
Litter Accumulation and Nutrient Content of Roadside Plant Communities in Sichuan Basin, China


Abstract
:1. Introduction
2. Results
2.1. Sampling Plot Variation in Canopy Cover of Primary Life Forms
2.2. Principal Component Analysis of Litter Characteristics
2.3. Plant Species Correlation with PCA Axes
3. Discussion
4. Materials and Methods
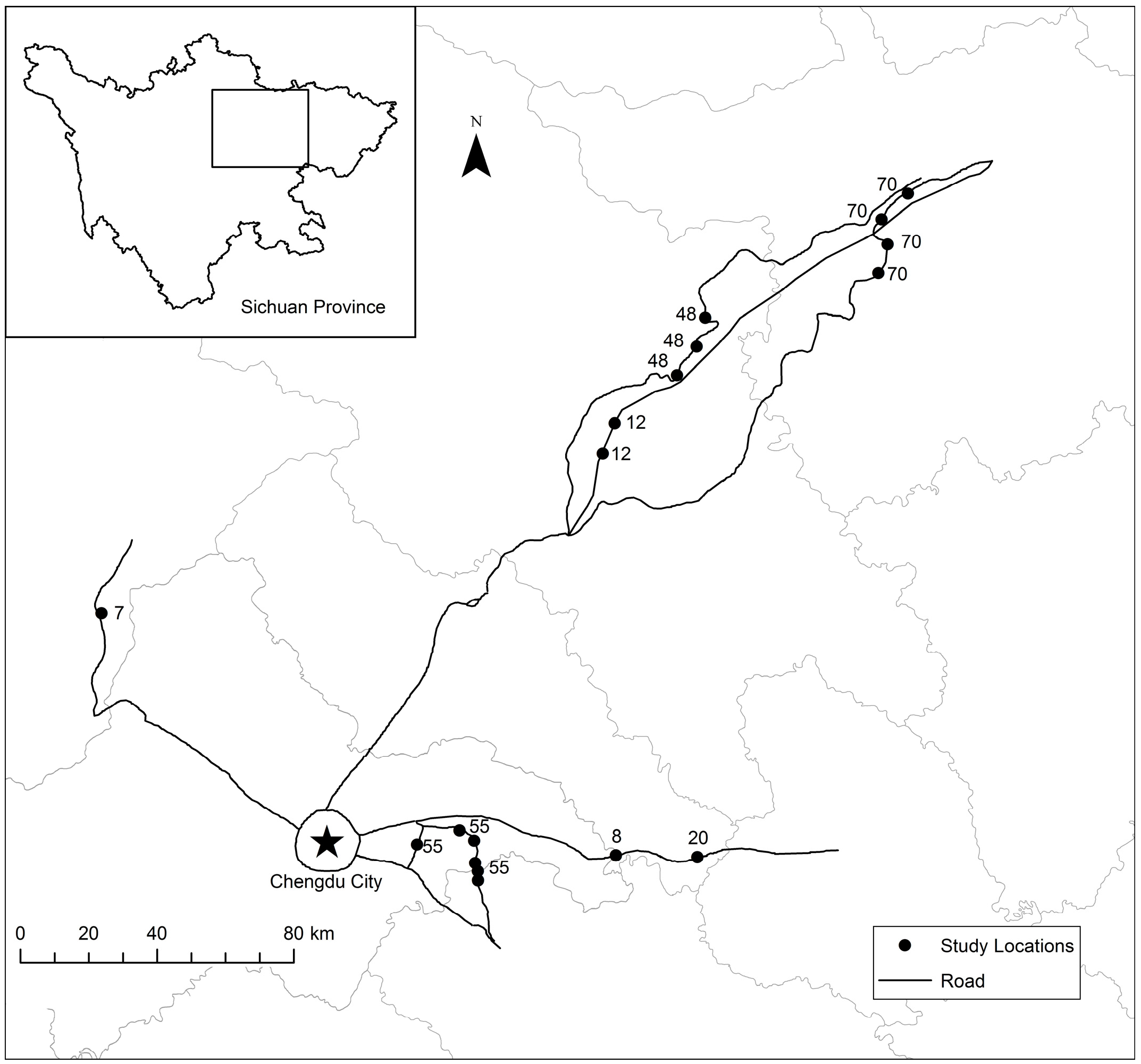
4.1. Vegetation Sampling
4.2. Litter Sampling
4.3. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Powers, J.S.; Montgomery, R.A.; Adair, E.C.; Brearley, F.Q.; DeWalt, S.J.; Castanho, C.T.; Chave, J.; Deinert, E.; Ganzhorn, J.U.; Gilbert, M.E.; et al. Decomposition in tropical forests: A pan-tropical study of the effects of litter type, litter placement and mesofaunal exclusion across a precipitation gradient. J. Ecol. 2009, 97, 801–811. [Google Scholar] [CrossRef]
- Li, D.; Peng, S.; Chen, B. The effects of leaf litter evenness on decomposition depend on which plant functional group is dominant. Plant Soil 2013, 365, 255–266. [Google Scholar] [CrossRef]
- Loydi, A.; Eckstein, R.L.; Otte, A.; Donath, T.W. Effects of litter on seedling establishment in natural and semi-natural grasslands: A meta-analysis. J. Ecol. 2013, 101, 454–464. [Google Scholar] [CrossRef]
- Santos, S.L.D.; Valio, I.F.M. Litter accumulation and its effect on seedling recruitment in a southeast brazilian tropical forest. Braz. J. Bot. 2002, 25, 89–92. [Google Scholar] [CrossRef]
- Villalobos-Vega, R.; Goldstein, G.; Haridasan, M.; Franco, A.C.; Miralles-Wilhelm, F.; Scholz, F.G.; Bucci, S.J. Leaf litter manipulations alter soil physicochemical properties and tree growth in a neotropical savanna. Plant Soil 2011, 346, 385. [Google Scholar] [CrossRef]
- Tiegs, S.D.; Akinwole, P.O.; Gessner, M.O. Litter decomposition across multiple spatial scales in stream networks. Oecologia 2009, 161, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Engber, E.A.; Varner, J.M.; Arguello, L.A.; Suihara, N.G. The effects of conifer encroachment and overstory structure on fuels and fire in an oak woodland landscape. Fire Ecol. 2012, 7, 32–50. [Google Scholar] [CrossRef]
- Wood, T.E.; Lawrence, D.; Clark, D.A.; Chazdon, R.L. Rain forest nutrient cycling and productivity in response to large-scale litter manipulation. Ecology 2009, 90, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Fahey, T.J.; Yavitt, J.B.; Sherman, R.E.; Groffman, P.M.; Fisk, M.C.; Maerz, J.C. Transport of carbon and nitrogen between litter and soil organic matter in a northern hardwood forest. Ecosystems 2011, 14, 326–340. [Google Scholar] [CrossRef]
