Differential Mechanisms of Photosynthetic Acclimation to Light and Low Temperature in Arabidopsis and the Extremophile Eutrema salsugineum

Abstract
:1. Introduction
2. Results
2.1. Growth Regimes
2.2. Photosynthetic Pigmentation
2.2.1. Analysis of Chlorophyll and Carotenoid Contents
2.2.2. Effect of Cold Acclimation on Pigmentation across Growth Regimes
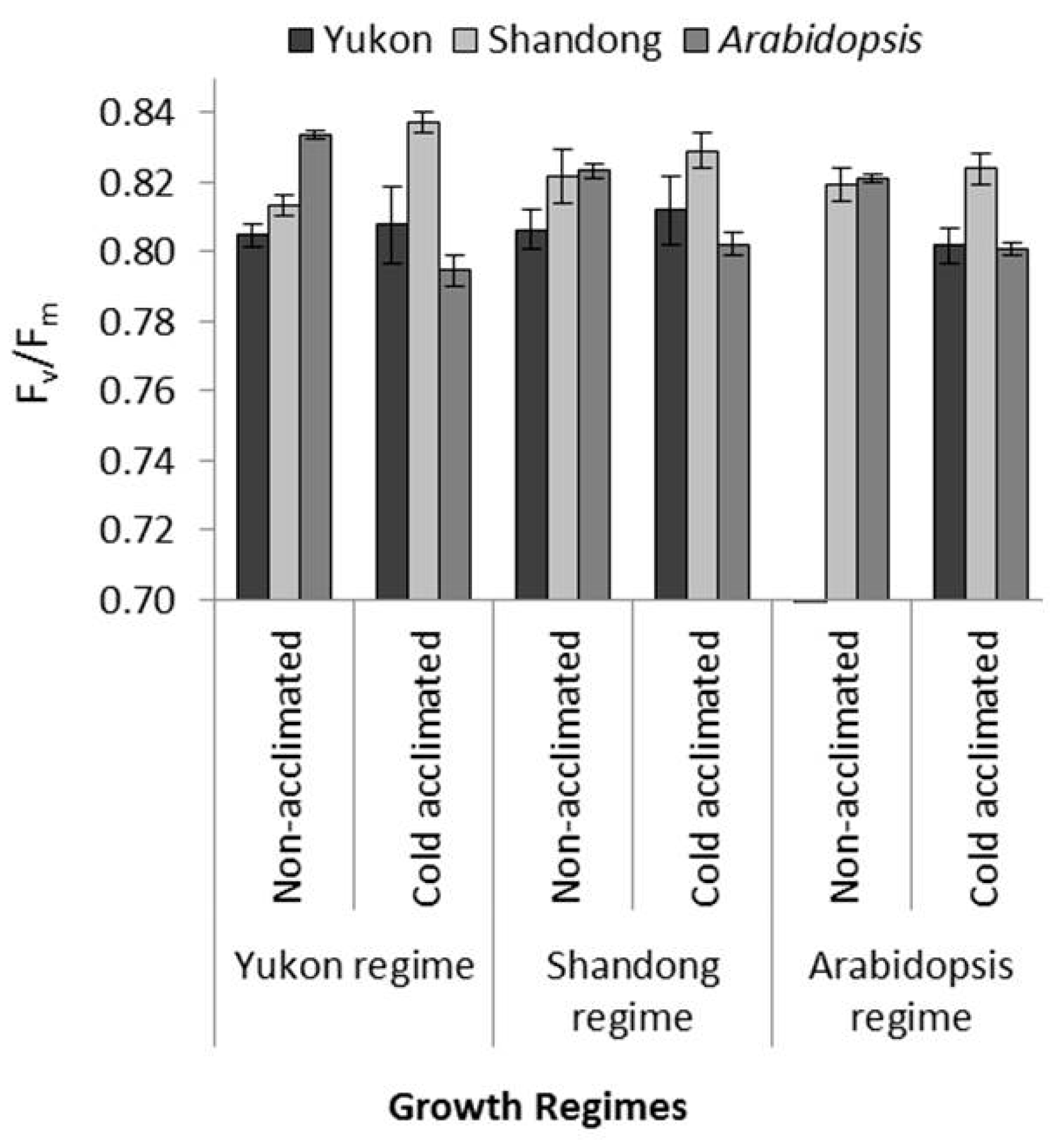
2.3. Comparative PSII Photochemistry
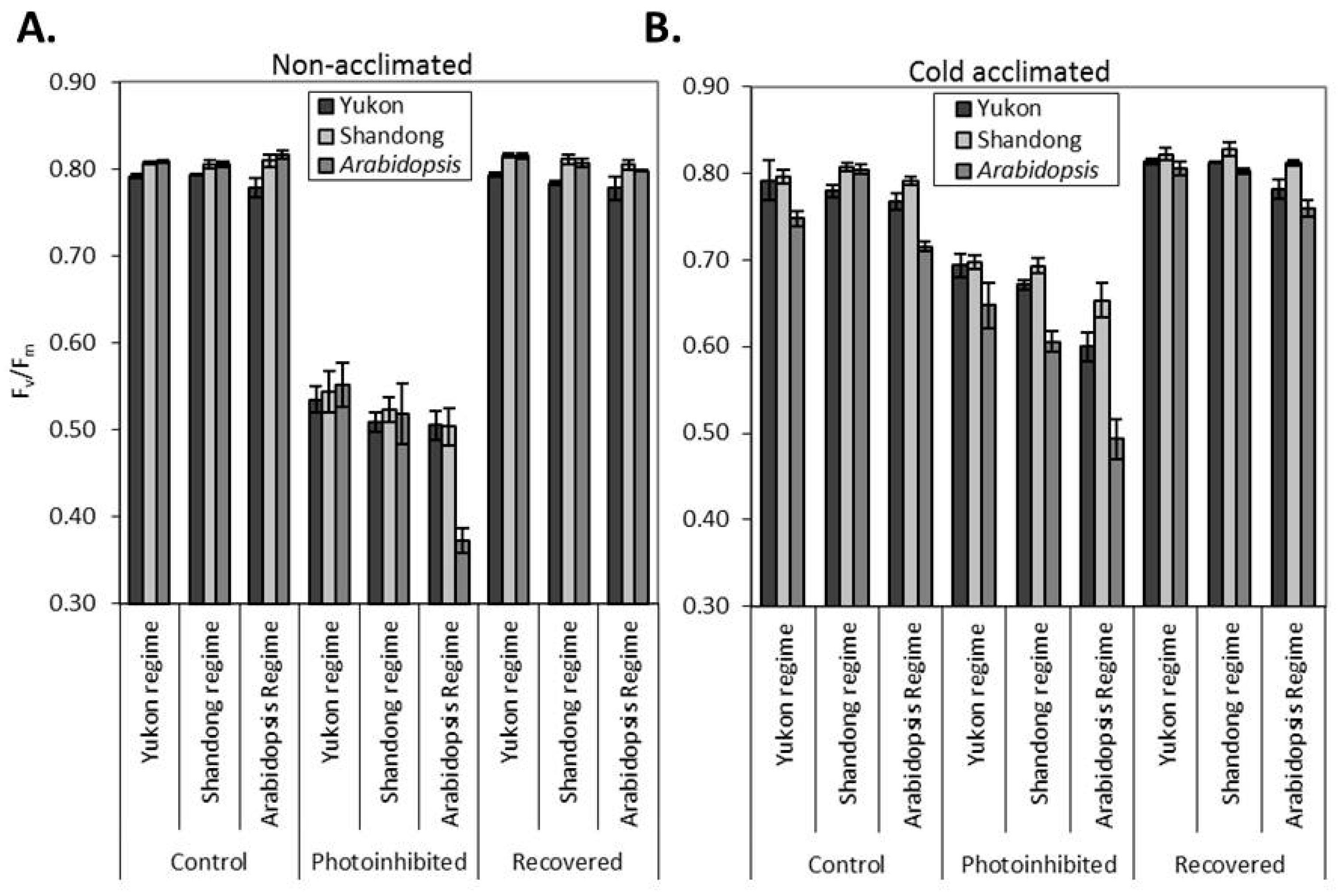
2.4. Photoinhibition and Recovery of PSII
2.5. Chlorophyll Fluorescence Quenching Analyses
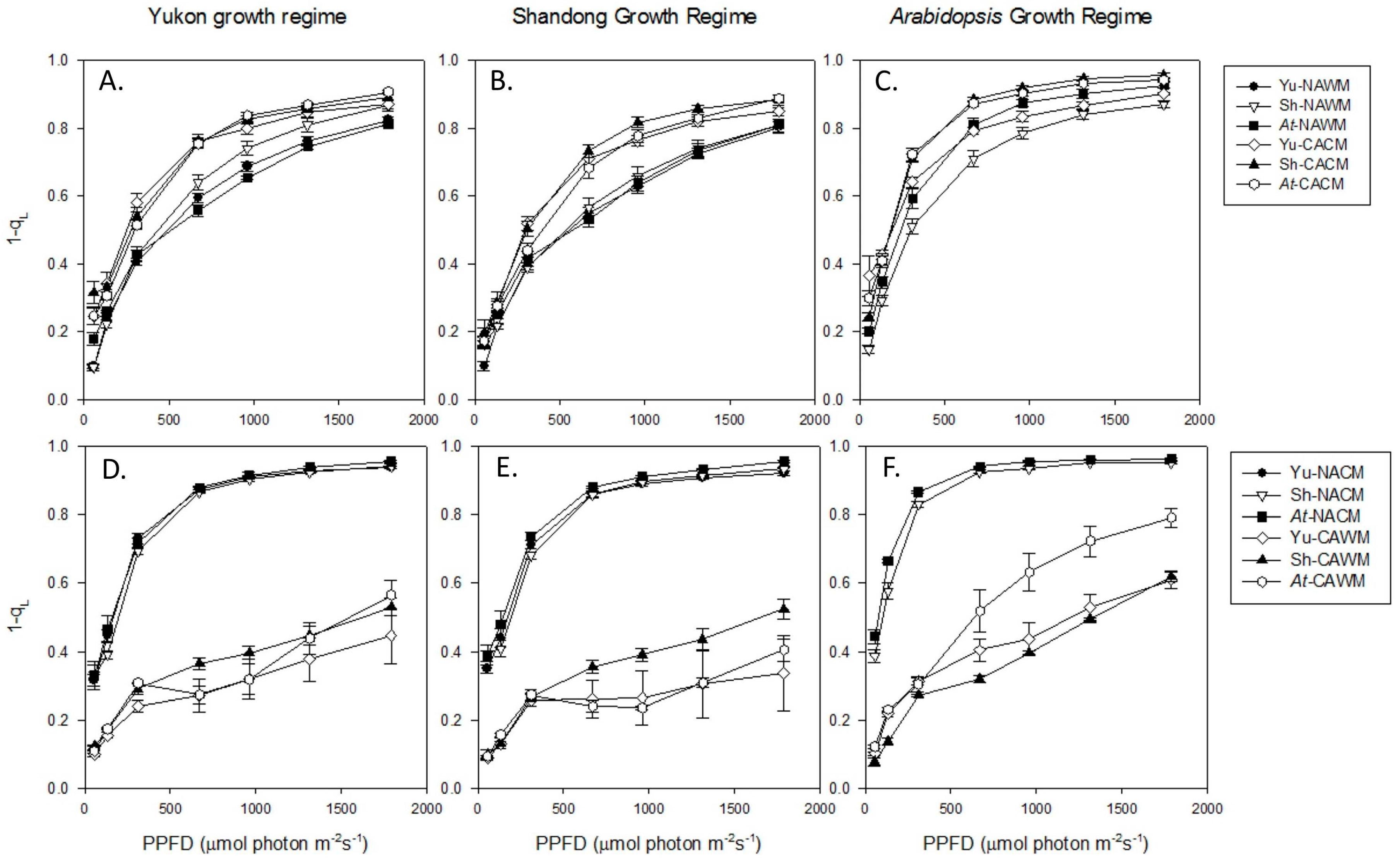
2.5.1. Excitation Pressure (1-qL)
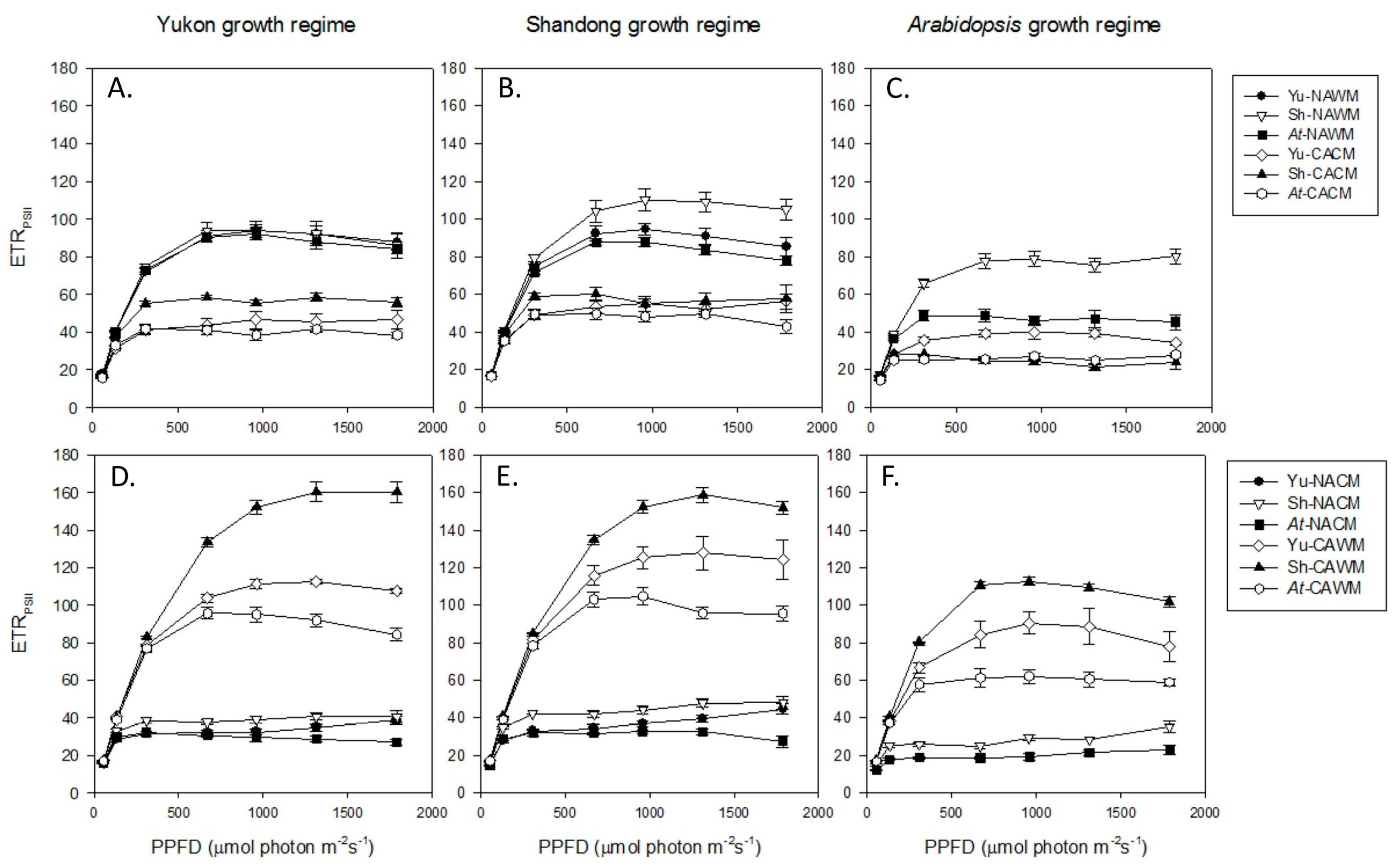
2.5.2. Relative Electron Transport Rate (RETRPSII)
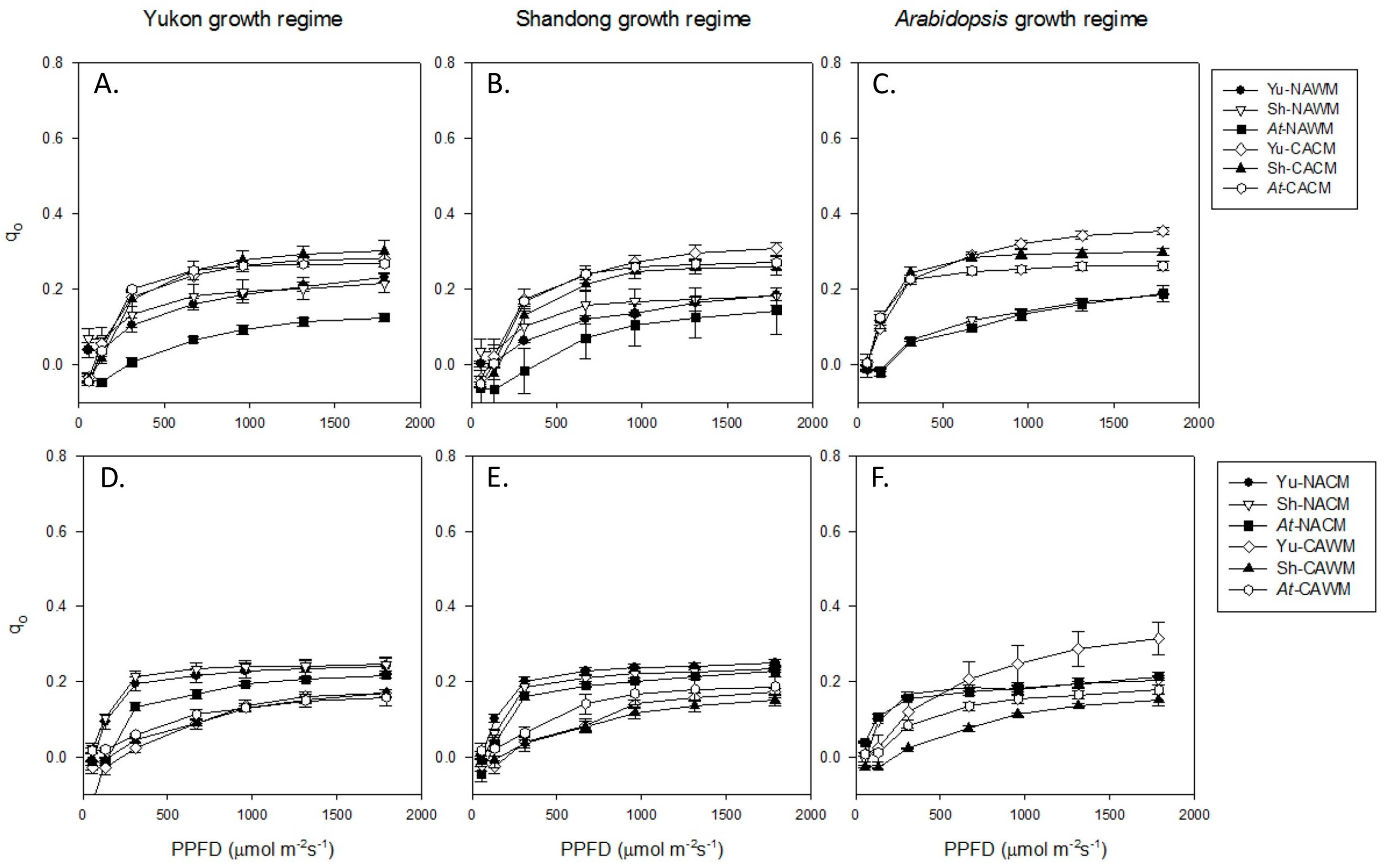
2.5.3. Basal Fluorescence Quenching (qO)
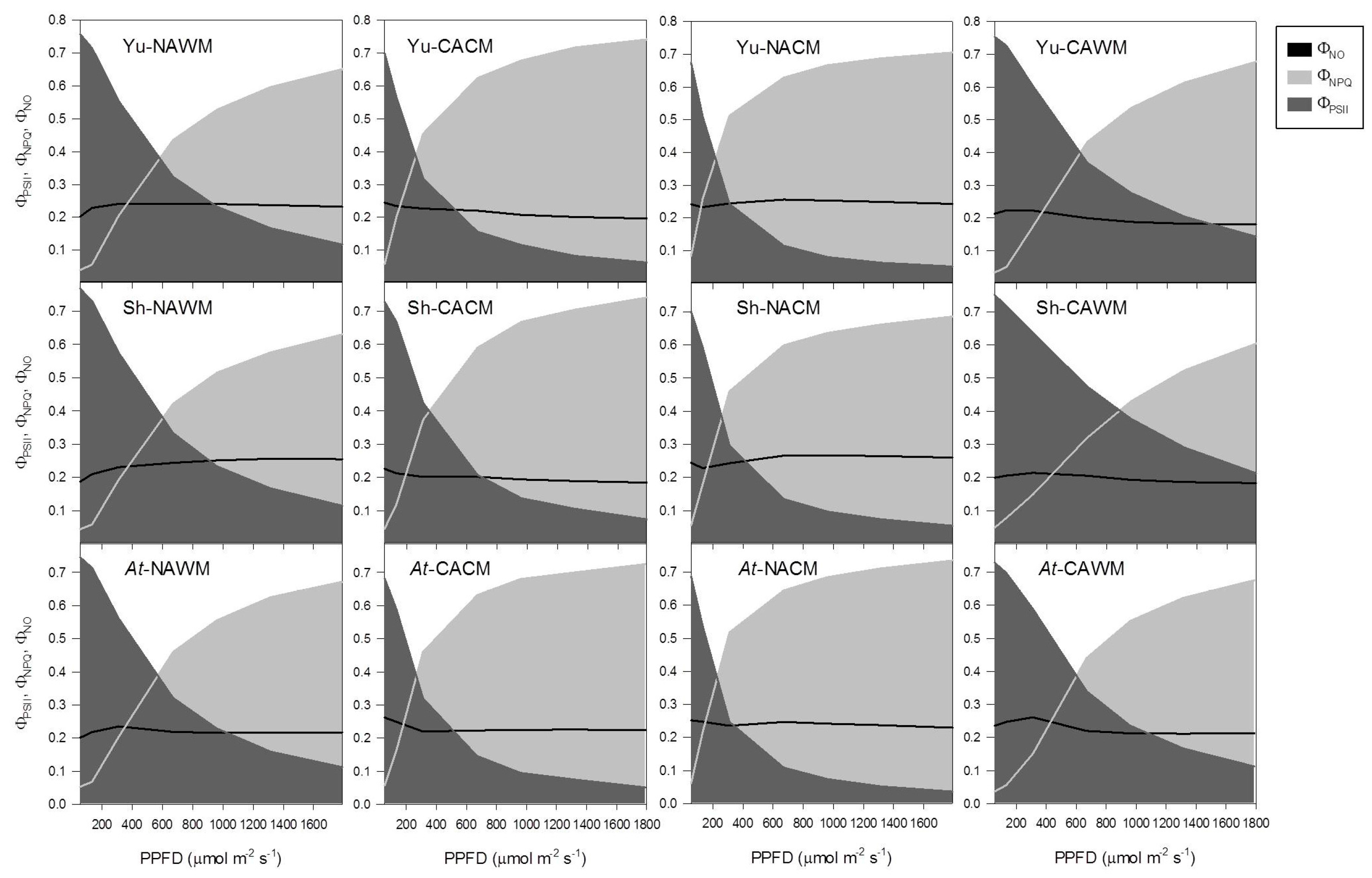
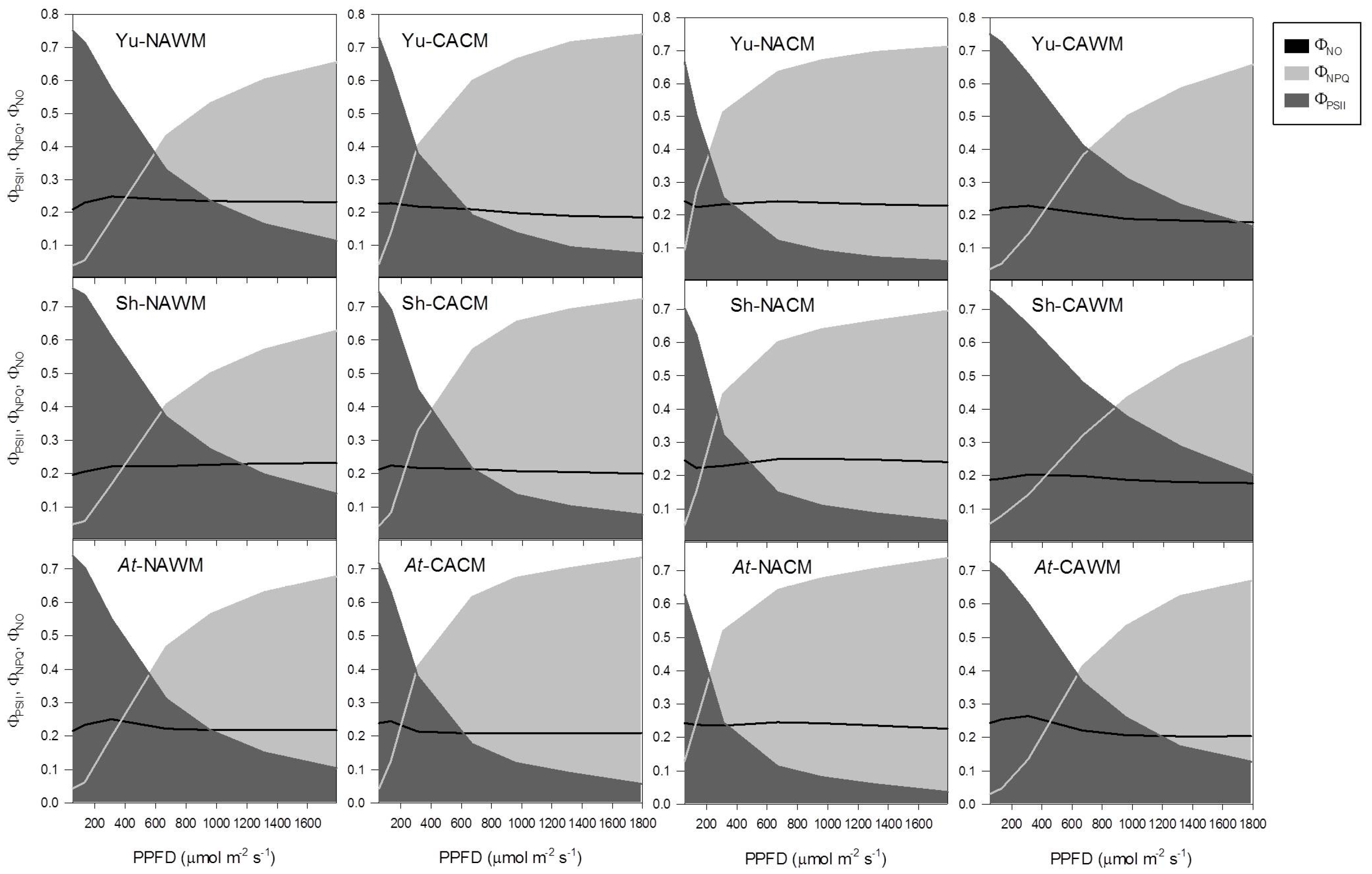
2.5.4. Excitation Energy Partitioning with Increasing Irradiance
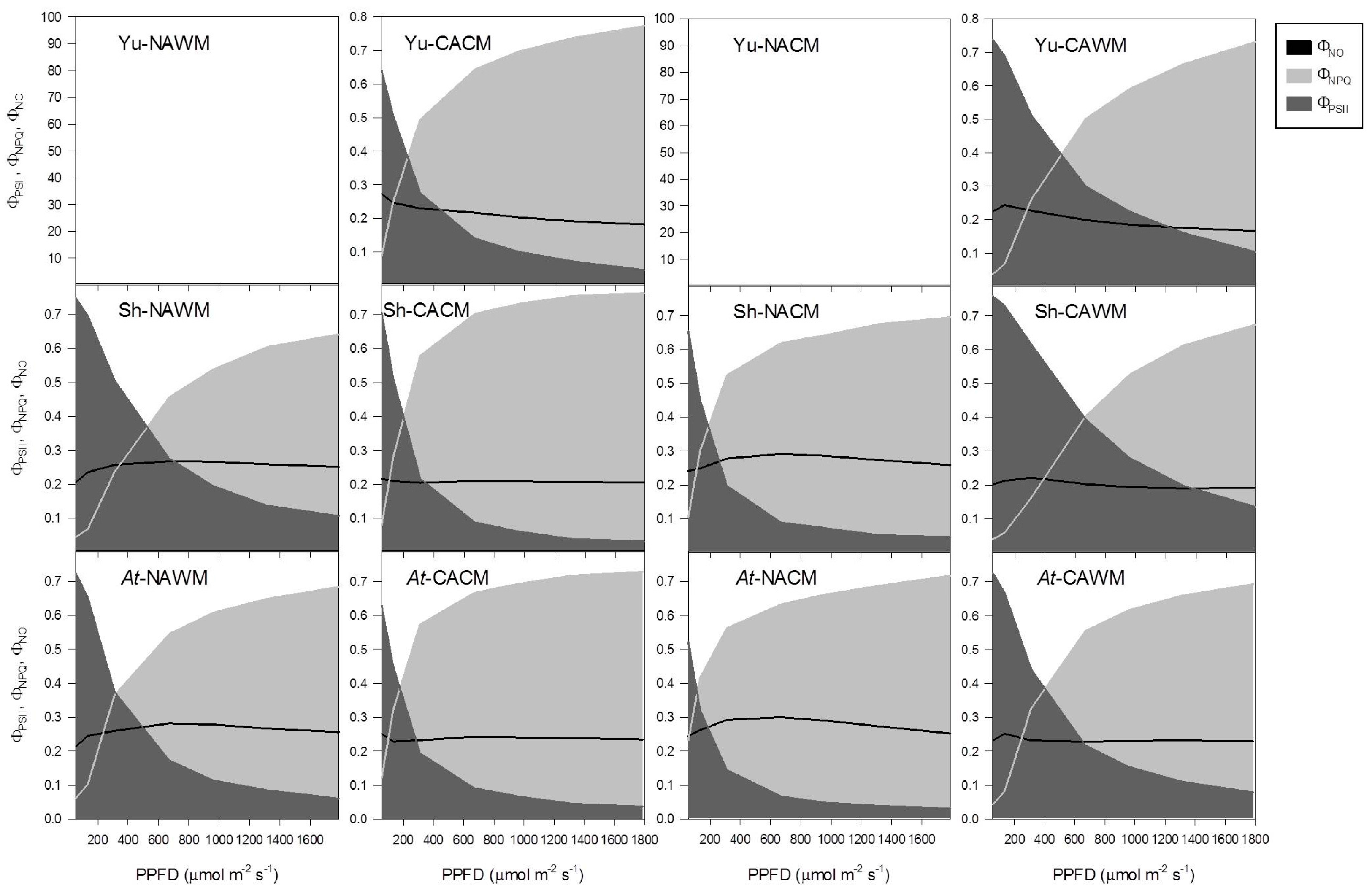
2.5.5. Excitation Energy Partitioning with Increasing Excitation Pressure
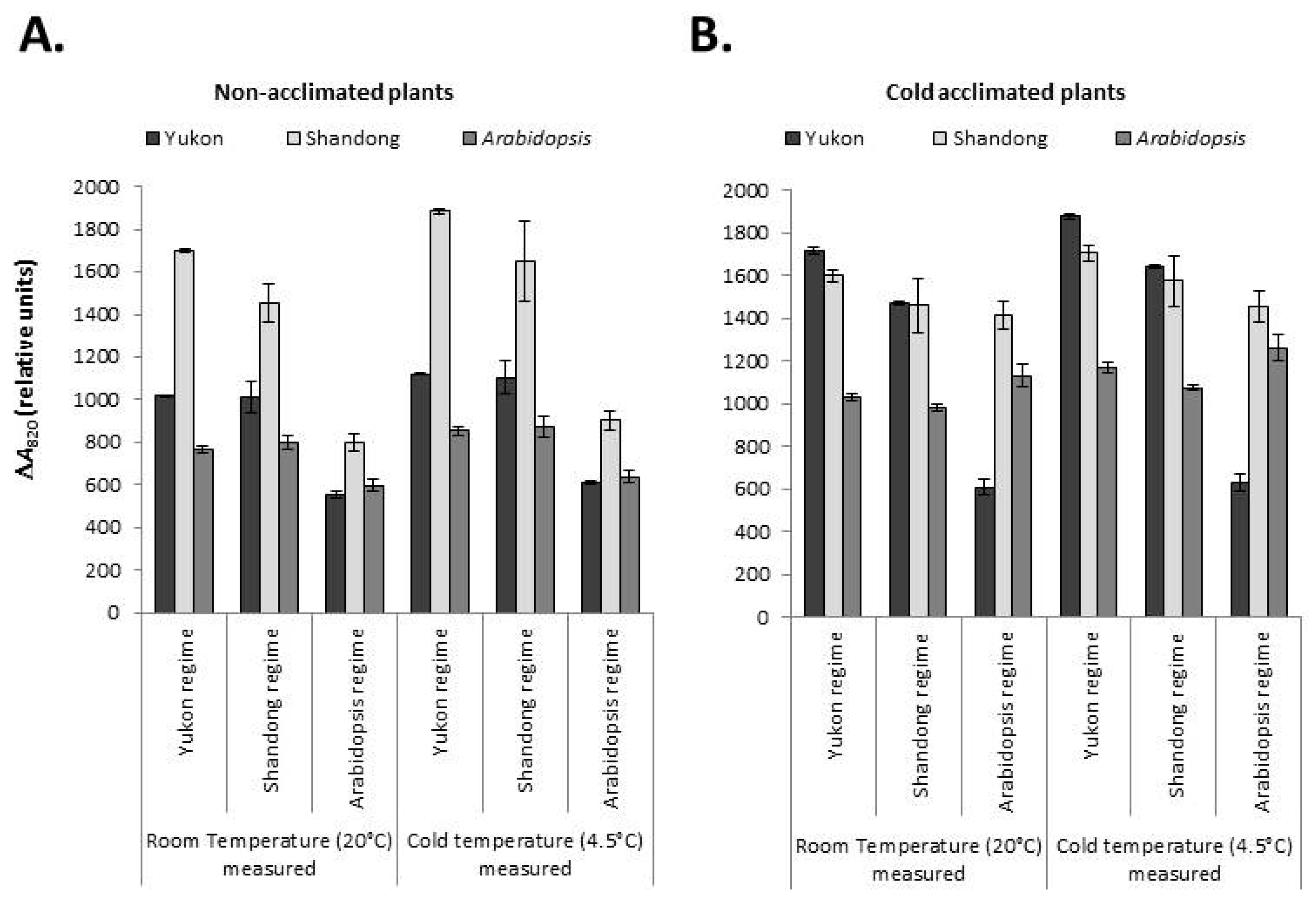
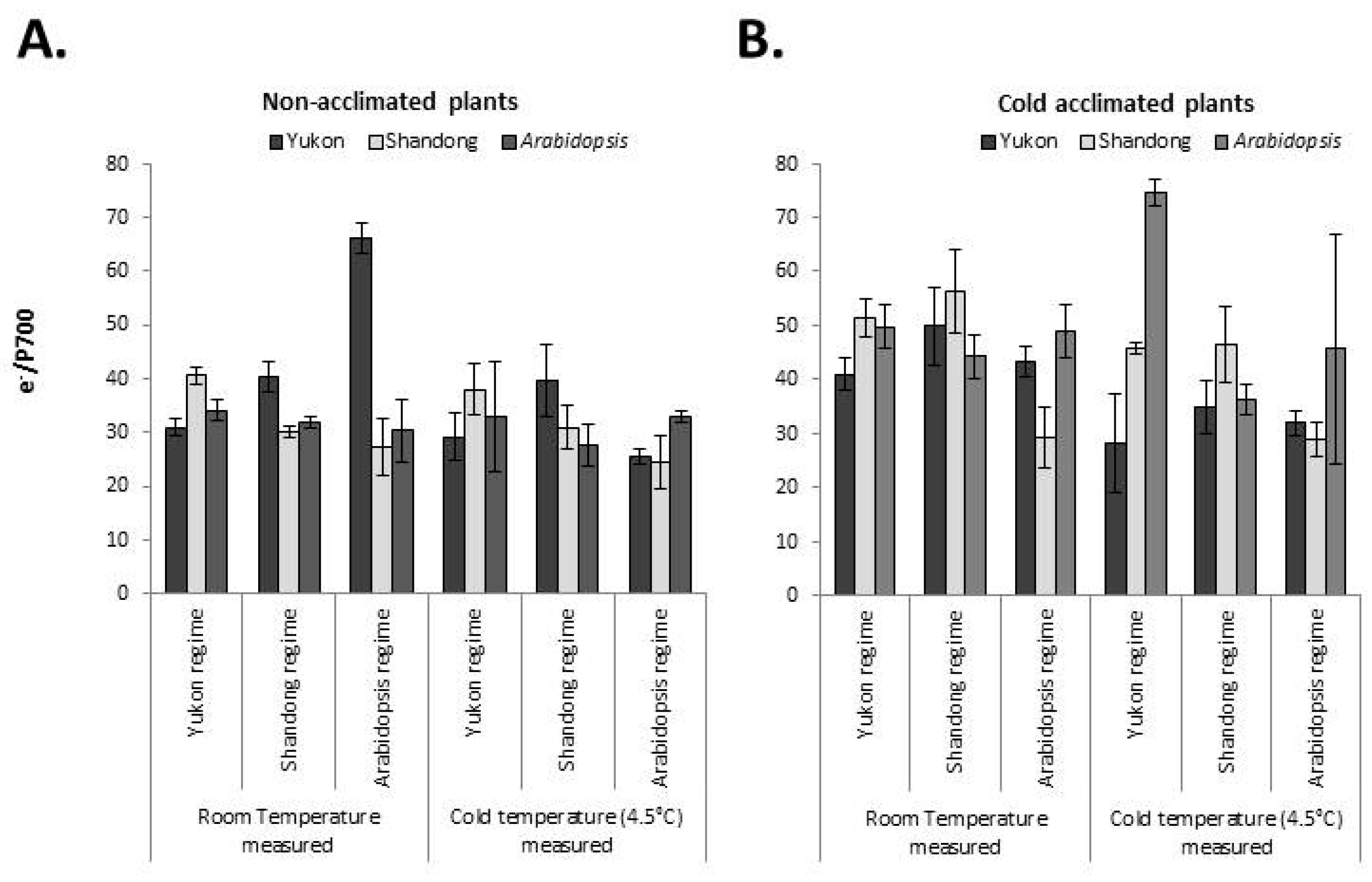
2.6. Redox State of PSI and the Intersystem Electron Pool
2.6.1. PSI Oxidation
2.6.2. Intersystem Electron Pool
3. Discussion
3.1. Differential Photoprotective Stratagies Indicated by Pigmentation
3.2. Similar Trends but Quantitative Differences in PSII Fluorescence Parameters
3.3. Differential Responses of Eutrema and Arabidopsis Plants to PSII Excitation Pressure
3.4. Yukon, Shandong and Arabidopsis Plants Show Divergent Trends in PSI Performance
4. Materials and Methods
4.1 Plant Material and Growth Conditions
4.2. Chlorophyll a Fluorescence
4.2.1. Steady-State Fluorescence Quenching
4.2.2. Chlorophyll Fluorescence Imaging
4.3. Photoinhibitory Treatments and Recovery
4.4. Photosystem I Spectroscopy
4.5. Photosynthetic Pigment Determination
4.6. Experimental Design and Statistical Analyses
5. Conclusions
5.1. Comparisons between Eutrema and Arabidopsis
5.2. Comparisons between Eutrema Accessions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Melis, A. Photostasis in Plants; Plenum Press: New York, NY, USA, 1998; pp. 207–220. [Google Scholar]
- Hüner, N.P.A.; Öquist, G.; Melis, A. Photostasis in Plants, Green Algae and Cyanobacteria: The Role of Light Harvesting Antenna Complexes; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; Volume 13, pp. 401–421. [Google Scholar]
- Ensminger, I.; Busch, F.; Huner, N.P.A. Photostasis and cold acclimation: Sensing low temperature through photosynthesis. Physiol. Plant. 2006, 126, 28–44. [Google Scholar] [CrossRef]
