
Multiprotein Complexes of Plant Glycosyltransferases Involved in Their Function and Trafficking



Abstract
:1. Introduction
2. Protein-Protein Interactions Among GTs Involved in Polysaccharide Synthesis and Protein Glycosylation in Golgi
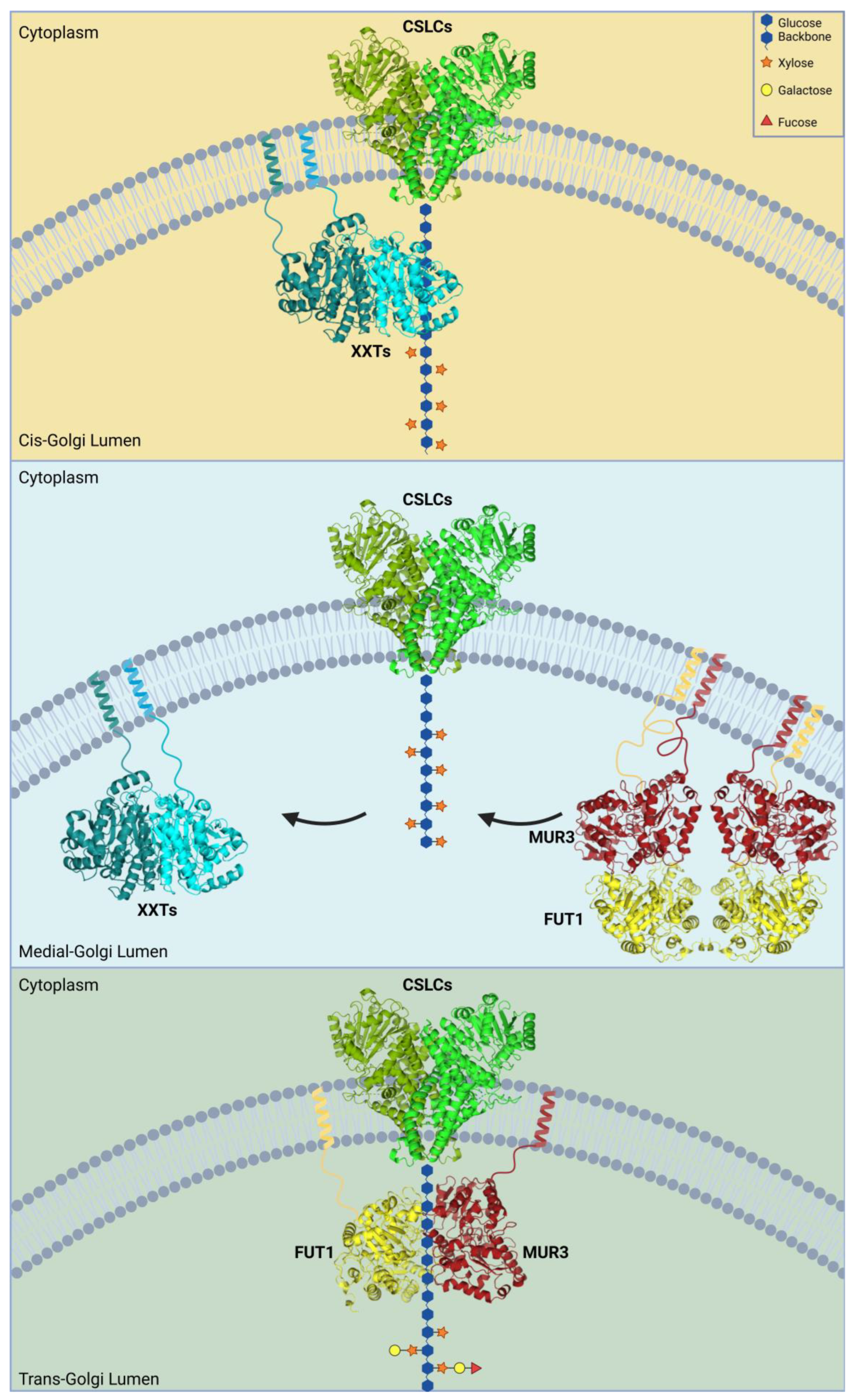
2.1. Xyloglucan Synthase Complex

{kind=link}
{kind=link}
{kind=link}
| Xyloglucan Synthase Complex | |||
| Protein | Heterodimer With | Homodimer | |
| AtCSLC4 | AtXXT2, AtXXT5, AtXLT2, AtMUR3, AtFUT1 |  | [50,59,60] |
| AtXXT1 | AtXXT2, AtXXT5 | | [50,59,60,63] |
| AtXXT2 | AtCSLC4, AtXXT1, AtXXT5, AtXLT2, AtFUT1 | | [50,59,60] |
| AtXXT5 | AtCSLC4, AtXXT1, AtXXT2, AtXLT2, AtFUT1 | [50,59,60] | |
| AtXLT2 | AtCSLC4, AtXXT2, AtXXT5, AtFUT1 | [59,60] | |
| AtMUR3 | AtCSLC4, AtFUT1 | [59,60] | |
| AtFUT1 | AtCSLC4, AtXXT2, AtXXT5, AtXLT2, AtMUR3 | | [59,60,64,65] |
| Xylan Synthase Complex | |||
| Protein | Heterodimer With | Homodimer | |
| AoIRX9 | AoIRX14 | | [66] |
| AoIRX10 | | [66] | |
| AoIRX14 | AoIRX9 | | [66] |
| TaGT43-4 | TaGT47-13, TaGLP, TaVER2, TaGT75-3, TaGT75-4 | | [67] |
| TaGLP | TaGT43-4 | | [67] |
| TaGT75-3 | TaGT43-4 | | [67] |
| TaGT75-4 | TaGT43-4 | | [67] |
| OsGT43B | OsGT47-2 | [68] | |
| OsGT43F | OsGT47-4 | | [68] |
| OsGT43I | OsGT47-1, OsGT47-3 | | [68] |
| OsGT43J | OsGT47-2 | | [68] |
| Pectin Synthase Complex | |||
| Protein | Heterodimer With | Homodimer | |
| AtGAUT1 | AtGAUT5, AtGAUT6, AtGAUT7 | | [69,70,71] |
| AtGAUT10, ATGAUT13, AtGAUT14 | | [70] | |
| Arabinogalactan synthesizing GTs | |||
| Protein | Heterodimer With | Homodimer | |
| AtARAD1 | AtARAD1 | | [72] |
| AtARAD2 | AtARAD2 | | [72] |
| NaARAD1 | AtARAD1 | | [73] |
| AtGALT29A | AtGALT14A, AtGALT31A | | [74] |
| AtGALT31A | | [74] | |
| Proteins involved in N-glycosylation | |||
| Protein | Heterodimer With | Homodimer | |
| GNTI | MNSI | | [75,76] |
| XYLT | GMII | | [75,76] |
| GALTI | GMII | [75] | |
check mark indicates the confirmation of protein homodimer.2.2. Xylan Synthase Complex
2.3. Pectin Synthase Complex
2.4. Arabinogalactan Synthesizing GTs
2.5. Proteins Involved in N-glycosylation
2.6. Structural Analysis of the Homo- and Heterodimerization of GTs
3. The Trafficking of GTs Between ER and Golgi and Their Interaction with Coat Proteins
4. Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cantarel, B.I.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes Database (CAZy): An Expert Resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef] [PubMed]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyl Transferases: Structures, Functions, and Mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Zabotina, O.A.; Zhang, N.; Weerts, R. Polysaccharide Biosynthesis: Glycosyltransferases and Their Complexes. Front. Plant Sci. 2021, 12, 625307. [Google Scholar] [CrossRef]
- Kumar, M.; Turner, S. Plant Cellulose Synthesis: CESA Proteins Crossing Kingdoms. Phytochemistry 2015, 112, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Structure and Growth of Plant Cell Walls. Nat. Rev. Mol. Cell Biol. 2024, 25, 340–358. [Google Scholar] [CrossRef]