- Jacob, M.; Viedenz, K.; Polle, A.; Thomas, F.M. Leaf litter decomposition in temperate deciduous forest stands with a decreasing fraction of beech (Fagus sylvatica). Oecologia 2010, 164, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Sariyildiz, T.; Küçük, M. Influence of slope position, stand type and rhododendron (Rhododendron ponticum) on litter decomposition rates of oriental beech (Fagus orientalis Lipsky.) and spruce [Picea orientalis (L.) link]. Eur. J. For. Res. 2009, 128, 351–360. [Google Scholar] [CrossRef]
- Rowe, E.C.; Healey, J.R.; Edwards-Jones, G.; Hills, J.; Howells, M.; Jones, D.L. Fertilizer application during primary succession changes the structure of plant and herbivore communities. Biol. Conserv. 2006, 131, 510–522. [Google Scholar] [CrossRef]
- Hui, D.; Jackson, R.B. Assessing interactive responses in litter decomposition in mixed species litter. Plant Soil 2009, 314, 263–271. [Google Scholar] [CrossRef]
- Berger, T.W.; Berger, P. Does mixing of beech (Fagus sylvatica) and spruce (Picea abies) litter hasten decomposition? Plant Soil 2014, 377, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Schwilk, D.W.; Caprio, A.C. Scaling from leaf traits to fire behaviour: Community composition predicts fire severity in a temperate forest. J. Ecol. 2011, 99, 970–980. [Google Scholar] [CrossRef]
- Hale, B.; Robertson, P. Plant community and litter composition in temperate deciduous woodlots along two field gradients of soil ni, cu and co concentrations. Environ. Pollut. 2016, 212, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Camill, P.; McKone, M.J.; Sturges, S.T.; Severud, W.J.; Ellis, E.; Limmer, J.; Martin, C.B.; Navratil, R.T.; Purdie, A.J.; Sandel, B.S.; et al. Community- and ecosystem-leve changes in species-rich tallgrass prairie. Ecol. Appl. 2004, 14, 1690–1694. [Google Scholar] [CrossRef]
- Wu, D.; Li, T.; Wan, S. Time and litter species composition affect litter-mixing effects on decomposition rates. Plant Soil 2013, 371, 355–366. [Google Scholar] [CrossRef]
