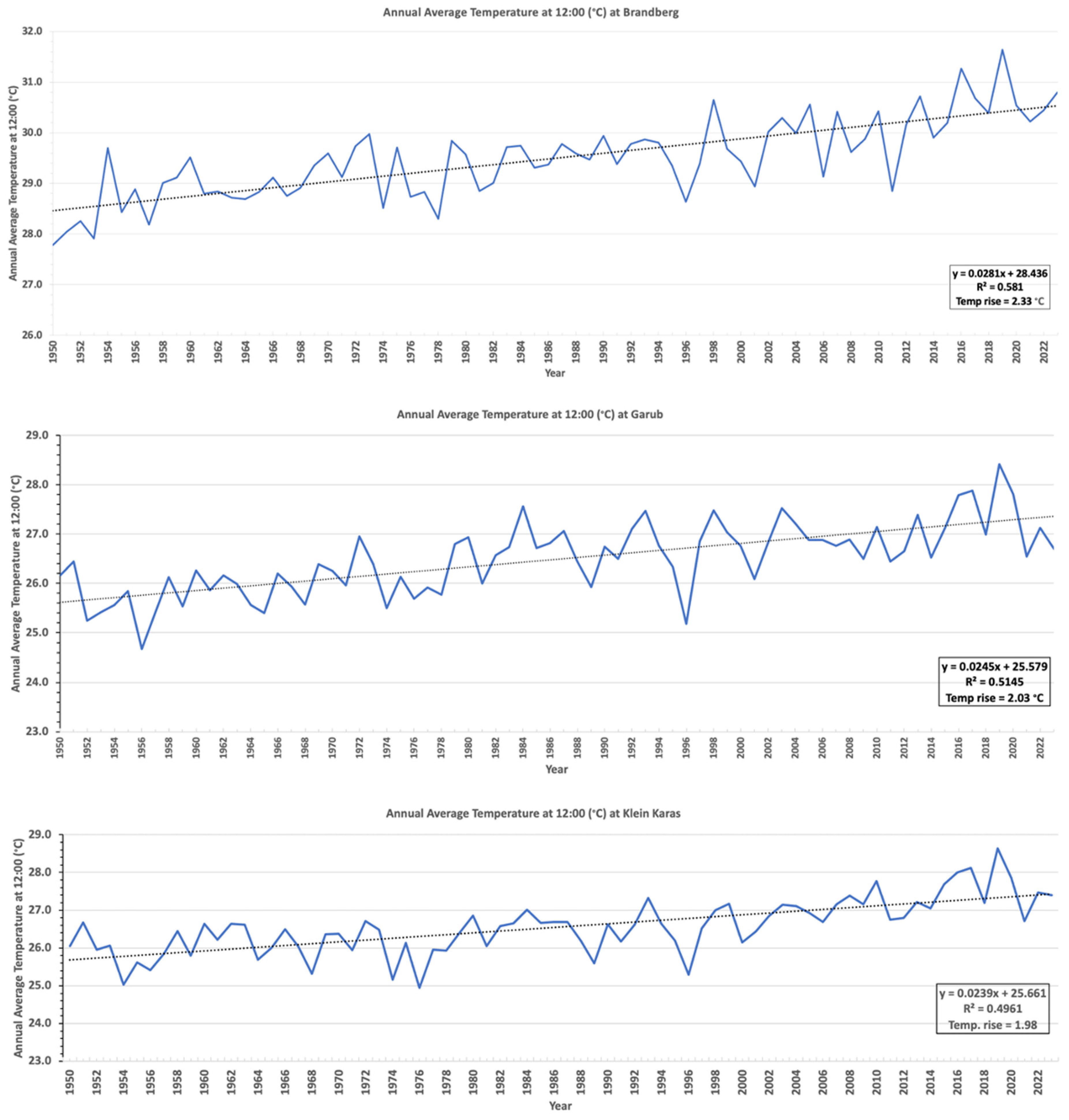
3.4. ERA5 Satellite Temperature Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mapani, B.S.; Shikangalah, R.N.; Mwetulundila, A.L. A review on water security and management under climate change conditions, Windhoek, Namibia. J. Afr. Earth Sci. 2023, 197, e104749. [Google Scholar] [CrossRef]
- Mendelsohn, J.; Jarvis, A.; Roberts, C.; Robertson, T. Atlas of Namibia; Ministry of Environment and Tourism; David Philip Publishers: Cape Town, South Africa, 2002. [Google Scholar]
- NASA Climate Change. 2024. Available online: https://climate.nasa.gov/vital-signs/carbon-dioxide/?intent=121 (accessed on 21 November 2024).
