Flavonoid Profile of the Cotton Plant, Gossypium hirsutum: A Review



Abstract
:1. Introduction
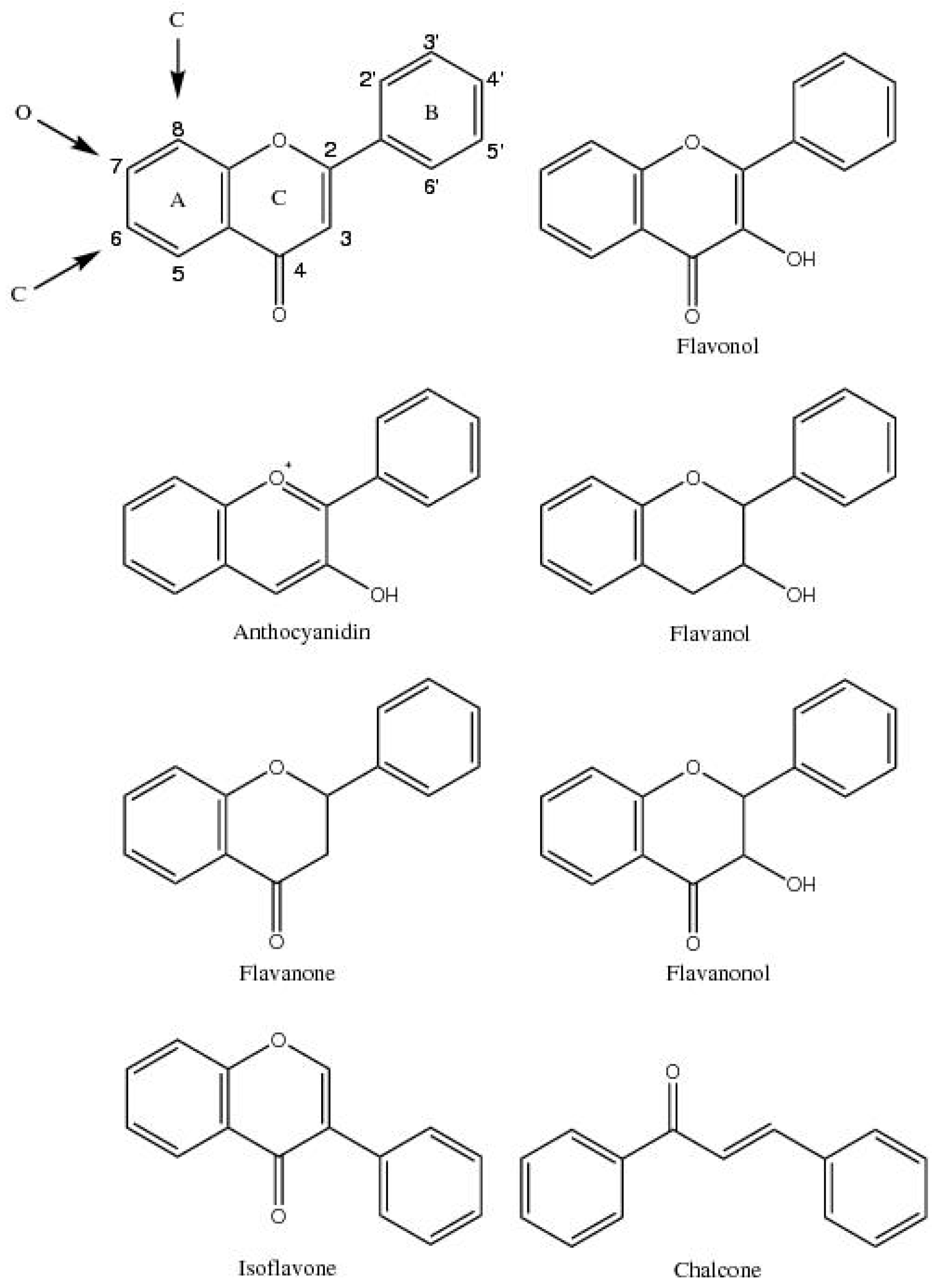
2. Review General Flavonoid Structure and Biosynthesis
2.1. Flavonoids: Defensive Role in Cotton
2.2. Leaf Reddening
2.3. Role of Flavonoids in Cotton Production
3. Flavonoids of Gossypium hirsutum and Their Distribution
4. Future Research
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Chaudhry, M.R. Cotton Production and Processing, Industrial Applications of Natural Fibres; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 219–234. [Google Scholar]
- Rani, P.U.; Pratyusha, S. Defensive role of Gossypium hirsutum L. anti-oxidative enzymes and phenolic acids in response to Spodoptera litura F. feeding. J. Asia-Pac. Entomol. 2013, 16, 131–136. [Google Scholar] [CrossRef]
- Kouakou, T.H.; Waffo-Teguo, P.; Kouadio, Y.J.; Valls, J.; Richard, T.; Decendit, A.; Merillon, J.M. Phenolic compounds and somatic embryogenesis in cotton (Gossypium hirsutum L.). Plant Cell Tissue Organ Cult. 2007, 90, 25–29. [Google Scholar] [CrossRef]
- International Cotton Advisory Committee (ICAC). Cotton: World Statistics; ICAC: Washington, DC, USA, 2014. [Google Scholar]
- Andersen, O.M.; Markham, K.R. Flavonoids Chemistry, Biochemistry and Applications; Taylor and Francis: Hoboken, NJ, USA, 2006. [Google Scholar]
- Peer, W.A.; Murphy, A.S. Flavonoids and auxin transport: modulators or regulators? Trends Plant Sci. 2007, 12, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Akashi, T.; Ayabe, S. Flavonoids of leguminous plants: Structure, biological activity, and biosynthesis. J. Plant Res. 2000, 113, 475–488. [Google Scholar] [CrossRef]
- Green, P.W.C.; Stevenson, P.C.; Simmonds, M.S.J.; Sharma, H.C. Phenolic compounds on the pod-surface of pigeonpea, Cajanus cajan, mediate feeding behavior of Helicoverpa armigera larvae. J. Chem. Ecol. 2003, 29, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Almaraz-Abarca, N.; Gonzalez-Elizondo, M.S.; Tena-Flores, J.A.; Avila-Reyes, J.A.; Herrera-Corral, J.; Naranjo-Jimenez, N. Foliar flavonoids distinguish Pinus leiophylla and Pinus chihuanuana (Coniferales: Pinaceae). Proc. Biol. Soc. Wash. 2006, 119, 426–436. [Google Scholar] [CrossRef]
- Emerenciano, V.P.; Militao, J.; Campos, C.C.; Romoff, P.; Kaplan, M.A.C.; Zambon, M.; Brant, A.J.C. Flavonoids as chemotaxonomic markers for Asteraceae. Biochem. Syst. Ecol. 2001, 29, 947–957. [Google Scholar] [CrossRef]
- Parks, C.R. Floral pigmentation studies in genus Gossypium. 1. Species specific pigmentation patterns. Am. J. Bot. 1965, 52, 309–316. [Google Scholar] [CrossRef]
- Parks, C.R. Floral pigmentation studies in genus Gossypium. 2. Chemotaxonomic analysis of diploid Gossypium species. Am. J. Bot. 1965, 52, 849–856. [Google Scholar] [CrossRef]