- Loydi, A.; Lohse, K.; Otte, A.; Donath, T.W.; Eckstein, R.L. Distribution and effects of tree leaf litter on vegetation composition and biomass in a forest–grassland ecotone. J. Plant Ecol. 2014, 7, 264–275. [Google Scholar] [CrossRef]
- Facelli, J.M.; Carson, W.P. Heterogeneity of plant litter accumulation in successional communities. Bull. Torrey Bot. Soc. 1991, 118, 62–66. [Google Scholar] [CrossRef]
- Bonet, A. Secondary succession of semi-arid mediterranean old-fields in south-eastern spain: Insights for conservation and restoration of degraded lands. J. Arid Environ. 2004, 56, 213–233. [Google Scholar] [CrossRef]
- Walck, J.L.; Baskin, J.M.; Baskin, C.C. Ecology of the endangered species Solidago shortii. Vi. Effects of habitat type, leaf litter, and soil type on seed germination. J. Torrey Bot. Soc. 1999, 126, 117–123. [Google Scholar] [CrossRef]
- Becerra, P.I.; Celis-Diez, J.L.; Bustamante, R.O. Effects of leaf litter and precipitation on germination and seedling survival of the endangered tree Beilschmiedia miersii. J. Veg. Sci. 2004, 7, 253–257. [Google Scholar] [CrossRef]
- Solly, E.F.; Schöning, I.; Boch, S.; Kandeler, E.; Marhan, S.; Michalzik, B.; Müller, J.; Zscheischler, J.; Trumbore, S.E.; Schrumpf, M. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. Plant Soil 2014, 382, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.H.; Wang, H.H.; Tsai, C.C.; Hseu, Z.Y. Litter production, decomposition and nutrient return of uplifted coral reef tropical forest. For. Ecol. Manag. 2006, 235, 174–185. [Google Scholar] [CrossRef]
- Ma, Y.; Filley, T.R.; Szlavecz, K.; McCormick, M.K. Controls on wood and leaf litter incorporation into soil fractions in forests at different successional stages. Soil Biol. Biochem. 2014, 69, 212–222. [Google Scholar] [CrossRef]
- Kazakou, E.; Violle, C.; Roumet, C.; Pintor, C.; Gimenez, O.; Garnier, E. Litter quality and decomposability of species from a mediterranean succession depend on leaf traits but not on nitrogen supply. Ann. Bot. 2009, 104, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- McLaren, J.R.; Turkington, R. Plant identity influences decomposition through more than one mechanism. PLoS ONE 2011, 6, e23702. [Google Scholar] [CrossRef] [PubMed]
- Scherer-Lorenzen, M. Functional diversity affects decomposition processes in experimental grasslands. Funct. Ecol. 2008, 22, 547–555. [Google Scholar] [CrossRef]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Quested, H.; Eriksson, O. Litter species composition influences the performance of seedlings of grassland herbs. Funct. Ecol. 2006, 20, 522–532. [Google Scholar] [CrossRef]