- Bombi, P.; Salvi, D.; Shuuya, T.; Vignoli, L.; Wassenaar, T. Climate change effects on desert ecosystems: A case study on the keystone species of the Namib Desert Welwitschia mirabilis. PLoS ONE 2021, 16, e0259767. [Google Scholar] [CrossRef] [PubMed]
- Foden, W.; Midgley, G.F.; Hughes, G.A.; Bond, W.J.; Thuiller, W.; Hoffman, M.T.; Prince Kaleme, P.; Underhill, L.G.; Rebelo, A.; Lee, H. A changing climate is eroding the geographical range of the Namib Desert tree Aloe through population declines and dispersal lags. Divers. Distrib. 2007, 13, 645–653. [Google Scholar] [CrossRef]
- Meyer, J.J.M.; Schutte, C.E.; Hurter, J.W.; Galt, N.S.; Degashu, P.; Breetzke, G.; Baranenko, D.; Meyer, N.L. The allelopathic, adhesive, hydrophobic and toxic latex of Euphorbia species is the cause of fairy circles investigated at several locations in Namibia. BMC Ecol. 2020, 20, e45. [Google Scholar] [CrossRef] [PubMed]
- Degashu, M.P.; Meyer, J.J.M.; Alberts, P.S.F.; Meyer, N.L.; Blignaut, M.; Makhaba, M.; Hussein, A.A. Isolation and identification of the primary toxin in the smoke of the Namibian milk bush, Euphorbia damarana. SA J. Bot. 2024, 170, 88–96. [Google Scholar] [CrossRef]
- Meyer, J.J.M.; Meyer, A.C.; Meyer, N.L. Sand circles in stony landscapes of Namibia are caused by large Euphorbia shrubs. SA J. Bot. 2020, 130, 25–29. [Google Scholar] [CrossRef]
- Curtis, B.; Mannheimer, C. Tree Atlas of Namibia; National Botanical Research Institute: Windhoek, Namibia, 2005. [Google Scholar]
- Digital Atlas of Namibia. 2023. Available online: https://www.unikoeln.de/sfb389/e/e1/download/atlas_namibia/index_e.htm (accessed on 25 October 2024).
- Hamlyn, G.J.; Robin, A.V. Remote Sensing of Vegetation. Principles, Techniques, and Applications; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Copernicus Climate Change Service. 2024. Available online: https://confluence.ecmwf.int/display/CKB/ERA5%3A+data+documentation (accessed on 20 April 2024).
- Lewandrowski, W.; Stevens, J.C.; Webber, B.L.; Dalziell, E.L.; Trudgen, M.S.; Bateman, A.M.; Todd, E.E. Global change impacts on arid zone ecosystems: Seedling establishment processes are threatened by temperature and water stress. Ecol. Evol. 2021, 11, 8071–8084. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.J.M.; Schutte, C.S.; Galt, N.; Hurter, J.W.; Meyer, N.L. The fairy circles (circular barren patches) of the Namib Desert - What do we know about their cause 50 years after their first description? S. Afr. J. Bot. 2021, 140, 226–239. [Google Scholar] [CrossRef]
- NASA Earth Observatory. 2024. Available online: https://earthobservatory.nasa.gov/world-of-change/global-temperatures (accessed on 25 October 2024).
- NASA GISS. 2024. Available online: https://climate.nasa.gov/vital-signs/global-temperature/?intent=121 (accessed on 22 July 2024).
- US EPA. 2024. Available online: https://www.epa.gov/climate-indicators/climate-change-indicators-us-and-global-temperature (accessed on 25 October 2024).