- Parks, C.R. Floral pigmentation studies in genus Gossypium. 3. Qualitative analysis of total flavonol content for taxonomic studies. Am. J. Bot. 1967, 54, 306–315. [Google Scholar] [CrossRef]
- Parks, C.R.; Ezell, W.L.; Williams, D.E.; Dreyer, D.L. Symposium on biochemical sytematics, genetics and origin of cultivated plants. 7. Application of flavonoid distribution to taxonomic problems in genus Gossypium. Bull. Torrey Bot. Club 1975, 102, 350–361. [Google Scholar] [CrossRef]
- Hedin, P.A.; Minyard, J.P.; Thompson, A.C. Constituents of Cotton Bud. 7. Identification of Anthocyanin as Chrysanthemin. Phytochemistry 1967, 6, 1165–1167. [Google Scholar] [CrossRef]
- Hanny, B.W. Gossypol, flavonoid, and condensed tannin content of cream and yellow anthers of five cotton (Gossypium hirsutum L.) cultivars. J. Agric. Food Chem. 1980, 28, 504–506. [Google Scholar] [CrossRef]
- Tan, J.F.; Tu, L.L.; Deng, F.L.; Hu, H.Y.; Nie, Y.C.; Zhang, X.L. A genetic and metabolic analysis revealed that cotton fiber cell development was retarded by flavonoid Naringenin. Plant Physiol. 2013, 162, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Edreva, A.; Dagnon, S.; Gurel, A.; Gesheva, E.; Hakerlerler, H. Reddening of cotton (Gossypium hirsutum L.) leaves: Analysis of the polyphenol complex. Agrochimica 2006, 50, 54–61. [Google Scholar]
- Wu, T.; Abdulla, R.; Yang, Y.; Aisa, H.A. Flavonoids from Gossypium hirsutum flowers. Chem. Nat. Compd. 2008, 44, 370–371. [Google Scholar] [CrossRef]
- Feng, H.J.; Tian, X.H.; Liu, Y.C.; Li, Y.J.; Zhang, X.Y.; Jones, B.J.; Sun, Y.Q.; Sun, J. Analysis of flavonoids and the flavonoid structural genes in brown fiber of Upland Cotton. PLoS ONE 2013, 8, e58820. [Google Scholar] [CrossRef] [PubMed]
- Ismailov, A.I.; Karimdzhanov, A.K.; Islambekov, S.Y.; Rakhimkhanov, Z.B. Flavonoids of the cotton plant and plants close to it. Chem. Nat. Compd. 1994, 30, 1–14. [Google Scholar] [CrossRef]
- Corradini, E.; Foglia, P.; Giansanti, P.; Gubbiotti, R.; Samperi, R.; Lagana, A. Flavonoids: Chemical properties and analytical methodologies of identification and quantitation in foods and plants. Nat. Prod. Res. 2011, 25, 469–495. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.E. The Flavonoids: Advances in Research Since 1986; Chapman & Hall: London, UK, 1994. [Google Scholar]
- Brunetti, C.; Di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants and developmental regulators: Relative significance in plants and humans. Int. J. Mol. Sci. 2013, 14, 3540–3555. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.P.; Grotewold, E. Flavonoids as developmental regulators. Curr. Opin. Plant Biol. 2005, 8, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.M.; Schwinn, K.E. Molecular biology and biotechnology of flavonoid biosynthesis. In Flavonoids Chemistry, Biochemistry and Applications; Andersen, O.M., Markham, K.R., Eds.; Taylor and Francis: Hoboken, NJ, USA, 2006; pp. 143–218. [Google Scholar]