- Delmer, D.; Dixon, R.A.; Keegstra, K.; Mohnen, D. The Plant Cell Wall-Dynamic, Strong, and Adaptable-Is a Natural Shapeshifter. Plant Cell 2024, 36, 1257–1311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The Plant Cell Wall: Biosynthesis, Construction, and Functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Cui, D.; Ye, Z.H. Secondary Cell Wall Biosynthesis. New Phytol. 2019, 221, 1703–1723. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.T. We Be Jammin’: An Update on Pectin Biosynthesis, Trafficking and Dynamics. J. Exp. Bot. 2016, 67, 495–502. [Google Scholar] [CrossRef]
- Julian, J.D.; Zabotina, O.A. Xyloglucan Biosynthesis: From Genes to Proteins and Their Functions. Front. Plant Sci. 2022, 13, 920494. [Google Scholar] [CrossRef]
- Mohnen, D. Pectin Structure and Biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Zabotina, O.A. Xyloglucan and Its Biosynthesis. Front. Plant Sci. 2012, 3, 134. [Google Scholar] [CrossRef] [PubMed]
- Polko, J.K.; Kieber, J.J. The Regulation of Cellulose Biosynthesis in Plants. Plant Cell 2019, 31, 282–296. [Google Scholar] [CrossRef]
- van Tol, W.; Wessels, H.; Lefeber, D.J. O-Glycosylation Disorders Pave the Road for Understanding the Complex Human O-Glycosylation Machinery. Curr. Opin. Struct. Biol. 2019, 56, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Van Den Steen, P.; Rudd, P.M.; Dwek, R.A.; Opdenakker, G. Concepts and Principles of O-Linked Glycosylation. Crit. Rev. Biochem. Mol. Biol. 1998, 33, 151–208. [Google Scholar] [CrossRef] [PubMed]
- Joshi, H.J.; Narimatsu, Y.; Schjoldager, K.T.; Tytgat, H.L.P.; Aebi, M.; Clausen, H.; Halim, A. SnapShot: O-Glycosylation Pathways across Kingdoms. Cell 2018, 172, 632–632.e2. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.L.; MacAlister, C.A.; Ulvskov, P. Plant Protein O-Arabinosylation. Front. Plant Sci. 2021, 12, 645219. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.; Seifert, G.; Doblin, M.S.; Johnson, K.L.; Ruprecht, C.; Pfrengle, F.; Bacic, A.; Estevez, J.M. Cracking the “Sugar Code”: A Snapshot of N- and O-Glycosylation Pathways and Functions in Plants Cells. Front. Plant Sci. 2021, 12, 640919. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, D.H.; Yang, B.; D’Souza, A.K.; Shen, D.; Woo, C.M. Truncation of the TPR Domain of OGT Alters Substrate and Glycosite Selection. Anal. Bioanal. Chem. 2021, 413, 7385–7399. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wei, X.; Cong, J.; Zou, J.; Wan, L.; Xu, S. Structural Insights into Mechanism and Specificity of the Plant Protein O-Fucosyltransferase SPINDLY. Nat. Commun. 2022, 13, 7424. [Google Scholar] [CrossRef]
- Aizezi, Y.; Zhao, H.; Zhang, Z.; Bi, Y.; Yang, Q.; Guo, G.; Zhang, H.; Guo, H.; Jiang, K.; Wang, Z.Y. Structure-Based Virtual Screening Identifies Small-Molecule Inhibitors of O-Fucosyltransferase SPINDLY in Arabidopsis. Plant Cell 2024, 36, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Shi, X.; Yang, C.; Zhao, X.; Zhuang, J.; Liu, Y.; Gao, L.; Xia, T. Two UDP-Glycosyltransferases Catalyze the Biosynthesis of Bitter Flavonoid 7-O-Neohesperidoside through Sequential Glycosylation in Tea Plants. J. Agric. Food Chem. 2022, 70, 2354–2365. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.E.; Feng, X.; Ono, N.N.; Holland, D.; Amir, R.; Tian, L. Characterization of a UGT84 Family Glycosyltransferase Provides New Insights into Substrate Binding and Reactivity of Galloylglucose Ester-Forming UGTs. Biochemistry 2017, 56, 6389–6400. [Google Scholar] [CrossRef]