- Lehmann, D.; Kazgeba, J.; Mfune, E.; Gewers, E.; Cloete, J.; Brain, J.; Voigt, C. Dietary Plasticity of Generalist and Specialist Ungulates in the Namibian Desert: A Stable Isotopes Approach. PLoS ONE 2013, 8, e72190. [Google Scholar] [CrossRef]
- Attwood, K.; Washington, R.; Munday, C. The southern African heat low: Structure, seasonal and diurnal variability, and climatological trends. J. Climatol. 2024, 37, 3037–3053. [Google Scholar] [CrossRef]
- Scott, L. Palynology of hyrax middens: 2000 years of palaeoenvironmental history in Namibia. Quat. Int. 1996, 33, 73–79. [Google Scholar] [CrossRef]
- Chase, B.M.; Boom, A.; Carr, A.S.; Meadows, M.E.; Lim, S. A ca. 39,000-year record of vegetation and climate change from the margin of the Namib Sand Sea. Quat. Res. 2023, 116, 1–11. [Google Scholar] [CrossRef]
- Scott, L.; Bousman, C.B. Palynological analysis of hyrax middens from southern Africa. Paleogeogr. Paleoclimatol. Paleoecol. 1990, 78, 367–379. [Google Scholar] [CrossRef]
- Scott, L.; Cooremans, B. Pollen and recent Procavia (hyrax), Petromus (dassie rat) and bind dung in South Africa. J. Biogeogr. 1992, 19, 205–215. [Google Scholar] [CrossRef]
- Holmgren, K.; Tyson, P.D.; Moberga, A.; Svanered, O. A preliminary 3000-year regional temperature reconstruction for South Africa. S. Afr. J. Sci. 2001, 97, 49–51. [Google Scholar]
- Chase, B.M.; Meadows, M.E.; Scott, L.; Thomas, D.S.G.; Marais, E.; Sealy, J.; Reimer, P.J. A record of rapid Holocene climate change preserved in hyrax middens from southwestern Africa. Geology 2009, 37, 703–706. [Google Scholar] [CrossRef]
- Lim, S.; Chase, B.M.; Chevalier, M.; Reimer, P.J. 50,000 years of vegetation and climate change in the southern Namib Desert, Pella, South Africa. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 451, 197–209. [Google Scholar] [CrossRef]
- Burke, A. Succulent plants on arid inselbergs. Flora 2013, 208, 321–329. [Google Scholar] [CrossRef]
- Hernández-Teixidor, D.; Santos, I.; Suárez, D.; Oromí, P. The importance of threatened host plants for arthropod diversity: The fauna associated with dendroid Euphorbia plants endemic to the Canary and Madeira archipelagos. J. Insect Conserv. 2020, 24, 867–876. [Google Scholar] [CrossRef]
- Taha, A.; Ettaqy, A.; El Mderssa, M.; Belaqziz, M.; Fokar, M.; Boukcim, H.; Zine El Abidine, A.; Abbas, Y. Comprehensive review of morphological adaptations and onservation strategies of cactiform succulents: A case study of Euphorbia species in arid ecosystems. Biosyst. Divers. 2023, 31, 358–367. [Google Scholar] [CrossRef]
- Al-Namazi, A.A.; Al-Khulaidi, A.W.A.; Algarni, S.; Al-Sagheer, N.A. Natural plant species inventory of hotspot areas in Arabian Peninsula: Southwest Al-Baha region, Saudi Arabia. Saudi J. Biol. Sci. 2021, 28, 3309–3324. [Google Scholar] [CrossRef] [PubMed]
- Grace, O.M. Succulent plant diversity as natural capital. Plants People Planet 2019, 1, 336–345. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Historical Photo (Year) | Euphorbias Visible on Historical Photos | Euphorbias Visible on Recent Satellite Images | Euphorbias not Visible (Died) on Satellite Images | Percentage of Dead Euphorbias |
|---|---|---|---|---|---|
| Garub1 | 1967 | 185 | 64 | 121 | 65.4 |
| Garub2 | 1967 | 176 | 92 | 101 | 57.4 |
| Brandberg1 | 1969 | 133 | 17 | 116 | 87.2 |
| Brandberg2 | 1969 | 149 | 7 | 142 | 95.3 |
| Klein Karas1 | 2010 | 255 | 35 | 220 | 86.3 |
| Klein Karas2 | 2010 | 287 | 29 | 258 | 89.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, J.J.M.; Potgieter, M.M.; Meyer, N.L.; Meyer, A.C. Climate Change-Induced Decline in Succulent Euphorbia in Namibia’s Arid Regions. Plants 2025, 14, 190. https://doi.org/10.3390/plants14020190
Meyer JJM, Potgieter MM, Meyer NL, Meyer AC. Climate Change-Induced Decline in Succulent Euphorbia in Namibia’s Arid Regions. Plants. 2025; 14(2):190. https://doi.org/10.3390/plants14020190
Chicago/Turabian StyleMeyer, J. J. Marion, Marie M. Potgieter, Nicole L. Meyer, and Anika C. Meyer. 2025. "Climate Change-Induced Decline in Succulent Euphorbia in Namibia’s Arid Regions" Plants 14, no. 2: 190. https://doi.org/10.3390/plants14020190
APA StyleMeyer, J. J. M., Potgieter, M. M., Meyer, N. L., & Meyer, A. C. (2025). Climate Change-Induced Decline in Succulent Euphorbia in Namibia’s Arid Regions. Plants, 14(2), 190. https://doi.org/10.3390/plants14020190