- Brown, D.E.; Rashotte, A.M.; Murphy, A.S.; Normanly, J.; Tague, B.W.; Peer, W.A.; Taiz, L.; Muday, G.K. Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis. Plant Physiol. 2001, 126, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.; Peer, W.A.; Taiz, L. Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta 2000, 211, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Peer, W.A.; Bandyopadhyay, A.; Blakeslee, J.J.; Makam, S.I.; Chen, R.J.; Masson, P.H.; Murphy, A.S. Variation in expression and protein localization of the PIN family of auxin efflux facilitator proteins in flavonoid mutants with altered auxin transport in Arabidopsis thaliana. Plant Cell 2004, 16, 1898–1911. [Google Scholar] [CrossRef] [PubMed]
- Edwards, W.R.; Hall, J.A.; Rowlan, A.R.; Schneider-Barfield, T.; Sun, T.J.; Patil, M.A.; Pierce, M.L.; Fulcher, R.G.; Bell, A.A.; Essenberg, M. Light filtering by epidermal flavonoids during the resistant response of cotton to Xanthomonas protects leaf tissue from light-dependent phytoalexin toxicity. Phytochemistry 2008, 69, 2320–2328. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.G.; Waiss, A.C.; Binder, R.G.; Elliger, C.A. Inhibition of Lepidopterous larval growth by cotton constituents. Entomol. Exp. Appl. 1978, 24, 294–300. [Google Scholar] [CrossRef]
- Chan, B.G.; Waiss, A.C.; Lukefahr, M. Condensed tannin, an antibiotic chemical from Gossypium hirsutum. J. Insect Physiol. 1978, 24, 113–118. [Google Scholar] [CrossRef]
- Shaver, T.N.; Lukefahr, M.J. Effect of flavonoid pigments and Gossypol on growth and development of bollworm, tobacco budworm and pink bollworm. J. Econ. Entomol. 1969, 62, 643–646. [Google Scholar] [CrossRef]
- Harborne, J.B. Twenty-five years of chemical ecology. Nat. Prod. Rep. 2001, 18, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Hedin, P.A.; Jenkins, J.N.; Collum, D.H.; White, W.H.; Parrott, W.L. Multiple Factors in Cotton Contributing to Resistance to the Tobacco Budworm, Heliothis-Virescens F. Acs Symp. Ser. 1983, 208, 347–365. [Google Scholar] [CrossRef]
- Hedin, P.A.; Parrott, W.L.; Jenkins, J.N.; Mulrooney, J.E.; Menn, J.J. Elucidating mechanisms of tobacco budworm resistance to allelochemicals by dietary tests with insecticide synergists. Pestic. Biochem. Physiol. 1988, 32, 55–61. [Google Scholar] [CrossRef]
- Hedin, P.A.; Jenkins, J.N.; Parrott, W.L. Evaluation of flavonoids in Gossypium arboreum (L.) cottons as potential source of resistance to tobacco budworm. J. Chem. Ecol. 1992, 18, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.E. Inactivation of pectic enzymes by polyphenols in cotton seedlings of different ages infected with Rhizoctonia solani. Physiol. Plant Pathol. 1974, 4, 151–159. [Google Scholar] [CrossRef]