- Yilmaz, H.; Yilmaz, O.Y.; Akyuz, Y.F. Determining the factors affecting the distribution of muscari latifolium, an endemic plant of turkey, and a mapping species distribution model. Ecol. Evol. 2017, 7, 1112–1124. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.; Kinzig, A.; Landgridge, J. Plant attribute diversity, resilience, and ecosystem function: The nature and significance of dominant and minor species. Ecosystems 1999, 2, 95–113. [Google Scholar] [CrossRef]
- Jain, M.; Flynn, D.F.; Prager, C.M.; Hart, G.M.; Devan, C.M.; Ahrestani, F.S.; Palmer, M.I.; Bunker, D.E.; Knops, J.M.; Jouseau, C.F.; et al. The importance of rare species: A trait-based assessment of rare species contributions to functional diversity and possible ecosystem function in tall-grass prairies. Ecol. Evol. 2014, 4, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Jobidon, R.; Cyr, G.; Thiffault, N. Plant species diversity and composition along an experimental gradient of northern hardwood abundance in picea mariana plantations. For. Ecol. Manag. 2004, 198, 209–221. [Google Scholar] [CrossRef]
- Nakamura, T. Ecological relationships between seedling emergence and litter cover in the earliest stage of plant succession on sandy soil. Ecol. Res. 1996, 11, 105–110. [Google Scholar] [CrossRef]
- Spooner, P.G.; Lunt, I.D.; Briggs, S.V.; Freudenberger, D. Effects of soil disturbance from roadworks on roadside shrubs in a fragmented agricultural landscape. Biol. Conserv. 2004, 117, 393–406. [Google Scholar] [CrossRef]
- Agherkakli, B.; Najafi, A.; Sadeghi, S.H. Ground based operation effects on soil disturbance by steel tracked skidder in a steep slope of forest. J. For. Sci. 2010, 56, 278–284. [Google Scholar]
- He, H.; Li, S.; Sun, H.; Yang, T. Environmental factors of road slope stability in mountain area using principal component analysis and hierarchy cluster. Environ. Earth Sci. 2011, 62, 55–59. [Google Scholar] [CrossRef]
- Bansal, S.; Sheley, R.L.; Blank, B.; Vasquez, E.A. Plant litter effects on soil nutrient availability and vegetation dynamics: Changes that occur when annual grasses invade shrub-steppe communities. Plant Ecol. 2014, 215, 367–378. [Google Scholar] [CrossRef]
