Functions of Flavonoids in Plant–Nematode Interactions


Abstract
:1. Introduction to Plant Parasitic Nematodes
2. Plant–Nematode Interactions
3. Flavonoids in Plants
4. Flavonoids as Defense Compounds against Nematodes
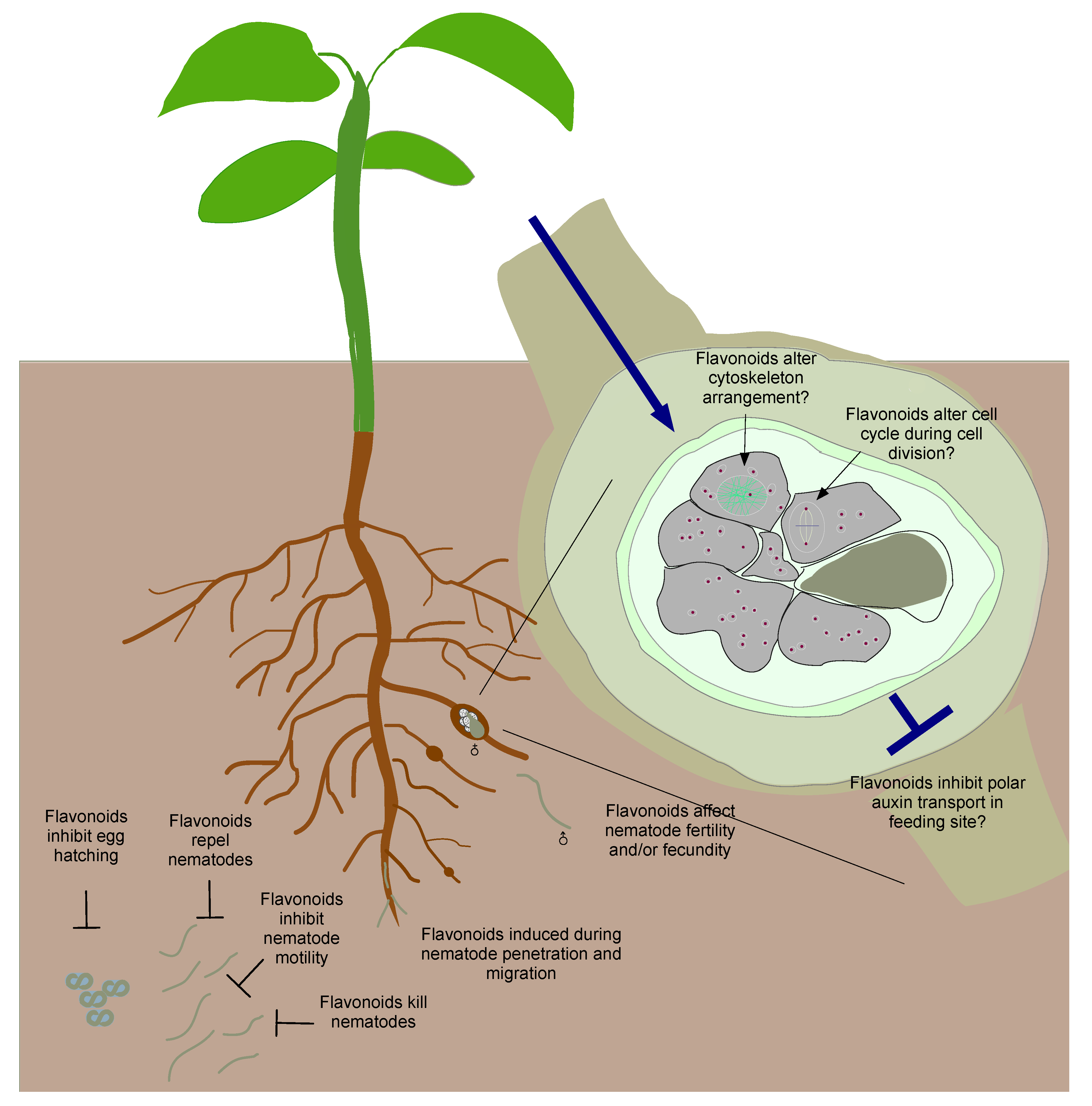
5. Flavonoids Play Minor Roles in the Development of Nematode Feeding Sites
6. Summary and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abad, P.; Williamson, V.M. Plant nematode interaction: A sophisticated dialogue. Adv. Botanical Res. 2010, 53, 147–192. [Google Scholar]
- Bird, A.F. Moulting of parasitic nematodes. Int. J. Parasitol. 1987, 17, 233–239. [Google Scholar] [CrossRef]
- Decraemer, W.; Hunt, D.J. Structure and classification. In Plant Nematology; Perry, R.N., Moens, M., Eds.; CAB International: Wallingford, UK, 2006; pp. 3–32. [Google Scholar]
- Nicol, J.M.; Turner, S.J.; Coyne, D.L.; Nijs, L.D.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Springer: Dordrecht, The Netherlands, 2011; pp. 21–44. [Google Scholar]
- Sasser, J.N.; Freckman, D.W. A World Perspective on Nematology: The Role of the Society. In Vistae on Nematology; Veech, J.A., Dickson, D.W., Eds.; Society of Nematologists Inc.: Hyattsville, MD, USA, 1987; pp. 7–14. [Google Scholar]
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture–Agricultural Research Service. Pest Manag. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef] [PubMed]
- McCarter, J.P. Molecular approaches toward resistance to plant-parasitic nematodes. In Cell Biology of Plant Nematode Parasitism; Berg, R.H., Taylor, C.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 239–268. [Google Scholar]
- Perry, R.N.; Moens, M. Introduction to plant-parasitic nematodes; Modes of parasitism. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 3–20. [Google Scholar]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.; Gaur, H.S.; Helder, J.; Jones, M.G.; Perry, R.N. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tytgat, T.; De Meutter, J.; Gheysen, G.; Coomans, A. Sedentary endoparasitic nematodes as a model for other plant parasitic nematodes. Nematology 2000, 2, 113–121. [Google Scholar] [CrossRef]
- Grandison, G.S. Relationships of plant-parasitic nematodes and their hosts. New Z. Entomol. 1977, 6, 262–266. [Google Scholar] [CrossRef]
- Quitst, C.W.; Smant, G.; Helder, J. Evolution of plant parasitism in the phylum Nematoda. Annu. Rev. Phytopathol. 2015, 53, 289–310. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Eves-van den Akker, S.; Jones, J.T. Genome evolution of plant-parasitic nematodes. Annu. Rev. Phytopathol. 2017, 55, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.N. Chemoreception in plant parasitic nematodes. Annu. Rev. Phytopathol. 1996, 34, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Curtis, R.H.C. Plant-nematode interactions: Environmental signals detected by the nematode’s chemosensory organs control changes in the surface cuticle and behaviour. Parasite 2008, 15, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Gang, S.S.; Hallem, E.A. Mechanisms of host seeking by parasitic nematodes. Mol. Biochem. Parasitol. 2016, 208, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengarajan, S.; Hallem, E.A. Olfactory circuits and behaviors of nematodes. Curr. Opin. Neurobiol. 2016, 41, 136–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tefft, P.M.; Bone, L.W. Plant-induced hatching of eggs of the soybean cyst nematode, Heterodera glycines. J. Nematol. 1985, 17, 275–279. [Google Scholar] [PubMed]