- Mace, M.E.; Bell, A.A.; Stipanovic, R.D. Histochemistry and identification of flavanols in Verticillium wilt-resistant and -susceptible cottons. Physiol. Plant Pathol. 1978, 13, 143–146. [Google Scholar] [CrossRef]
- Howell, C.R.; Bell, A.A.; Stipanovic, R.D. Effect of aging on flavonoid content and resistance of cotton leaves to verticillium wilt. Physiol. Plant Pathol. 1976, 8, 181–188. [Google Scholar] [CrossRef]
- Edreva, A.; Gurel, A.; Gesheva, E.; Hakerlerler, H. Reddening of cotton (Gossypium hirsutum L.) leaves. Biol. Plant 2002, 45, 303–306. [Google Scholar] [CrossRef]
- Pagare, G.A.; Durge, D.V. Pigment analysis with reference to leaf reddening in Bt cotton. Ann. Plant Physiol. 2010, 24, 102–103. [Google Scholar]
- Aaby, K.; Hvattum, E.; Skrede, G. Analysis of flavonoids and other phenolic compounds using high-performance liquid chromatography with coulometric array detection: Relationship to antioxidant activity. J. Agric. Food Chem. 2004, 52, 4595–4603. [Google Scholar] [CrossRef] [PubMed]
- Kangatharalingam, N.; Pierce, M.L.; Bayles, M.B.; Essenberg, M. Epidermal anthocyanin production as an indicator of bacterial blight resistance in cotton. Physiol. Mol. Plant Pathol. 2002, 61, 189–195. [Google Scholar] [CrossRef]
- Manetas, Y. Why some leaves are anthocyanic and why most anthocyanic leaves are red? Flora 2006, 201, 163–177. [Google Scholar] [CrossRef]
- Pratt, C.; Wender, S.H. Identification of Kaempferol-3-rhamnoglucoside and Quercetin-3-glucoglucoside in cottonseed. J. Am. Oil Chem. Soc. 1961, 38, 403–405. [Google Scholar] [CrossRef]
- Struck, R.F.; Kirk, M.C. Methylated flavonols in Genus Gossypium. J. Agric. Food Chem. 1970, 18, 548–549. [Google Scholar] [CrossRef]
- Pakudina, Z.P.; Sadykov, A.S. Isoastragalin—A flavonol glycoside from the flowers of Gossypium hirsutum. Chem. Nat. Compd. 1970, 6, 24–25. [Google Scholar] [CrossRef]
- Blouin, F.A.; Zarins, Z.M. High-perfomance liquid chromatography of cottonseed flavonoids. J. Chromatogr. 1988, 441, 443–447. [Google Scholar] [CrossRef]
- Makhsudova, B.; Pakudina, Z.P.; Sadykov, A.S. The structure of hybridin. Chem. Nat. Compd. 1969, 5, 273. [Google Scholar] [CrossRef]
- Pratt, C.; Wender, S. Identification of rutin and isoquercitrin in cottonseed. J. Am. Oil Chem. Soc. 1959, 36, 392–394. [Google Scholar] [CrossRef]
- Pakudina, Z.P.; Leont’ev, V.B.; Kamaev, F.G.; Sadykov, A.S. Structure and PMR spectra of isoquercitrin and hirsutrin. Chem. Nat. Compd. 1970, 6, 572–574. [Google Scholar] [CrossRef]
- Pakudina, Z.P.; Sadykov, A.S.; Denliev, P.K. The flavonols of the flowers of Gossypium hirsutum L. (cotton variety 108-F). Chem. Nat. Compd. 1965, 1, 52–54. [Google Scholar] [CrossRef]