- Huner, N.P.A.; Maxwell, D.P.; Gray, G.R.; Savitch, L.V.; Krol, M.; Ivanov, A.G.; Falk, S. Sensing environmental change: PSII excitation pressure and redox signalling. Physiol. Plant. 1996, 98, 358–364. [Google Scholar] [CrossRef]
- Huner, N.P.A.; Öquist, G.; Sarhan, F. Energy balance and acclimation to light and cold. Trends Plant Sci. 1998, 3, 224–230. [Google Scholar] [CrossRef]
- Hüner, N.P.A.; Bode, R.; Dahal, K.; Busch, F.A.; Possmayer, M.; Szyszka, B.; Rosso, D.; Ensminger, I.; Krol, M.; Ivanov, A.G.; et al. Shedding some light on cold acclimation, cold adaptation, and phenotypic plasticity. Can. J. Bot. 2013, 91, 127–136. [Google Scholar] [CrossRef]
- Guy, C.L. Cold acclimation and freezing stress tolerance: Role of protein metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 187–223. [Google Scholar] [CrossRef]
- Guy, C.L.; Kaplan, F.; Kopka, J.; Selbig, J.; Hincha, D.K. Metabolomics of temperature stress. Physiol. Plant. 2008, 132, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. Molecular basis of plant cold acclimation: Insights gained from studying the CBF cold response pathway. Plant Physiol. 2010, 154, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, A.; Clément, C.; Barka, E.A. Physiological and molecular changes in plants grown at low temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Huner, N.P.A.; Öquist, G.; Hurry, V.M.; Krol, M.; Falk, S.; Griffith, M. Photosynthesis, photoinhibition and low temperature acclimation in cold tolerant plants. Photosynth. Res. 1993, 317, 9–39. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.R.; Heath, D. A global reorganization of the metabolome in Arabidopsis during cold acclimation is revealed by metabolic fingerprinting. Physiol. Plant. 2005, 124, 236–248. [Google Scholar] [CrossRef]
- Powles, S.B. Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol. 1984, 35, 15–44. [Google Scholar] [CrossRef]
- Aro, E.M.; Virgin, I.; Andersson, B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Scheller, H.; Haldrup, A. Photoinhibition of photosystem I. Planta 2005, 221, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Adams, W.W. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A.V.; Walters, R.G. Regulation of light harvesting in green plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 655–684. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.G.; Sane, P.V.; Hurry, V.; Öquist, G.; Huner, N.P.A. Photosystem II reaction centre quenching: Mechanisms and physiological role. Photosynth. Res. 2008, 98, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M.; Hurry, V. A plant for all seasons: Alterations in photosynthetic carbon metabolism during cold acclimation in Arabidopsis. Curr. Opin. Plant Biol. 2002, 5, 199–206. [Google Scholar] [CrossRef]