- Yin, Q.; Shen, G.; Chang, Z.; Tang, Y.; Gao, H.; Pang, Y. Involvement of Three Putative Glucosyltransferases from the UGT72 Family in Flavonol Glucoside/Rhamnoside Biosynthesis in Lotus Japonicus Seeds. J. Exp. Bot. 2017, 68, 597–612. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, Y.; von Schaewen, A.; Koiwa, H. Function of N-Glycosylation in Plants. Plant Sci. 2018, 274, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R. Plant Protein Glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef]
- Aebi, M. N-Linked Protein Glycosylation in the ER. Biochim. Biophys. Acta. Mol. Cell Res. 2013, 1833, 2430–2437. [Google Scholar] [CrossRef]
- Liebminger, E.; Hüttner, S.; Vavra, U.; Fischl, R.; Schoberer, J.; Grass, J.; Blaukopf, C.; Seifert, G.J.; Altmann, F.; Mach, L.; et al. Class I α-Mannosidases Are Required for N-Glycan Processing and Root Development in Arabidopsis Thaliana. Plant Cell 2009, 21, 3850–3867. [Google Scholar] [CrossRef] [PubMed]
- Boisson, M.; Gomord, V.; Audran, C.; Berger, N.; Dubreucq, B.; Granier, F.; Lerouge, P.; Faye, L.; Caboche, M.; Lepiniec, L. Arabidopsis Glucosidase I Mutants Reveal a Critical Role of N-Glycan Trimming in Seed Development. EMBO J. 2001, 20, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Liebminger, E.; Christiane, V.; Mach, L.; Strasser, R. Mannose Trimming Reactions in the Early Stages of the N-Glycan Pathway. Plant Signal Behav. 2010, 5, 476–478. [Google Scholar] [CrossRef] [PubMed]
- Von Schaewen, A.; Sturm, A.; O’neill, J.; Chrispeels, M.J. Isolation of a Mutant Arabidopsis Plant that Lacks N-acetyl Glucosaminyl Transferase I and Is Unable to Synthesize Golgi-modified Complex N-linked Glycans. Plant Physiol. 1993, 102, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.; Altmann, F.; Mach, L.; Glössl, J.; Steinkellner, H. Generation of Arabidopsis Thaliana Plants with Complex N-Glycans Lacking Β1,2-Linked Xylose and Core A1,3-Linked Fucose. FEBS Lett. 2004, 561, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Kajiura, H.; Okamoto, T.; Misaki, R.; Matsuura, Y.; Fujiyama, K. Arabidopsis Β1,2-Xylosyltransferase: Substrate Specificity and Participation in the Plant-Specific N-Glycosylation Pathway. J. Biosci. Bioeng. 2012, 113, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Leiter, H.; Mucha, J.; Staudacher, E.; Grimm, R.; Glössl, J.; Altmann, F. Purification, CDNA Cloning, and Expression of GDP-L-Fuc:Asn-Linked GlcNAc A1,3-Fucosyltransferase from Mung Beans. J. Biol. Chem. 1999, 274, 21830–21839. [Google Scholar] [CrossRef]
- Strasser, R.; Mucha, J.; MacH, L.; Altmann, F.; Wilson, I.B.H.; Glössl, J.; Steinkellner, H. Molecular Cloning and Functional Expression of Β1,2-Xylosyltransferase CDNA from Arabidopsis Thaliana. FEBS Lett. 2000, 472, 105–108. [Google Scholar] [CrossRef]