- Kouakou, T.H.; Kouadio, Y.J.; Teguo, P.W.; Valls, J.; Badoc, A.; Merillon, J.M.; Decendit, A. Polyphenol levels in two cotton (Gossypium hirsutum L.) callus cultures. Acta Bot. Gall. 2009, 156, 223–231. [Google Scholar] [CrossRef]
- Elliger, C.A. Sexangularetin 3-glucoside-7-rhamnoside from Gossypium hirsutum. Phytochemistry 1984, 23, 1199–1201. [Google Scholar] [CrossRef]
- Ghosh, D.; Joham, H.E. Leaf anthocyanin content of Gossypium hirsutum as influenced by magnesium and nitrogen deficiencies. Plant Physiol. 1964, 39, 15501. [Google Scholar]
- Karimdzhanov, A.K.; Kuznetsova, N.N.; Dzhataev, S.A. Phenolic compounds of the plant Gossypium hirsutum and of callus tissue from its anthers. Chem. Nat. Compd. 1997, 33, 187–189. [Google Scholar] [CrossRef]
- Rakhimkhanov, Z.B.; Karimdzhanov, A.K.; Ismailov, A.I.; Sadykov, A.S. A study of the anthocyanins of the cotton plant. Chem. Nat. Compd. 1968, 4, 190–191. [Google Scholar] [CrossRef]
- Zhang, H.L.; Nagatsu, A.; Okuyama, H.; Mizukami, H.; Sakakibara, J. Sesquiterpene glycosides from cotton oil cake. Phytochemistry 1998, 48, 665–668. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, Y.; Wu, K.; Wang, W. HPLC analysis of main flavonoid chemicals and their spatio-temporal dynamics in Bt transgenic cotton. Chin. J. Appl. Ecol. 2003, 14, 246–248. [Google Scholar]
- Baker, G.H.; Tann, C.R.; Fitt, G.P. Production of Helicoverpa spp. (Lepidoptera, Noctuidae) from different refuge crops to accompany transgenic cotton plantings in eastern Australia. Aust. J. Agric. Res. 2008, 59, 723–732. [Google Scholar] [CrossRef]
- Akhurst, R.J.; James, W.; Bird, L.J.; Beard, C. Resistance to the Cry1Ac delta-endotoxin of Bacillus thuringiensis in the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2003, 96, 1290–1299. [Google Scholar] [CrossRef] [PubMed]
- Mahon, R.J.; Olsen, K.M.; Garsia, K.A.; Young, S.R. Resistance to Bacillus thuringiensis toxin Cry2Ab in a strain of Helicoverpa armigera (Lepidoptera: Noctuidae) in Australia. J. Econ. Entomol. 2007, 100, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Elliger, C.A.; Chan, B.C.; Waiss, A.C. Flavonoids as larval growth-Inhibitors-Structural factors governing toxicity. Naturwissenschaften 1980, 67, 358–360. [Google Scholar] [CrossRef]
- Nishida, R.; Ohsugi, T.; Fukami, H.; Nakajima, S. Oviposition deterrent of a Rutaceae-feeding swallowtail butterfly, Papilio xuthus, from a non-host rutaceous plant. Orixa japonica. Agric. Biol. Chem. 1990, 54, 1265–1270. [Google Scholar]