- Meier, C.L.; Bowman, W.D. Links between plant litter chemistry, species diversity, and below-ground ecosystem function. Proc. Natl. Acad. Sci. USA 2008, 105, 19780–19785. [Google Scholar] [CrossRef] [PubMed]
- Facelli, J.M.; Pickett, S.T.A. Plant litter—Its dynamics and effects on plant community structure. Bot. Rev. 1991, 57, 1–32. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Eppinga, M.B.; Kaproth, M.A.; Collins, A.R.; Molofsky, J. Litter feedbacks, evolutionary change and exotic plant invasion. J. Ecol. 2011, 99, 503–514. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Ravit, B.; Elgersma, K. Feedback in the plant-soil system. Ann. Rev. Environ. Res. 2005, 30, 75–115. [Google Scholar] [CrossRef]
- Xiong, S.J.; Nilsson, C. The effects of plant litter on vegetation: A meta-analysis. J. Ecol. 1999, 87, 984–994. [Google Scholar] [CrossRef]
- Dick, D.A.; Gilliam, F.S. Spatial heterogeneity and dependence of soils and herbaceous plant communities in adjacent seasonal wetland and pasture sites. Wetlands 2007, 27, 951–963. [Google Scholar] [CrossRef]
- Tang, F.K.; Cui, M.; Lu, Q.; Liu, Y.G.; Guo, H.Y.; Zhou, J.X. Effects of vegetation restoration on the aggregate stability and distribution of aggregate-associated organic carbon in a typical karst gorge region. Solid Earth 2016, 7, 141–151. [Google Scholar] [CrossRef]
- James, J.J. Leaf nitrogen productivity as a mechanism driving the success of invasive annual grasses under low and high nitrogen supply. J. Arid Environ. 2008, 72, 1775–1784. [Google Scholar] [CrossRef]
- Hossain, M.; Siddique, M.R.H.; Rahman, M.S.S.; Hossain, M.Z.; Hassan, M.M. Nutrient dynamics associated with leaf litter decomposition of three agroforestry tree species (Azadirachta indica, Dalbergia sissoo, and Melia azedarach) of bangladesh. J. For. Res. 2011, 22, 577–582. [Google Scholar] [CrossRef]
- Ghestem, M.; Cao, K.; Ma, W.; Rowe, N.; Leclerc, R.; Gadenne, C.; Stokes, A. A framework for identifying plant species to be used as ‘ecological engineers’ for fixing soil on unstable slopes. PLoS ONE 2014, 9, e95876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaston, K.J. The importance of being rare. Nature 2012, 487, 46–47. [Google Scholar] [CrossRef] [PubMed]
- Leitão, R.P.; Zuanon, J.; Villeger, S.; Williams, S.E.; Baraloto, C.; Fortunel, C.; Mendonca, F.P.; Mouillot, D. Rare species contribute disproportionately to the functional structure of species assemblages. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160084. [Google Scholar] [CrossRef] [PubMed]