- Wilson, I.B.H.; Zeleny, R.; Kolarich, D.; Staudacher, E.; Stroop, C.J.M.; Kamerling, J.P.; Altmann, F. Analysis of Asn-linked Glycans from Vegetable Foodstuffs: Widespread Occurrence of Lewis A, Core Alpha1,3-linked Fucose and Xylose Substitutions. Glycobiology 2001, 11, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.; Bondili, J.S.; Vavra, U.; Schoberer, J.; Svoboda, B.; Glössl, J.; Léonard, R.; Stadlmann, J.; Altmann, F.; Steinkellner, H.; et al. A Unique Β1,3-Galactosyltransferase Is Indispensable for the Biosynthesis of N-Glycans Containing Lewis a Structures in Arabidopsis Thaliana. Plant Cell 2007, 19, 2278–2292. [Google Scholar] [CrossRef]
- Allen, H.; Wei, D.; Gu, Y.; Li, S. A Historical Perspective on the Regulation of Cellulose Biosynthesis. Carbohydr. Polym. 2021, 252, 117022. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, G.B.; Blaschek, L.; Frandsen, K.E.H.; Noack, L.C.; Persson, S. Cellulose Synthesis in Land Plants. Mol. Plant 2023, 16, 206–231. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Keegstra, K. Biosynthesis of the Plant Cell Wall Matrix Polysaccharide Xyloglucan. Annu. Rev. Plant Biol. 2016, 67, 235–259. [Google Scholar] [CrossRef]
- Hsieh, Y.S.Y.; Harris, P.J. Xyloglucans of Monocotyledons Have Diverse Structures. Mol. Plant 2009, 2, 943–965. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.J.; Darvill, A.G.; Eberhard, S.; York, W.S.; O’Neill, M.A. Moss and Liverwort Xyloglucans Contain Galacturonic Acid and Are Structurally Distinct from the Xyloglucans Synthesized by Hornworts and Vascular Plants. Glycobiology 2008, 18, 891–904. [Google Scholar] [CrossRef]
- Hsieh, Y.S.Y.; Harris, P.J. Structures of Xyloglucans in Primary Cell Walls of Gymnosperms, Monilophytes (Ferns Sensu Lato) and Lycophytes. Phytochemistry 2012, 79, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.; Jia, Z.; Peña, M.J.; Cash, M.; Harper, A.; Blackburn, A.R.; Darvill, A.; York, W.S. Structural Analysis of Xyloglucans in the Primary Cell Walls of Plants in the Subclass Asteridae. Carbohydr. Res. 2005, 340, 1826–1840. [Google Scholar] [CrossRef] [PubMed]
- Schultink, A.; Liu, L.; Zhu, L.; Pauly, M. Structural Diversity and Function of Xyloglucan Sidechain Substituents. Plants 2014, 3, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Fry, S.C.; York, W.S.; Albersheim, P.; Darvill, A.; Hayashi, T.; Joseleau, J.-P.; Kato, Y.; Lorences, E.P.; Maclachlan, G.A.; McNeil, M.; et al. An Unambiguous Nomenclature for Xyloglucan-derived Oligosaccharides. Physiol. Plant 1993, 89, 1–3. [Google Scholar] [CrossRef]
- Tuomivaara, S.T.; Yaoi, K.; O’Neill, M.A.; York, W.S. Generation and Structural Validation of a Library of Diverse Xyloglucan-Derived Oligosaccharides, Including an Update on Xyloglucan Nomenclature. Carbohydr. Res. 2015, 402, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.J.; Kong, Y.; York, W.S.; O’Neill, M.A. A Galacturonic Acid-Containing Xyloglucan Is Involved in Arabidopsis Root Hair Tip Growth. Plant Cell 2012, 24, 4511–4524. [Google Scholar] [CrossRef] [PubMed]
- Cocuron, J.-C.; Lerouxel, O.; Drakakaki, G.; Alonso, A.P.; Liepman, A.H.; Keegstra, K.; Raikhel, N.; Wilkerson, C.G. A Gene from the Cellulose Synthase-like C Family Encodes a Beta-1,4 Glucan Synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 8550–8555. [Google Scholar] [CrossRef]