- Baker, G.H.; Tann, C.R. Mating of Helicoverpa armigera (Lepidoptera: Noctuidae) moths and their host plant origins as larvae within Australian cotton farming systems. Bull. Entomol. Res. 2013, 103, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Brevault, T.; Nibouche, S.; Achaleke, J.; Carriere, Y. Assessing the role of non-cotton refuges in delaying Helicoverpa armigera resistance to Bt cotton in West Africa. Evol. Appl. 2012, 5, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orth, R.G.; Head, G.; Mierkowski, M. Determining larval host plant use by a polyphagous lepidopteran through analysis of adult moths for plant secondary metabolites. J. Chem. Ecol. 2007, 33, 1131–1148. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, F.; Proksch, P.; Fiedler, K. Flavonoid sequestration by the common blue butterfly Polyommatus icarus: Quantitative intraspecific variation in relation to larval hostplant, sex and body size. Biochem. Syst. Ecol. 2001, 29, 875–889. [Google Scholar] [CrossRef]
- Ford, E.B. Studies on the chemistry of pigments in the Lepidoptera, with reference to their bearing on systematics. I. The anthoxanthins. Proc. R. Entomol. Soc. Lond. Ser. A 1941, 16, 65–90. [Google Scholar] [CrossRef]
- Wiesen, B.; Krug, E.; Fiedler, K.; Wray, V.; Proksch, P. Sequestration of host-plant-derived flavonoids by lycaenid butterfly Polyommatus icarus. J. Chem. Ecol. 1994, 20, 2523–2538. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A. Flavonoid pigments in marbled white butterfly (Melanargia galathea) are dependent on flavonoid content of larval diet. J. Chem. Ecol. 1985, 11, 1161–1179. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A. Flavonoid pigments of butterflies in the genus Melanargia. Phytochemistry 1985, 24, 1685–1691. [Google Scholar] [CrossRef]
- Wilson, A. Flavonoid Pigments in Swallowtail Butterflies. Phytochemistry 1986, 25, 1309–1313. [Google Scholar] [CrossRef]
- Wilson, A. Flavonoid pigments in chalkhill blue (Lysandra coridon Poda) and other lycaenid butterflies. J. Chem. Ecol. 1987, 13, 473–493. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Flavonoid Class | Plant Organ | Fibre | Total | |||||
|---|---|---|---|---|---|---|---|---|
| Leaves | Roots/Root Bark | Stem/Stem Bark | Seed | Boll | Flower | |||
| Isoflavones | 1 | 1 | ||||||
| Flavonols | 11 | 2 | 1 | 10 | 2 | 29 | 2 | 36 |
| Flavanones | 2 | 2 | 1 | 2 | 2 | 2 | ||
| Anthocyanidins | 3 | 2 | 2 | 3 | 5 | |||
| Flavanols | 1 | 4 | 4 | 1 | 4 | 4 | 4 | |
| Flavanonol | 2 | 2 | 2 | 2 | 2 | |||
| Leucoanthocyanidins | 2 | 2 | 2 | 2 | 2 | |||
| Total | 19 | 12 | 11 | 11 | 10 | 42 | 6 | 52 |
| Name | Structure | Organ Isolated | Reference |
|---|---|---|---|
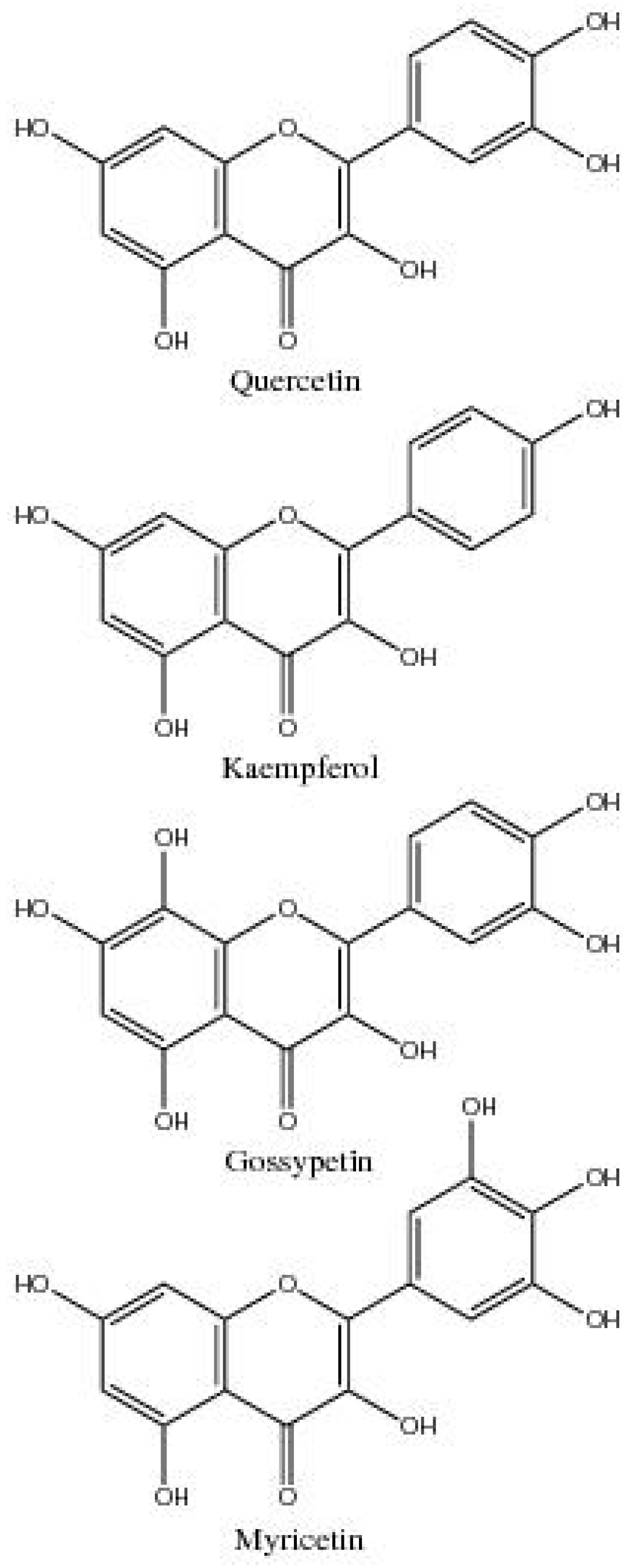
| Gossypetin | 3,5,7,8,3′,4′-Hexahydroxyflavone | Flower petals | [14] |
| Gossypetin 3′,7-diglucosidoglucoside | Anthers | [16] | |
| Gossypetin 3-glucoside | Anthers | [16] | |
| Gossypetin glycoside (C7-linked unknown sugar) | Flower petals | [11] | |
| Gossypin | Gossypetin 8-glucoside | Flower petals, anthers | [11,14,16] |