- Rasmann, S.; Ali, J.G.; Helder, J.; van der Putten, W.H. Ecology and evolution of soil nematode chemotaxis. J. Chem. Ecol. 2012, 38, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Pline, M.; Dusenbery, D.B. Responses of plant-parasitic nematode Meloidogyne incognita to carbon dioxide determined by video camera-computer tracking. J. Chem. Ecol. 1986, 13, 873–888. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Bruening, G.; Williamson, V.M. Determination of preferred pH for root-knot nematode aggregration using Pluronic F-127 gel. J. Chem. Ecol. 2009, 35, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Sommerville, R.I.; Davey, K.G. Diapause in parasitic nematodes: A review. Can. J. Zool. 2002, 80, 1817–1840. [Google Scholar] [CrossRef]
- Evans, A.A.F.; Perry, R.N. Survival mechanisms. In Root-knot Nematodes; Perry, R.N., Moens, M., Starr, J.L., Eds.; CAB International: Wallingford, UK, 2009; pp. 201–222. [Google Scholar]
- Lambert, K.N.; Bekal, S. Introduction to plant parasitic nematodes. Plant Health Instr. 2002, 10, 1094–1218. [Google Scholar] [CrossRef]
- Wyss, U.; Grundler, F.M.W. Feeding behavior of sedentary plant parasitic nematodes. Eur. J. Plant Pathol. 1992, 98, 165–173. [Google Scholar] [CrossRef]
- Sijmons, P.C.; Atkinson, H.J.; Wyss, U. Parasitic strategies of root nematodes and associated host cell responses. Annu. Rev. Phytopathol. 1994, 32, 235–259. [Google Scholar] [CrossRef]
- Bird, D.M.; Kaloshian, I. Are roots special? Nematodes have their say. Physiol. Mol. Plant Pathol. 2003, 62, 115–123. [Google Scholar] [CrossRef]
- Fuller, V.L.; Lilley, C.J.; Urwin, P.E. Nematode Resistance. New Phytol. 2008, 180, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Stirling, G.R.; Pattison, B. Beyond chemical dependency for managing plant-parasitic nematodes: Examples from the banana, pineapple and vegetable industries of tropical and subtropical Australia. Aust. Plant Pathol. 2008, 37, 254–267. [Google Scholar] [CrossRef]
- Davies, K.G.; Spiegel, Y. Biological control of plant-parasitic nematodes: Towards understanding field variation through molecular mechanisms. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J.T., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 493–516. [Google Scholar]
- Williamson, V.M.; Kumar, A. Nematode resistance in plants: The battle underground. Trends Genet. 2006, 22, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant flavonoids-biosynthesis, transport and involvement in stress responses. Int. J. Mol. Sci. 2013, 14, 14950–14973. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid biosynthesis: A colourful model for genetics, biochemistry, cell biology and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Hassan, S.; Mathesius, U. The role of flavonoids in root-rhizosphere signalling: Opportunities and challenges for improving plant-microbe interactions. J. Exper. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef] [PubMed]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, A.; Shitan, N.; Yazaki, K. Involvement of a soybean ATP-binding cassette-type transporter in the secretion of genistein, a signal flavonoid in legume-rhizobium symbiosis. Plant Physiol. 2007, 144, 2000–2008. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Loyola-Vargas, V.M.; Broeckling, C.D.; De-la-Peña, C.; Jasinski, M.; Santelia, D.; Vivanco, J.M. Altered profile of secondary metabolites in the root exudates of Arabidopsis ATP-binding cassette transporter mutants. Plant Physiol. 2008, 146, 762–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesco, S.; Neumann, G.; Tomasi, N.; Pinton, R.; Weisskopf, L. Release of plant-borne flavonoids into the rhizosphere and their role in plant nutrition. Plant Soil 2010, 329, 1–25. [Google Scholar] [CrossRef]