- Chou, Y.H.; Pogorelko, G.; Zabotina, O.A. Xyloglucan Xylosyltransferases XXT1, XXT2, and XXT5 and the Glucan Synthase CSLC4 Form Golgi-Localized Multiprotein Complexes. Plant Physiol. 2012, 159, 1355–1366. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Chandrasekar, B.; Rea, A.C.; Danhof, L.; Zemelis-Durfee, S.; Thrower, N.; Shepard, Z.S.; Pauly, M.; Brandizzi, F.; Keegstra, K. The Synthesis of Xyloglucan, an Abundant Plant Cell Wall Polysaccharide, Requires CSLC Function. Proc. Natl. Acad. Sci. USA 2020, 117, 20316–20324. [Google Scholar] [CrossRef] [PubMed]
- Faik, A.; Price, N.J.; Raikhel, N.V.; Keegstra, K. An Arabidopsis Gene Encoding An-Xylosyltransferase Involved in Xyloglucan Biosynthesis. Proc. Natl. Acad. Sci. USA 2002, 99, 7797–7802. [Google Scholar] [CrossRef] [PubMed]
- Cavalier, D.M.; Keegstra, K. Two Xyloglucan Xylosyltransferases Catalyze the Addition of Multiple Xylosyl Residues to Cellohexaose. J. Biol. Chem. 2006, 281, 34197–34207. [Google Scholar] [CrossRef] [PubMed]
- Zabotina, O.A.; Avci, U.; Cavalier, D.; Pattathil, S.; Chou, Y.H.; Eberhard, S.; Danhof, L.; Keegstra, K.; Hahn, M.G. Mutations in Multiple XXT Genes of Arabidopsis Reveal the Complexity of Xyloglucan Biosynthesis. Plant Physiol. 2012, 159, 1367–1384. [Google Scholar] [CrossRef]
- Culbertson, A.T.; Chou, Y.H.; Smith, A.L.; Young, Z.T.; Tietze, A.A.; Cottaz, S.; Fauré, R.; Zabotina, O.A. Enzymatic Activity of Xyloglucan Xylosyltransferase 5. Plant Physiol. 2016, 171, 1893–18904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Julian, J.D.; Yap, C.E.; Swaminathan, S.; Zabotina, O.A. The Arabidopsis Xylosyltransferases, XXT3, XXT4, and XXT5 Are Essential to Complete the Fully Xylosylated Glucan Backbone XXXG-type Structure of Xyloglucans. New Phytol. 2023, 238, 1986–1999. [Google Scholar] [CrossRef]
- Madson, M.; Dunand, C.; Li, X.; Verma, R.; Vanzin, G.F.; Caplan, J.; Shoue, D.A.; Carpita, N.C.; Reiter, W.D. The MUR3 Gene of Arabidopsis Encodes a Xyloglucan Galactosyltransferase That Is Evolutionarily Related to Animal Exostosins. Plant Cell 2003, 15, 1662–1670. [Google Scholar] [CrossRef]
- Jensen, J.K.; Schultink, A.; Keegstra, K.; Wilkerson, C.G.; Pauly, M. RNA-Seq Analysis of Developing Nasturtium Seeds (Tropaeolum Majus): Identification and Characterization of an Additional Galactosyltransferase Involved in Xyloglucan Biosynthesis. Mol. Plant 2012, 5, 984–992. [Google Scholar] [CrossRef]
- Chou, Y.H.; Pogorelko, G.; Young, Z.T.; Zabotina, O.A. Protein-Protein Interactions among Xyloglucan-Synthesizing Enzymes and Formation of Golgi-Localized Multiprotein Complexes. Plant Cell Physiol. 2015, 56, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Lund, C.H.; Bromley, J.R.; Stenbæk, A.; Rasmussen, R.E.; Scheller, H.V.; Sakuragi, Y. A Reversible Renilla Luciferase Protein Complementation Assay for Rapid Identification of Protein-Protein Interactions Reveals the Existence of an Interaction Network Involved in Xyloglucan Biosynthesis in the Plant Golgi Apparatus. J. Exp. Bot. 2015, 66, 85–97. [Google Scholar] [CrossRef]
- Chevalier, L.; Bernard, S.; Ramdani, Y.; Lamour, R.; Bardor, M.; Lerouge, P.; Follet-Gueye, M.L.; Driouich, A. Subcompartment Localization of the Side Chain Xyloglucan-Synthesizing Enzymes within Golgi Stacks of Tobacco Suspension-Cultured Cells. Plant J. 2010, 64, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Parsons, H.T.; Stevens, T.J.; McFarlane, H.E.; Vidal-Melgosa, S.; Griss, J.; Lawrence, N.; Butler, R.; Sousa, M.M.L.; Salemi, M.; Willats, W.G.T.; et al. Separating Golgi Proteins from Cis to Trans Reveals Underlying Properties of Cisternal Localization. Plant Cell 2019, 31, 2010–2034. [Google Scholar] [CrossRef] [PubMed]