| Gossypitrin | Gossypetin 7-glucoside | Flower petals, anthers | [11,14,16] |
| Kaempferol | 3,5,7,4′-Tetrahydroxyflavone | Ovules, Fibre, Roots, Cotyledons, Leaves, Flower buds/petals, Bolls, Seed | [14,17,20,46,47] |
| Astragalin | Kaempferol 3-O-β-d-glucopyranoside | Flowers | [14,19] |
| Isoastragalin | Kaempferol 3-α-d-glucofuranoside | Flowers | [48] |
| Kaempferide | 4′-O-Methylkaempferol | Flower petals | [47] |
| Kaempferol C3-linked glycoside (unknown sugar) | Flower petals | [11] | |
| Kaempferol 3-diglucoside | Seed | [49] | |
| Kaempferol-3-O-b-d-(6″-O-p-coumaroyl)-glycoside | Flowers | [19] | |
| Kaempferol-3-O-neohesperidoside | Seed | [49] | |
| Nicotiflorin | Kaempferol 3-rutinoside | Flower petals, leaves, and seed | [11,14,18,46] |
| Tiliroside | Kaempferol 3-O-β-d-(6″-O-(E)-p-coumaroyl) glucopyranoside | Flowers | [19] |
| Trifolin | Kaempferol-3-O-galactoside | Flower petals | [11,13] |
| Myricetin | 3,5,7,3′,4′,5′-Hexahydroxyflavone | Ovules and fibres from flower buds and bolls | [20] |
| Quercetin | 3,5,7,3’,4’-Pentahydroxyflavone | Flowers/petals, leaves, ovules, fibre, roots, and cotyledons | [14,17,18,19,20,47] |
| Hirsutrin | Quercetin 3-O-β-d-glucopyranoside | Flower and Leaves | [48] |
| Hybridin | Quercetin 3-O-[O-β-d-galactofuranosyl-(l→3)-O-β-d-glucopyranosyl-(l→3)-xylopyranoside] | Leaf | [50] |
| Hyperoside | Quercetin-3-galactoside | Flowers | [19] |
| Isoquercitrin | Quercetin 3-β-d-glucoside | Flowers, leaves, cotyledons, and seed | [11,13,14,16,18,19,30,49,51,52] |
| Quercetin 3′-glucoside | Flowers, anthers | [16,19,53] | |
| Quercetin 3-diglucoside | Anthers and seed | [16,46,49] | |
| Quercetin 7-rhamnoglucoside | Anthers | [16] | |
| Quercetin C7-linked glycoside (unknown sugar) | Flower petals | [11] | |
| Quercetin-3-O-neohesperidoside | Seed | [49] | |
| Quercetin-3-O-robinoside | Seed | [49] | |
| Quercimeritrin | Quercetin 7-glucoside | Flowers/petals | [11,13,14,15,53] |
| Quercitrin | Quercetin 3-rhamnoside | Leaves | [18] |
| Rutin | Quercetin 3-rutinoside | Flower petals, anthers, leaves, hypocotyls, seed, and calli | [4,11,13,14,16,18,49,51,54] |
| Sexangularetin 3-glucoside-7-rhamnoside | Immature flower buds | [55] | |
| Spiraeoside | Quercetin 4′-O-glucoside | Flower petals, and seed | [14,55] |
| Tamarixetin | Quercetin 4′-methyl ether | Flower petals | [47] |
| Tamarixetin 7-glucoside | Quercetin-4′-O-methyl-7-glucoside | Flower petals | [14] |
| Name | Structure | Organ Isolated from | Reference |
|---|---|---|---|
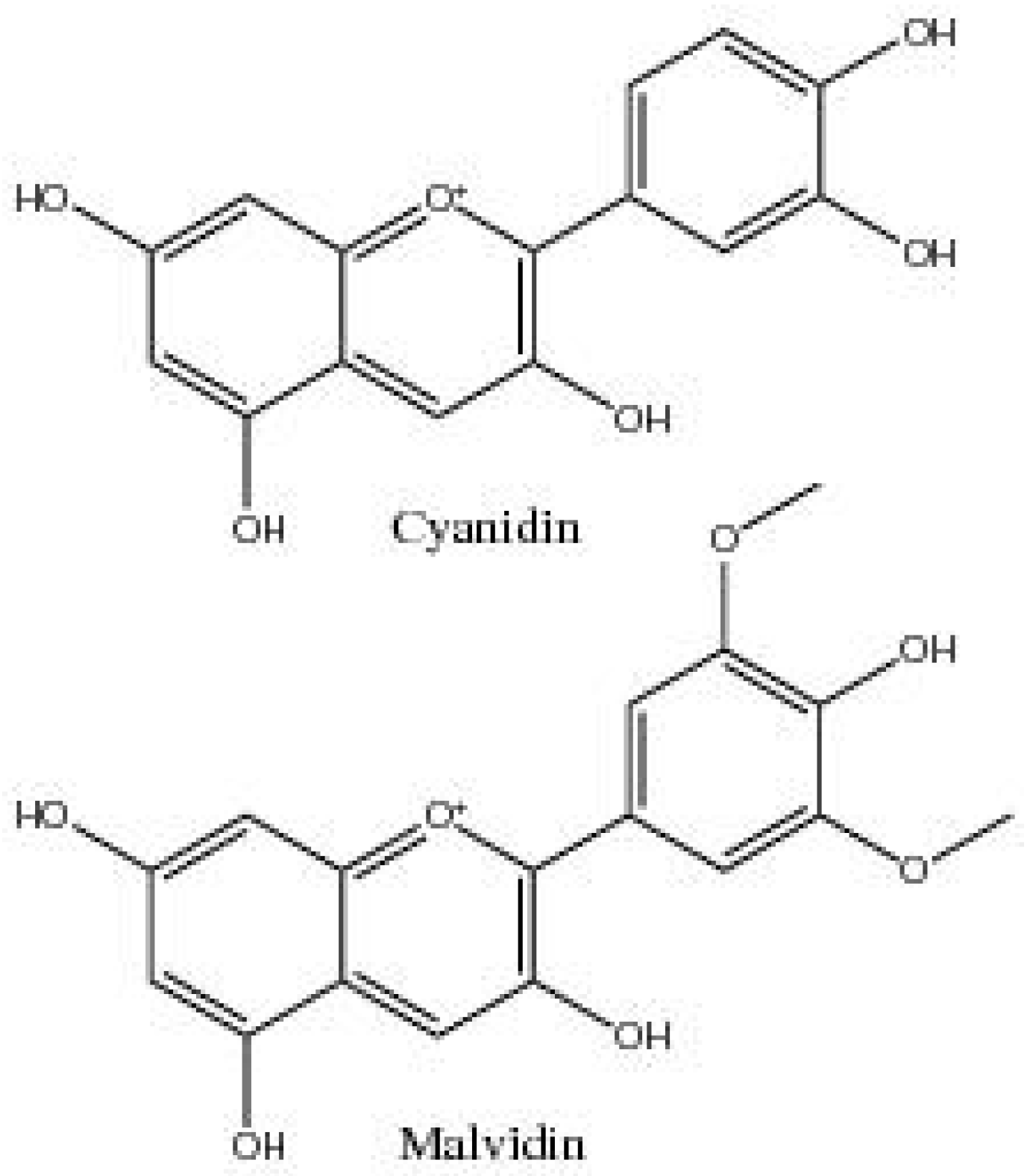
| Cyanidin | 3,5,7,3′,4′-Pentahydroxyflavylium | Flower petals and leaves | [11,18,56] |
| Chrysanthemin | Cyanidin 3-glucoside | Flower/buds, leaves, cotyledons, anther tissue culture, boll valves, and stem bark | [15,16,30,57,58] |