- Hawes, M.C.; Brigham, L.A.; Wen, F.; Woo, H.H.; Zhu, Y. Function of root border cells in plant health: Pioneers in the rhizosphere. Annu. Rev. Phytopathol. 2008, 36, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Wuyts, N.; Swennen, R.; De Waele, D. Effects of plant phenylpropanoid pathway products and selected terpenoids and alkaloids on the behaviour of the plant-parasitic nematodes Radopholus similis, Pratylenchs penetrans and Meloidogyne incognita. Nematology 2006, 8, 89–101. [Google Scholar] [CrossRef]
- Faizi, S.; Fayyaz, S.; Bano, S.; Yawar Iqbal, E.; Lubna; Siddiqi, H.; Naz, A. Isolation of nematicidal compounds from Tagetes patula L. yellow flowers: Structure–activity relationship studies against cyst nematode Heterodera zeae infective stage larvae. J. Agric. Food Chem. 2011, 59, 9080–9093. [Google Scholar] [CrossRef] [PubMed]
- Kampkötter, A.; Gombitang Nkwonkam, C.; Zurawski, R.F.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. Effects of the flavonoids kaempferol and fisetin on thermotolerance, oxidative stress and FoxO transcription factor DAF-16 in the model organism Caenorhabditis elegans. Arch. Toxicol. 2007, 81, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Kampkötter, A.; Timpel, C.; Zurawski, R.F.; Ruhl, S.; Chovolou, Y.; Proksch, P.; Wätjen, W. Increase of stress resistance and lifespan of Caenorhabditis elegans by quercetin. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 149, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Grünz, G.; Haas, K.; Soukup, S.; Klingenspor, M.; Kulling, S.E.; Daniel, H.; Spanier, B. Structural features and bioavailability of four flavonoids and their implications for lifespan-extending and antioxidant actions in C. elegans. Mech. Ageing Dev. 2012, 133, 1–10. [Google Scholar] [CrossRef] [PubMed]
- González, J.A.; Estévez-Braun, A. Effect of (E)-Chalcone on Potato-Cyst Nematodes (Globodera pallida and G. rostochiensis). J. Agric. Food Chem. 1998, 46, 1163–1165. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.; Tiller, S.A.; Mizen, K.A.; Edwards, R. Isoflavonoid metabolism in resistant and susceptible cultivars of white clover infected with the stem nematode Ditylenchus dipsaci. J. Plant Physiol. 1995, 146, 348–354. [Google Scholar] [CrossRef]
- Vlachopoulos, E.G.; Smith, L. Flavonoids in potato cyst nematodes. Fundam. Appl. Nematol. 1993, 16, 103–106. [Google Scholar]
- Goverse, A.; Smant, G. The activation and suppression of plant innate immunity by parasitic nematodes. Annu. Rev. Phytopathol. 2014, 52, 243–265. [Google Scholar] [CrossRef] [PubMed]
- Holbein, J.; Grundler, F.M.W.; Siddique, S. Plant basal resistance to nematodes: An update. J. Exp. Bot. 2016, 67, 2049–2061. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dixon, R.A. The ‘ins’ and ‘outs’ of flavonoid transport. Trends Plant Sci. 2009, 15, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.; Mizen, T.; Cook, R. Isoflavonoid conjugate accumulation in the roots of lucerne (Medicago sativa) seedlings following infection by the stem nematode (Ditylenchus dipsaci). Nematologica 1995, 51–66. [Google Scholar] [CrossRef]