- Osmond, B.; Badger, M.; Maxwell, K.; Björkman, O.; Leegood, R. Too many photons: Photorespiration, photoinhibition and photooxidation. Trends Plant Sci. 1997, 2, 119–121. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Rosso, D.; Savitch, L.V.; Stachula, P.; Rosembert, M.; Öquist, G.; Hurry, V.; Hüner, N.P.A. Implications of alternative electron sinks in increased resistance of PSII and PSI photochemistry to high light stress in cold acclimated Arabidopsis thaliana. Photosynth. Res. 2012, 113, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Ort, D.R. When there is too much light. Plant Physiol. 2001, 125, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Öquist, G.; Hüner, N.P.A. Photosynthesis of overwintering evergreen plants. Annu. Rev. Plant Biol. 2003, 54, 329–355. [Google Scholar] [CrossRef] [PubMed]
- Amtmann, A. Learning from evolution: Thellungiella generates new knowledge on essential and critical components of abiotic stress tolerance in plants. Mol. Plant 2009, 2, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.H.; Dassanayake, M.; Bohnert, H.J.; Cheeseman, J.M. Life at the extreme: Lessons from the genome. Genome Biol. 2012, 13, 241. [Google Scholar] [CrossRef] [PubMed]
- Bressan, R.A.; Park, H.C.; Orsini, F.; Oh, D.H.; Dassanayake, M.; Inan, G.; Yun, D.J.; Bohnert, H.J.; Maggio, A. Biotechnology for mechanisms that counteract salt stress in extremophile species: A genome-based view. Plant Biotechnol. Rep. 2013, 7, 27–37. [Google Scholar] [CrossRef]
- Koch, M.A.; German, D.A. Taxonomy and systematics are key to biological information: Arabidopsis, Eutrema (Thellungiella), Noccaea and Schrenkiella (Brassicaceae) as examples. Front. Plant Sci. 2013, 4, 267. [Google Scholar] [CrossRef] [PubMed]
- Inan, G.; Zhang, Q.; Li, P.; Wang, Z.; Cao, Z.; Zhang, H.; Zhang, C.; Quist, T.M.; Goodwin, S.M.; Zhu, J.; et al. Salt cress: A halophyte and cryophyte Arabidopsis relative model system and it applicability to molecular genetic analyses of growth and development of extremophiles. Plant Physiol. 2004, 135, 1718–1737. [Google Scholar] [CrossRef] [PubMed]
- Taji, T.; Seki, M.; Satou, M.; Sakurai, T.; Kobayashi, M.; Ishiyama, K.; Narusaka, Y.; Narusaka, M.; Zhu, J.K.; Shinozaki, K. Comparative genomics in salt tolerance between Arabidopsis and Arabidopsis-related halophyte salt cress using Arabidopsis microarray. Plant Physiol. 2004, 135, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.Q.; Li, P.H.; Ma, S.S.; Rupassara, S.I.; Bohnert, H.J. Salinity stress adaptation competence in the extremophile Thellungiella halophila in comparison with its relative Arabidopsis thaliana. Plant J. 2005, 44, 826–839. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.; Timonin, M.; Wong, C.E.; Gray, G.R.; Akhter, S.R.; Saldanha, M.; Rogers, M.A.; Weretilnyk, E.A.; Moffatt, B. Thellungiella: An Arabidopsis-related model plant adapted to cold temperatures. Plant Cell Environ. 2007, 30, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Bi, Y.M.; Weretilnyk, E.; Barak, S.; Rothstein, S.J. The Arabidopsis halophytic relative Thellungiella halophila tolerates nitrogen-limiting conditions by maintaining growth, nitrogen uptake, and assimilation. Plant Physiol. 2008, 147, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Lugan, R.; Niogret, M.F.; Leport, L.; Guegan, J.P.; Larher, F.R.; Savouré, A.; Kopka, J.; Bouchereau, A. Metabolome and water homeostasis analysis of Thellungiella salsuginea suggests that dehydration tolerance is a key response to osmotic stress in this halophyte. Plant J. 2010, 64, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Guevara, D.R.; Champigny, M.J.; Tattersall, A.; Dedrick, J.; Wong, C.E.; Li, Y.; Labbe, A.; Ping, C.L.; Wang, Y.; Nuin, P.; et al. Transcriptomic and metabolomic analysis of Yukon Thellungiella plants grown in cabinets and their natural habitat show phenotypic plasticity. BMC Plant Biol. 2012, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.P.; Funk, C.; Erban, A.; Kopka, J.; Köhl, K.I.; Zuther, E.; Hincha, D.K. Salt stress responses in a geographically diverse collection of Eutrema/Thellungiella spp. accessions. Funct. Plant Biol. 2016, 43, 590–606. [Google Scholar]
- Bravo, L.A.; Saavedra-Mella, F.A.; Vera, F.; Guerra, A.; Cavieres, L.A.; Ivanov, A.G.; Huner, N.P.A.; Corcuera, L.J. Effect of cold acclimation on the photosynthetic performance of two ecotypes of Colobanthus quitensis (Kunth) Bartl. J. Exp. Bot. 2007, 58, 3581–3590. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Noguchi, K.; Hikosaka, K.; Terashima, I. Phenotypic plasticity in photosynthetic temperature acclimation among crop species with different cold tolerances. Plant Physiol. 2010, 152, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.P.; Babakov, A.; de Boer, B.; Zuther, E.; Hincha, D.K. Comparison of freezing tolerance,compatible solutes and polyamines in geographically diverse collections of Thellungiella sp. and Arabidopsis thaliana accessions. BMC Plant Biol. 2012, 12, 131. [Google Scholar] [CrossRef] [PubMed]