| Gossypicyanin | Cyanidin 3-O-[O-β-d-xylopyranosyl-(1→4)-β-D-glucopyranoside | Anther tissue culture, boll valves, stem bark, flowers | [57] |
| Ilicicyanin | Cyanidin 3-xylosylglucoside | [21] | |
| Malvidin | 3′,5′-Dimethoxy-3,4′,5,7-tetrahydroxyflavylium | Leaves | [18] |
| Class | Name | Structure | Organ Isolated from | Reference |
|---|---|---|---|---|
| Isoflavone | ||||
| Genistein | 5,7,4′-trihydroxyisoflavone | Hypocotyls | [3] | |
| Flavanones | ||||
| Eriodictyol | (2S)-2-(3,4-Dihydroxyphenyl)-5,7-dihydroxy-4-chromanone | Ovules, fibre, roots, cotyledons, and leaves | [17] | |
| Naringenin | 5,7-dihydroxy-2-(4-hydroxyphenyl)chroman-4-one | Hypocotyls, ovules, fibre, roots, cotyledons & leaves | [3,17,20] | |
| Flavanols | ||||
| Catechin | (2R,3S)-2-(3,4-dihydroxyphenyl)-3,4-dihydro-2H-chromene-3,5,7-triol | Leaves, hypocotyls, stem, calli, anther, boll valves, stem bark, root bark, and cotton oil cake | [2,3,39,54,57,59] | |
| (−)-Epicatechin | (−)-cis-3,3′,4′,5,7-Pentahydroxyflavane | Anther, boll valves, stem bark and root bark | [57] | |
| (−)-Epigallocatechin | (−)-cis-3,3′,4′,5,5′,7-Hexahydroxyflavane | Anther, boll valves, stem bark, root bark | [57] | |
| Gallocatechin | (2S,3R)-2-(3,4,5-Trihydroxyphenyl)-3,4-dihydro-1(2H)-benzopyran-3,5,7-triol | Hypocotyls/stem steles, anther, boll valves, stem bark, root bark | [39,57] | |
| Flavanonol | ||||
| Aromadendrin | (2R,3R)-3,5,7-trihydroxy-2-(4-hydroxyphenyl)-2,3-dihydrochromen-4-one | Ovules, fibre, roots, cotyledons, and leaves | [17] | |
| Taxifolin | (2R,3R)-2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxy-2, 3-dihydrochromen-4-one | Ovules, fibre, roots, cotyledons, and leaves | [17] | |
| Leucoanthocyanidins | ||||
| Leucocyanidin | (2R,3S,4S)-2-(3,4-dihydroxyphenyl)-3,4-dihydro-2H-chromene-3,4,5,7-tetrol | Anther, boll valves, stem bark, root bark | [57] | |
| Leucodelphinidin | (2R,3S,4S)-2-(3,4,5-trihydroxyphenyl)-3,4-dihydro-2H-chromene-3,4,5,7-tetrol | Anther, boll valves, stem bark, root bark | [57] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nix, A.; Paull, C.; Colgrave, M. Flavonoid Profile of the Cotton Plant, Gossypium hirsutum: A Review. Plants 2017, 6, 43. https://doi.org/10.3390/plants6040043
Nix A, Paull C, Colgrave M. Flavonoid Profile of the Cotton Plant, Gossypium hirsutum: A Review. Plants. 2017; 6(4):43. https://doi.org/10.3390/plants6040043
Chicago/Turabian StyleNix, Aaron, Cate Paull, and Michelle Colgrave. 2017. "Flavonoid Profile of the Cotton Plant, Gossypium hirsutum: A Review" Plants 6, no. 4: 43. https://doi.org/10.3390/plants6040043
APA StyleNix, A., Paull, C., & Colgrave, M. (2017). Flavonoid Profile of the Cotton Plant, Gossypium hirsutum: A Review. Plants, 6(4), 43. https://doi.org/10.3390/plants6040043