- Mouillot, D.; Bellwood, D.R.; Baraloto, C.; Chave, J.; Galzin, R.; Harmelin-Vivien, M.; Kulbicki, M.; Lavergne, S.; Lavorel, S.; Mouquet, N.; et al. Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol. 2013, 11, e1001569. [Google Scholar] [CrossRef] [PubMed]
- Gascon, C.; Brooks, T.M.; Contreras-MacBeath, T.; Heard, N.; Konstant, W.; Lamoreux, J.; Launay, F.; Maunder, M.; Mittermeier, R.A.; Molur, S.; et al. The importance and benefits of species. Curr. Biol. 2015, 25, R431–R438. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, O.S.; Holt, A.R.; Warren, P.H.; Evans, K.L. Optimising uk urban road verge contributions to biodiversity and ecosystem services with cost-effective management. J. Environ. Manag. 2017, 191, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z. Characteristics and conservation priority of threatened plants in the yangtze valley. Biodivers. Conserv. 2003, 12, 65–72. [Google Scholar] [CrossRef]
- Karim, M.N.; Mallik, A.U. Roadside revegetation by native plants: I. Roadside microhabitats, floristic zonation and species traits. Ecol. Eng. 2008, 32, 222–237. [Google Scholar] [CrossRef]
- Hua, K.; Zhu, B.; Wang, X. Dissolved organic carbon loss fluxes through runoff and sediment on sloping upland of purple soil in the Sichuan Basin. Nutr. Cycl. Agroecosyst. 2014, 98, 125–135. [Google Scholar] [CrossRef]
- Wang, C.; Li, S.; Xia, J.; Wu, J. Clay composition and fertilizer conservation property of red soil and yellow soils in the southwest. J. Sichuan Agric. Univ. 1996, 2, 211–218. [Google Scholar]
- Li, Q.; Ma, M.; Liu, Y.; Ding, H.; Chen, M.; Chen, Y. Study on soil carbon and nutrients pools of several evergreen broad-leaved forest types in northwest sichuan. J. Soil Water Conserv. 2007, 6, 114–125. [Google Scholar]
- Peng, W.; Wang, G.; Zhou, J.; Zhao, J.; Yang, C. Studies on the temporal and spatial variations of urban expansion in chengdu, western china, from 1978 to 2010. Sustain. Cities Soc. 2015, 17, 141–150. [Google Scholar] [CrossRef]