- Khanal, N.; Moffatt, B.A.; Gray, G.R. Acquisition of freezing tolerance in Arabidopsis and two contrasting ecotypes of the extremophile Eutrema salsugineum (Thellungiella salsuginea). J. Plant Physiol. 2015, 180, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zhou, Y.; Zhu, W.; Li, X.; Fan, L.; Zhang, G. Proteomic analysis of cold stress-responsive proteins in Thellungiella rosette leaves. Planta 2009, 230, 1033–1046. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.E.; Li, Y.; Whitty, B.; Akhter, S.; Diaz, C.; Brandle, J.; Golding, B.; Weretinylk, E.; Moffatt, B.A.; Griffith, M. Expressed sequence tags from the Yukon ecotype of Thellungiella salsuginea reveal that gene expression in response to cold, drought and salinity shows little overlap. Plant Mol. Biol. 2005, 58, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.E.; Li, Y.; Labbe, A.; Guevara, D.; Nuin, P.; Whitty, B.; Diaz, C.; Golding, G.B.; Gray, G.R.; Weretilnyk, E.A.; et al. Transcriptional profiling implicates novel interactions between abiotic stress and hormonal responses in Thellungiella, a close relative of Arabidopsis. Plant Physiol. 2006, 140, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- Champigny, M.J.; Sung, W.W.L.; Catana, V.; Salwan, R.; Summers, P.S.; Dudley, S.A.; Provart, N.J.; Cameron, R.K.; Golding, G.B.; Weretilnyk, E.A. RNA-Seq effectively monitors gene expression in Eutrema salsugineum plants growing in an extreme natural habitat and in controlled growth cabinet conditions. BMC Genom. 2013, 14, 578. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.P.; Giorgi, F.M.; Lohse, M.; Kvederaviciute, K.; Klages, S.; Usadel, B.; Meskiene, I.; Reinhardt, R.; Hincha, D.K. Transcriptome sequencing and microarray design for functional genomics in the extremophile Arabidopsis relative Thellungiella salsuginea (Eutrema salsugineum). BMC Genom. 2013, 14, 793. [Google Scholar] [CrossRef] [PubMed]
- Stepien, P.; Johnson, G.N. Contrasting responses of photosynthesis to salt stress in the glycophytic Arabidopsis and the halophytic Thellungiella: Role of the plastid terminal oxidase as an alternative electron sink. Plant Physiol. 2009, 149, 1154–1165. [Google Scholar] [CrossRef] [PubMed]
- Sui, N.; Han, G. Salt-induced photoinhibition of PSII is alleviated in halophyte Thellungiella halophila by increases of unsaturated fatty acids in membrane lipids. Acta Physiol. Plant. 2014, 36, 983–992. [Google Scholar] [CrossRef]
- Malik, V.M.; Lobo, J.M.; Stewart, C.; Irani, S.; Todd, C.D.; Gray, G.R. Growth irradiance affects ureide accumulation and tolerance to photoinhibition in Eutrema salsugineum (Thellungiella salsuginea). Photosynthetica 2016, 54, 93–100. [Google Scholar] [CrossRef]
- Nishio, J.N. Why are higher plants green? Evolution of the higher plant photosynthetic pigment complement. Plant Cell Environ. 2000, 6, 539–548. [Google Scholar] [CrossRef]