- Culbertson, A.T.; Ehrlich, J.J.; Choe, J.Y.; Honzatko, R.B.; Zabotina, O.A. Structure of Xyloglucan Xylosyltransferase 1 Reveals Simple Steric Rules That Define Biological Patterns of Xyloglucan Polymers. Proc. Natl. Acad. Sci. USA 2018, 115, 6064–6069. [Google Scholar] [CrossRef] [PubMed]
- Urbanowicz, B.R.; Bharadwaj, V.S.; Alahuhta, M.; Peña, M.J.; Lunin, V.V.; Bomble, Y.J.; Wang, S.; Yang, J.Y.; Tuomivaara, S.T.; Himmel, M.E.; et al. Structural, Mutagenic and in Silico Studies of Xyloglucan Fucosylation in Arabidopsis Thaliana Suggest a Water-Mediated Mechanism. Plant J. 2017, 91, 931–949. [Google Scholar] [CrossRef]
- Rocha, J.; Cicéron, F.; de Sanctis, D.; Lelimousin, M.; Chazalet, V.; Lerouxel, O.; Breton, C. Structure of Arabidopsis Thaliana FUT1 Reveals a Variant of the GT-B Class Fold and Provides Insight into Xyloglucan Fucosylation. Plant Cell 2016, 28, 2352–2364. [Google Scholar] [CrossRef]
- Zeng, W.; Lampugnani, E.R.; Picard, K.L.; Song, L.; Wu, A.M.; Farion, I.M.; Zhao, J.; Ford, K.; Doblin, M.S.; Bacic, A. Asparagus IRX9, IRX10, and IRX14A Are Components of an Active Xylan Backbone Synthase Complex That Forms in the Golgi Apparatus. Plant Physiol. 2016, 171, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Wiemels, R.E.; Soya, A.; Whitley, R.; Held, M.; Faik, A. Composition, Assembly, and Trafficking of a Wheat Xylan Synthase Complex. Plant Physiol. 2016, 170, 1999–2023. [Google Scholar] [CrossRef]
- Javaid, T.; Bhattarai, M.; Venkataraghavan, A.; Held, M.; Faik, A. Specific Protein Interactions between Rice Members of the GT43 and GT47 Families Form Various Central Cores of Putative Xylan Synthase Complexes. Plant J. 2024, 118, 856–878. [Google Scholar] [CrossRef]
- Atmodjo, M.A.; Sakuragi, Y.; Zhu, X.; Burrell, A.J.; Mohanty, S.S.; Atwood, J.A.; Orlando, R.; Scheller, H.V.; Mohnen, D. Galacturonosyltransferase (GAUT)1 and GAUT7 Are the Core of a Plant Cell Wall Pectin Biosynthetic Homogalacturonan:Galacturonosyltransferase Complex. Proc. Natl. Acad. Sci. USA 2011, 108, 20225–20230. [Google Scholar] [CrossRef]
- Engle, K.A.; Amos, R.A.; Yang, J.Y.; Glushka, J.; Atmodjo, M.A.; Tan, L.; Huang, C.; Moremen, K.W.; Mohnen, D. Multiple Arabidopsis Galacturonosyltransferases Synthesize Polymeric Homogalacturonan by Oligosaccharide Acceptor-Dependent or de Novo Synthesis. Plant J. 2022, 109, 1441–1456. [Google Scholar] [CrossRef] [PubMed]
- Lund, C.H.; Stenbæk, A.; Atmodjo, M.A.; Rasmussen, R.E.; Moller, I.E.; Erstad, S.M.; Biswal, A.K.; Mohnen, D.; Mravec, J.; Sakuragi, Y. Pectin Synthesis and Pollen Tube Growth in Arabidopsis Involves Three GAUT1 Golgi-Anchoring Proteins: GAUT5, GAUT6, and GAUT7. Front. Plant. Sci. 2020, 11, 585774. [Google Scholar] [CrossRef] [PubMed]
- Harholt, J.; Jensen, J.K.; Verhertbruggen, Y.; Søgaard, C.; Bernard, S.; Nafisi, M.; Poulsen, C.P.; Geshi, N.; Sakuragi, Y.; Driouich, A.; et al. ARAD Proteins Associated with Pectic Arabinan Biosynthesis Form Complexes When Transiently Overexpressed in Planta. Planta 2012, 236, 115–128. [Google Scholar] [CrossRef]