- Jennings, S.B.; Brown, N.D.; Sheil, D. Assessing forest canopies and understorey illumination: Canopy closure, canopy cover and other measures. Forestry 1999, 72, 59–74. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, J.F.; Chen, G.S.; Xie, J.S.; Cai, L.P.; Lin, P. Litterfall, nutrient return, and leaf-litter decomposition in four plantations compared with a nautral forest in subtropical china. Ann. For. Sci. 2004, 61, 465–476. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Principal Components Analysis; MjM Software Design: Gleneden Beach, OR, USA, 2002; pp. 114–121. [Google Scholar]




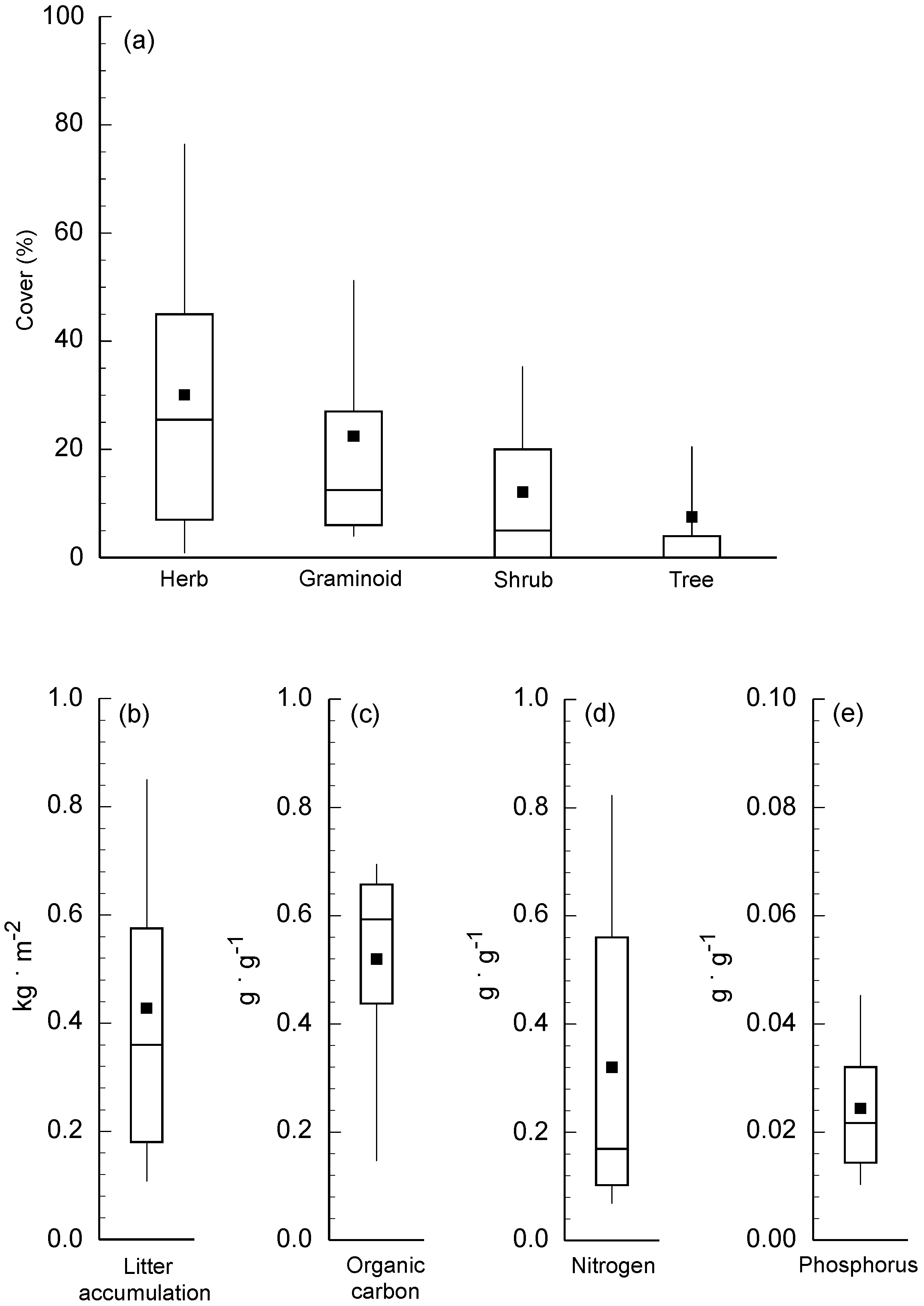{kind=link}
{kind=link}
{kind=link}
| Axis 1 | Axis 2 | Axis 3 | |
|---|---|---|---|
| Eigenvalues | 1.8 | 1.2 | 0.8 |
| Total variance (%) | 44.1 | 29.7 | 20.4 |
| Litter accumulation | 0.028 | 0.775 | −0.630 |
| Litter organic carbon | 0.938 | −0.650 | 0.041 |
| Litter total N | −0.939 | 0.012 | 0.067 |
| Litter total P | 0.065 | 0.764 | 0.642 |
| Axis 1 | Axis 3 | Frequency | Cover % | |
|---|---|---|---|---|
| Herbs | ||||
| Anemone vitifolia | 0.1207 | 0.0985 | 5 | 2.5 |
| Artemisia argyi | 0.1785 | 0.3175 | 35 | 8.5 |
| Bidens pilosa | −0.1387 | 0.0777 | 16 | 5.8 |
| Boenninghausenia albiflora | 0.0856 | 0.1851 | 3 | 24.7 |
| Cayratia japonica | 0.1271 | 0.1274 | 9 | 4.1 |
| Clematis florida | 0.0296 | 0.1864 | 3 | 2.7 |
| Commelina communis | 0.0106 | 0.1898 | 2 | 3.5 |
| Dendranthema indicum | 0.1225 | 0.0496 | 17 | 6.9 |
| Dryopteris bissetiaha | 0.0696 | 0.0040 | 10 | 6.9 |
| Epimedium brevicornum | −0.2596 | −0.0192 | 2 | 9.0 |
| Erigeron acer | 0.0280 | 0.1753 | 6 | 17.2 |
| Gelsemium elegans | 0.1109 | 0.0302 | 4 | 1.0 |
| Humulus japonicus | 0.1143 | 0.0100 | 7 | 2.7 |
| Iris japonica | −0.2758 | −0.0025 | 3 | 16.7 |
| Paederia scandens | 0.0564 | 0.1638 | 4 | 15.3 |
| Spora lygodii | −0.2032 | −0.0601 | 6 | 3.0 |
| Strobilanthes cusia | −0.0082 | −0.0142 | 6 | 2.7 |
| Taraxacum officinale | 0.1309 | 0.1963 | 4 | 2.5 |
| Torilis japonica | 0.1725 | −0.035 | 4 | 3.0 |
| Trifolium repens | 0.1435 | −0.1317 | 4 | 2.5 |
| Vicia carcca | 0.1194 | 0.2471 | 2 | 7.5 |
| Youngia japonica | 0.2022 | 0.0007 | 5 | 2.6 |
| Graminoids | ||||
| Arthraxon hispidus | −0.4535 | −0.0474 | 4 | 3.8 |
| Carex rigescens | 0.1604 | −0.0622 | 14 | 10.3 |
| Cyperus microiria | −0.1556 | −0.2874 | 16 | 9.5 |
| Cynodon dactylon | 0.0269 | 0.0664 | 2 | 15.0 |
| Cymbopogon goeringii | −0.0978 | −0.074 | 7 | 18.6 |
| Digitaria sanguinalis | 0.0281 | −0.0768 | 6 | 5.7 |
| Eragrostis ferruginea | −0.4156 | −0.0924 | 5 | 5.6 |