- M’rah, S.; Ouerghi, Z.; Berthomieu, C.; Havaux, M.; Jungas, C.; Hajji, M.; Grignon, C.; Lachaal, M. Effects of NaCl on the growth, ion accumulation and photosynthetic parameters of Thellungiella halophila. J. Plant Physiol. 2006, 163, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Savitch, L.V.; Barker-Astrom, J.; Ivanov, A.G.; Hurry, V.; Öquist, G.; Huner, N.P.A.; Gardeström, P. Cold acclimation of Arabidopsis thaliana results in incomplete recovery of photosynthetic capacity, associated with an increased reduction of the chloroplast stroma. Planta 2001, 214, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Bräutigam, K.; Dietzel, L.; Kleine, T.; Ströher, E.; Wormuth, D.; Dietz, K.J.; Radke, D.; Wirtz, M.; Hell, R.; Dörmann, P.; et al. Dynamic plastid redox signals integrate gene expression and metabolism to induce distinct metabolic states in photosynthetic acclimation in Arabidopsis. Plant Cell 2009, 21, 2715–2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The dynamics of photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef] [PubMed]
- Harbinson, J.; Hedley, C.L. Changes in P700 oxidation during the early stages of the induction of photosynthesis. Plant Physiol. 1993, 103, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Asada, K.; Heber, U.; Schreiber, U. Pool size of electrons that can be donated to P700+, as determined in intact leaves: Donation to P700+ from stromal components via the intersystem chain. Plant Cell Physiol. 1992, 33, 927–932. [Google Scholar]
- Bukhov, N.G.; Samson, G.; Carpentier, R. Non-photosynthetic reduction of the intersystem electron transport chain of chloroplasts following heat stress: The pool size of stromal reductants. Photochem. Photobiol. 2001, 74, 438–443. [Google Scholar] [CrossRef]
- Gray, G.R.; Hope, B.J.; Qin, X.Q.; Tayler, B.G.; Whitehead, C.L. The characterization of photoinhibition and recovery during cold acclimation in Arabidopsis thaliana using chlorophyll fluorescence imaging. Physiol. Plant. 2003, 119, 365–375. [Google Scholar] [CrossRef]
- Hendrickson, L.; Furbank, R.T.; Chow, W.S. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence. Photosynth. Res. 2004, 82, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Schreiber, U. Energy-dependent quenching of dark-level chlorophyll fluorescence in intact leaves. Photosynth. Res. 1986, 10, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll Fluorescence as a Nonintrusive Indicator for Rapid Assessment of In Vivo Photosynthesis; Spinger-Verlag: Berlin, Germany, 1994; pp. 49–70. [Google Scholar]