- Lampugnani, E.R.; Ho, Y.Y.; Moller, I.E.; Koh, P.L.; Golz, J.F.; Bacic, A.; Newbigin, E. A Glycosyltransferase from Nicotiana Alata Pollen Mediates Synthesis of a Linear (1,5)-α-L-Arabinan When Expressed in Arabidopsis. Plant Physiol. 2016, 170, 1962–1974. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Poulsen, P.C.; Vereb, G.; Kaneko, S.; Schulz, A.; Geshi, N. Galactosyltransferases from Arabidopsis Thaliana in the Biosynthesis of Type II Arabinogalactan: Molecular Interaction Enhances Enzyme Activity. BMC Plant Biol. 2014, 14, 90. [Google Scholar] [CrossRef]
- Schoberer, J.; Liebminger, E.; Botchway, S.W.; Strasser, R.; Hawes, C. Time-Resolved Fluorescence Imaging Reveals Differential Interactions of N-Glycan Processing Enzymes across the Golgi Stack in Planta. Plant Physiol. 2013, 161, 1737–1754. [Google Scholar] [CrossRef] [PubMed]
- Schoberer, J.; Izadi, S.; Kierein, C.; Vavra, U.; König-Beihammer, J.; Ruocco, V.; Grünwald-Gruber, C.; Castilho, A.; Strasser, R. The Tobacco GNTI Stem Region Harbors a Strong Motif for Homomeric Protein Complex Formation. Front. Plant Sci. 2023, 14, 1320051. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Jiang, N.; Nadella, R.; Killen, T.L.; Nadella, V.; Faik, A. A Glucurono (Arabino) Xylan Synthase Complex from Wheat Contains Members of the GT43, GT47, and GT75 Families and Functions Cooperatively. Plant Physiol. 2010, 154, 78–97. [Google Scholar] [CrossRef]
- Doong, R.L.; Mohnen, D. Solubilization and Characterization of a Galacturonosyltransferase that Synthesizes the Pectic Polysaccharide Homogalacturonan. Plant J. 1998, 13, 363–374. [Google Scholar] [CrossRef]
- Amos, R.A.; Pattathil, S.; Yang, J.Y.; Atmodjo, M.A.; Urbanowicz, B.R.; Moremen, K.W.; Mohnen, D. A Two-Phase Model for the Non-Processive Biosynthesis of Homogalacturonan Polysaccharides by the GAUT1:GAUT7 Complex. J. Biol. Chem. 2018, 293, 19047–19063. [Google Scholar] [CrossRef] [PubMed]
- Cicéron, F.; Rocha, J.; Kousar, S.; Hansen, S.F.; Chazalet, V.; Gillon, E.; Breton, C.; Lerouxel, O. Expression, Purification and Biochemical Characterization of AtFUT1, a Xyloglucan-Specific Fucosyltransferase from Arabidopsis Thaliana. Biochimie 2016, 128, 183–192. [Google Scholar] [CrossRef]
- Purushotham, P.; Ho, R.; Yu, L.; Fincher, G.B.; Bulone, V.; Zimmer, J. Mechanism of Mixed-Linkage Glucan Biosynthesis by Barley Cellulose Synthase-like CslF6 (1,3;1,4)-β-Glucan Synthase. Sci. Adv. 2022, 8, eadd1596. [Google Scholar] [CrossRef]
- Prabhakar, P.K.; Pereira, J.H.; Taujale, R.; Shao, W.; Bharadwaj, V.S.; Chapla, D.; Yang, J.Y.; Bomble, Y.J.; Moremen, K.W.; Kannan, N.; et al. Structural and Biochemical Insight into a Modular β-1,4-Galactan Synthase in Plants. Nat. Plants 2023, 9, 486–500. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zabotina, O.A. Critical Determinants in ER-Golgi Trafficking of Enzymes Involved in Glycosylation. Plants 2022, 11, 428. [Google Scholar] [CrossRef] [PubMed]