| Eriophorum vaginatum | 0.1036 | 0.1219 | 11 | 17.5 |
| Festuca arundinacea | 0.1253 | −0.3227 | 2 | 26.0 |
| Fimbristylis dichotoma | −0.2301 | 0.1107 | 2 | 9.0 |
| Imperata cylindrica | 0.0196 | −0.2238 | 19 | 26.5 |
| Miscanthus sinensis | 0.1812 | −0.0570 | 14 | 20.9 |
| Panicum brevifolium | 0.1014 | 0.0091 | 3 | 20.3 |
| Pogonatherum paniceum | 0.1480 | 0.0755 | 8 | 1.9 |
| Setaria viridis | 0.0709 | 0.1270 | 8 | 3.5 |
| Shrubs | ||||
| Berberis julianae | −0.2049 | −0.0133 | 2 | 8.5 |
| Boehmeria nivea | 0.0793 | −0.0142 | 3 | 7.7 |
| Broussonetia papyrifera | 0.1293 | −0.1376 | 2 | 9.5 |
| Clerodendrum bungei | 0.0958 | 0.1108 | 2 | 22.5 |
| Coriaria sinica | −0.0082 | 0.0061 | 7 | 14.7 |
| Hedera nepalensis | −0.0138 | −0.0243 | 5 | 5.6 |
| Lespedeza bicolor | −0.0929 | −0.2740 | 6 | 2.3 |
| Lindera glauca | 0.0062 | −0.1711 | 2 | 35.5 |
| Lycium chinense | 0.0783 | −0.4635 | 2 | 11.0 |
| Pueraria lobata | 0.1002 | −0.0367 | 3 | 13.3 |
| Rhus chinensis | −0.3156 | −0.1573 | 3 | 3.7 |
| Rubus corchorifolius | −0.1512 | −0.0418 | 6 | 7.2 |
| Solanum nigrum | 0.1061 | 0.2683 | 2 | 21.5 |
| Vitex negundo | −0.1467 | 0.0362 | 6 | 16.2 |
| Pyracantha fortuneana | −0.1172 | 0.0299 | 6 | 13.3 |
| Trees | ||||
| Cupressus funebris | −0.2208 | 0.0285 | 4 | 15.5 |
| Dalbergia hupeana | −0.3564 | −0.0056 | 3 | 7.7 |
| Diospyros kaki | 0.0575 | −0.1534 | 2 | 55.0 |
| Myrsine africana | −0.0512 | −0.1608 | 8 | 8.4 |
| Populus adenopoda | 0.0119 | −0.2225 | 3 | 30.7 |
| Ulmus pumila | 0.0577 | 0.1363 | 2 | 11.5 |
| Vaccinium bracteatum | −0.2469 | −0.0175 | 2 | 15.0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, H.; Monaco, T. Litter Accumulation and Nutrient Content of Roadside Plant Communities in Sichuan Basin, China. Plants 2017, 6, 36. https://doi.org/10.3390/plants6030036
He H, Monaco T. Litter Accumulation and Nutrient Content of Roadside Plant Communities in Sichuan Basin, China. Plants. 2017; 6(3):36. https://doi.org/10.3390/plants6030036
Chicago/Turabian StyleHe, Huiqin, and Thomas Monaco. 2017. "Litter Accumulation and Nutrient Content of Roadside Plant Communities in Sichuan Basin, China" Plants 6, no. 3: 36. https://doi.org/10.3390/plants6030036
APA StyleHe, H., & Monaco, T. (2017). Litter Accumulation and Nutrient Content of Roadside Plant Communities in Sichuan Basin, China. Plants, 6(3), 36. https://doi.org/10.3390/plants6030036