- Gray, G.R.; Ivanov, A.G.; Krol, M.; Huner, N.P.A. Adjustment of thylakoid plastoquinone content and Photosystem I electron donor pool size in response to growth temperature and growth irradiance in winter rye (Secale cereale L.). Photosynth. Res. 1998, 56, 209–221. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Regime | Acclimation Status | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Non-Acclimated | Cold Acclimated | |||||||||
| Temperature (Light/Dark) | Photoperiod (Light/Dark) | PPFD (μmol photons m−2 s−1) | DPR (μmol photons m−2) | DAT (°C) | Temperature (Light/Dark) | Photoperiod (Light/Dark) | PPFD (μmol photons m−2 s−1) | DPR (μmol photons m−2) | DAT (°C) | |
| Yukon | 22/10 °C | 21/3 h | 250 | 18.90 | 20.5 | 5/4 °C | 21/3 h | 250 | 18.90 | 4.9 |
| Shandong | 22/19 °C | 16/8 h | 250 | 14.40 | 21.0 | 5/4 °C | 16/8 h | 250 | 14.40 | 4.7 |
| Arabidopsis | 20/20 °C | 16/8 h | 100 | 5.75 | 20.0 | 5/4 °C | 16/8 h | 100 | 5.75 | 4.7 |
| Parameter | Taxa (Accession) | Growth Regime | |||||
|---|---|---|---|---|---|---|---|
| Yukon | Shandong | Arabidopsis | |||||
| Non-Acclimated | Cold Acclimated | Non-Acclimated | Cold Acclimated | Non-Acclimated | Cold Acclimated | ||
| ChlFW (μg mg−1) | Eutrema (Yukon) | 2.4 ± 0.07 a,A | 1.9 ± 0.08 a,B | 2.4 ± 0.10 a,A | 1.9 ± 0.05 a,B | 2.4±0.29 a,A | 2.2 ± 0.04 a,A |
| Eutrema (Shandong) | 2.2 ± 0.02 a,A | 1.6 ± 0.03 b,B | 2.2 ± 0.11 a,A | 1.6 ± 0.02 b,B | 2.5±0.03 a,A | 1.1 ± 0.21 c,B | |
| Arabidopsis (Columbia) | 1.3 ± 0.05 b,A | 1.3 ± 0.06 c,A | 1.2 ± 0.07 b,A | 1.5 ± 0.03 c,A | 1.7±0.05 b,A | 1.6 ± 0.07 b,A | |
| CarFW (μg mg−1) | Eutrema (Yukon) | 0.36 ± 0.009 a,B | 0.46 ± 0.010 a,A | 0.40 ± 0.005 a,B | 0.49 ± 0.012 a,A | 0.41 ± 0.047 a,A | 0.48 ± 0.022 a,A |
| Eutrema (Shandong) | 0.43 ± 0.003 a,A | 0.35 ± 0.010 b,B | 0.34 ± 0.016 b,A | 0.37 ± 0.008 b,A | 0.38 ± 0.008 a,A | 0.30 ± 0.053 b,A | |
| Arabidopsis (Columbia) | 0.23 ± 0.010 b,B | 0.33 ± 0.010 b,A | 0.20 ± 0.013 c,B | 0.29 ± 0.012 c,A | 0.29 ± 0.021 b,A | 0.33 ± 0.013 b,A | |
| ChlLA (μg cm−2) | Eutrema (Yukon) | 64.5 ± 5.22 a,A | 63.3 ± 1.35 b,A | 64.1 ± 2.45 a,A | 65.7 ± 4.08 a,A | 39.0 ± 2.2 b,B | 62.0 ± 2.48 a,A |
| Eutrema (Shandong) | 72.2 ± 2.69 a,A | 71.3 ± 2.67 a,A | 71.3 ± 4.05 a,A | 78.1 ± 4.82 a,A | 61.7 ± 1.84 a,A | 32.8 ± 5.80 b,B | |
| Arabidopsis (Columbia) | 36.1 ± 2.38 b,A | 38.3 ± 2.35 c,A | 26.8 ± 1.26 b,B | 46.8 ± 1.46 b,A | 32.9 ± 1.26 c,B | 42.6 ± 1.30 b,A | |
| CarLA (μg cm−2) | Eutrema (Yukon) | 9.9 ± 0.64 b,B | 15.4 ± 0.58 a,A | 10.8 ± 0.64 a,B | 17.3 ± 0.57 a,A | 6.7 ± 0.32 b,B | 13.5 ± 0.36 a,A |
| Eutrema (Shandong) | 14.1 ± 0.44 a,A | 15.4 ± 0.43 a,A | 11.2 ± 0.73 a,B | 18.1 ± 1.16 a,A | 9.4 ± 0.14 a,A | 8.9 ± 1.47 b,A | |