- Schoberer, J.; Vavra, U.; Stadlmann, J.; Hawes, C.; Mach, L.; Steinkellner, H.; Strasser, R. Arginine/Lysine Residues in the Cytoplasmic Tail Promote ER Export of Plant Glycosylation Enzyme. Traffic 2009, 10, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Schoberer, J.; Liebminger, E.; Vavra, U.; Veit, C.; Grünwald-Gruber, C.; Altmann, F.; Botchway, S.W.; Strasser, R. The Golgi Localization of Gnti Requires a Polar Amino Acid Residue within Its Transmembrane Domain. Plant Physiol. 2019, 180, 859–873. [Google Scholar] [CrossRef]
- Tu, L.; Tai, W.C.S.; Chen, L.; Banfield, D.K. Signal-Mediated Dynamic Retention of Glycosyltransferases in the Golgi. Science 2008, 321, 404–407. [Google Scholar] [CrossRef]
- Eckert, E.S.P.; Reckmann, I.; Hellwig, A.; Röhling, S.; El-Battari, A.; Wieland, F.T.; Popoff, V. Golgi Phosphoprotein 3 Triggers Signal-Mediated Incorporation of Glycosyltransferases into Coatomer-Coated (COPI) Vesicles. J. Biol. Chem. 2014, 289, 31319–31329. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, R.; Russo, D.; Kurokawa, K.; Sahu, P.; Lombardi, B.; Supino, D.; Zhukovsky, M.A.; Vocat, A.; Pothukuchi, P.; Kunnathully, V.; et al. Golgi Maturation-dependent Glycoenzyme Recycling Controls Glycosphingolipid Biosynthesis and Cell Growth via GOLPH3. EMBO J. 2021, 40, e107238. [Google Scholar] [CrossRef] [PubMed]
- Welch, L.G.; Peak-Chew, S.Y.; Begum, F.; Stevens, T.J.; Munro, S. Golph3 and Golph3l Are Broad-Spectrum Copi Adaptors for Sorting into Intra-Golgi Transport Vesicles. J. Cell Biol. 2021, 220, e202106115. [Google Scholar] [CrossRef]
- Liu, L.; Doray, B.; Kornfeld, S. Recycling of Golgi Glycosyltransferases Requires Direct Binding to Coatomer. Proc. Natl. Acad. Sci. USA 2018, 115, 8984–8989. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, C.G.; Maccioni, H.J.F. Endoplasmic Reticulum Export of Glycosyltransferases Depends on Interaction of a Cytoplasmic Dibasic Motif with Sar1. Mol. Biol. Cell 2003, 14, 3753–3766. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, J.J.; Weerts, R.M.; Shome, S.; Culbertson, A.T.; Honzatko, R.B.; Jernigan, R.L.; Zabotina, O.A. Xyloglucan Xylosyltransferase 1 Displays Promiscuity Toward Donor Substrates During in Vitro Reactions. Plant Cell Physiol. 2021, 62, 1890–1901. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Phillips, D.R.; Ye, Z.H. A Single Xyloglucan Xylosyltransferase Is Sufficient for Generation of the XXXG Xylosylation Pattern of Xyloglucan. Plant Cell Physiol. 2021, 62, 1589–1602. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Julian, J.D.; Zabotina, O.A. Multiprotein Complexes of Plant Glycosyltransferases Involved in Their Function and Trafficking. Plants 2025, 14, 350. https://doi.org/10.3390/plants14030350
Zhang N, Julian JD, Zabotina OA. Multiprotein Complexes of Plant Glycosyltransferases Involved in Their Function and Trafficking. Plants. 2025; 14(3):350. https://doi.org/10.3390/plants14030350
Chicago/Turabian StyleZhang, Ning, Jordan D. Julian, and Olga A. Zabotina. 2025. "Multiprotein Complexes of Plant Glycosyltransferases Involved in Their Function and Trafficking" Plants 14, no. 3: 350. https://doi.org/10.3390/plants14030350
APA StyleZhang, N., Julian, J. D., & Zabotina, O. A. (2025). Multiprotein Complexes of Plant Glycosyltransferases Involved in Their Function and Trafficking. Plants, 14(3), 350. https://doi.org/10.3390/plants14030350