| Arabidopsis (Columbia) | 6.5 ± 0.34 c,B | 9.5 ± 0.43 b,A | 4.4 ± 0.24 b,B | 9.0 ± 0.27 b,A | 5.5 ± 0.19 c,B | 8.7 ± 0.23 b,A | |
| Chl a:b | Eutrema (Yukon) | 3.1 ± 0.13 c,B | 4.9 ± 0.50 a,A | 3.3 ± 0.29 a,B | 4.8 ± 0.06 a,A | 2.8 ± 0.15 b,A | 3.8 ± 0.39 a,A |
| Eutrema (Shandong) | 4.7 ± 0.12 a,A | 4.2 ± 0.10a b,A | 3.6 ± 0.20 a,B | 4.8 ± 0.22 a,A | 3.2 ± 0.14 b,A | 2.2 ± 0.30 b,B | |
| Arabidopsis (Columbia) | 3.9 ± 0.05 b,A | 3.7 ± 0.37 b,A | 4.0 ± 0.08 a,A | 3.3 ± 0.24 b,B | 3.9 ± 0.22 a,A | 4.1 ± 0.11 a,A | |
| Chl:Car | Eutrema (Yukon) | 6.5 ± 0.21 a,A | 4.1 ± 0.16 a,B | 6.0 ± 0.28 a,A | 3.8 ± 0.14 b,B | 5.9 ± 0.41 a,A | 4.6 ± 0.29 a,A |
| Eutrema (Shandong) | 5.1 ± 0.04 b,A | 4.6 ± 0.12 a,A | 6.4 ± 0.27 a,A | 4.3 ± 0.10 b,B | 6.6 ± 0.16 a,A | 3.7 ± 0.20 b,B | |
| Arabidopsis (Columbia) | 5.5 ± 0.23 b,A | 4.0 ± 0.24 a,B | 6.1 ± 0.05 a,A | 5.2 ± 0.28 a,A | 6.0 ± 0.28 a,A | 4.9 ± 0.08 a,B | |
| Parameters | Cold Shock Effect (NAWM-NACM)/NAWM | Cold Acclimative Effect (CACM-NACM)/NACM | Relaxation/Augmentation Effect (CAWM-CACM)/CACM | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Yukon | Shandong | Arabidopsis | Yukon | Shandong | Arabidopsis | Yukon | Shandong | Arabidopsis | |
| 1-qL | −0.51 | −0.42 | −0.51 | −0.17 | −0.13 | −0.20 | −0.58 | −0.48 | −0.52 |
| RETRPSII | 0.54 | 0.49 | 0.57 | 0.34 | 0.32 | 0.40 | 1.05 | 1.18 | 0.94 |
| qO | −0.39 | −0.26 | −0.99 | 0.16 | 0.13 | 0.30 | −0.50 | −0.52 | −0.41 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khanal, N.; Bray, G.E.; Grisnich, A.; Moffatt, B.A.; Gray, G.R. Differential Mechanisms of Photosynthetic Acclimation to Light and Low Temperature in Arabidopsis and the Extremophile Eutrema salsugineum. Plants 2017, 6, 32. https://doi.org/10.3390/plants6030032
Khanal N, Bray GE, Grisnich A, Moffatt BA, Gray GR. Differential Mechanisms of Photosynthetic Acclimation to Light and Low Temperature in Arabidopsis and the Extremophile Eutrema salsugineum. Plants. 2017; 6(3):32. https://doi.org/10.3390/plants6030032
Chicago/Turabian StyleKhanal, Nityananda, Geoffrey E. Bray, Anna Grisnich, Barbara A. Moffatt, and Gordon R. Gray. 2017. "Differential Mechanisms of Photosynthetic Acclimation to Light and Low Temperature in Arabidopsis and the Extremophile Eutrema salsugineum" Plants 6, no. 3: 32. https://doi.org/10.3390/plants6030032
APA StyleKhanal, N., Bray, G. E., Grisnich, A., Moffatt, B. A., & Gray, G. R. (2017). Differential Mechanisms of Photosynthetic Acclimation to Light and Low Temperature in Arabidopsis and the Extremophile Eutrema salsugineum. Plants, 6(3), 32. https://doi.org/10.3390/plants6030032