- Dewick, P.M. The biosynthesis of shikimate metabolites. Nat. Prod. Rep. 1998, 15, 17–58. [Google Scholar] [CrossRef] [PubMed]
- Lambert, K.N.; Allen, K.D.; Sussex, I.M. Cloning and characterization of an esophageal-gland specific chorismate mutase from the phytoparasitic nematode Meloidogyne javanica. Mol. Plant Microbe Interact. 1999, 12, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.T.; Furlanetto, C.; Bakker, E.; Banks, B.; Blok, V.; Chen, Q.; Phillips, M.; Prior, A. Characterization of a chorismate mutase from the potato cyst nematode Globedera pallida. Mol. Plant Pathol. 2002, 4, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Bekal, S.; Niblack, T.L.; Lambert, K.N. A chorismate mutase from the soybean cyst nematode Heterodera glycines shows polymorphisms that correlate with virulence. Mol. Plant Microbe Interact. 2003, 16, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Doyle, E.A.; Lambert, K.N. Meloidogyne javanica chorismate mutase 1 alters plant cell development. Mol. Plant Microbe Interact. 2003, 16, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Dong, R.; Allen, R.; Davis, E.L.; Baum, T.J.; Hussey, R.S. Two chorismate mutase genes from the root-knot nematode Meloidogyne incognita. Mol. Plant Pathol. 2004, 6, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Lambert, K.N.; Bekal, S.; Domier, L.L.; Niblack, T.L.; Noel, G.R.; Smyth, C.A. Selection of Heterodera glycines chorismate mutase-1 alleles on nematode-resistant soybean. Mol. Plant Microbe Interact. 2005, 18, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, B.; Kast, P.; Haegeman, A.; Jacob, J.; Grunewald, W.; Gheysen, G. Structural and functional investigation of a secreted chorismate mutase from the plant-parasitic nematode Heterodera schachtii in the context of related enzymes from diverse origins. Mol. Plant Pathol. 2009, 10, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Grundler, F.; Betka, M.; Wyss, U. Influence of changes in the nurse cell system (syncytium) on sex determination and development of the cyst nematode Heterodera schachtii: Total amounts of proteins and amino acids. Physiol. Biochem. 1991, 81, 70–74. [Google Scholar]
- Jones, J.; Furlanetto, C.; Phillips, M. The role of flavonoids produced in response to cyst nematode infection of Arabidopsis thaliana. Nematology 2007, 9, 671–677. [Google Scholar] [CrossRef]
- Wuyts, N.; Lognay, G.; Swennen, R.; De Waele, D. Nematode infection and reproduction in transgenic and mutant Arabidopsis and tobacco with an altered phenylpropanoid metabolism. J. Exp. Bot. 2006, 57, 2825–2835. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-S.; Barker, K.R. Glyceollin I in soybean-cyst nematode interactions: Spatial and temporal distribution in roots of resistant and susceptible soybeans. Plant Physiol. 1991, 96, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.T.; Keen, N.T.; Thomason, I.J. Association of glyceollin with the incompatible response of soybean roots to Meloidogyne incognita. Physiol. Plant Pathol. 1980, 16, 309–318. [Google Scholar] [CrossRef]
- Kaplan, D.T.; Keen, N.T.; Thomason, I.J. Studies on the mode of action of glyceollin in soybean incompatibility to the root knot nematode, Meloidogyne incognita. Physiol. Plant Pathol. 1980, 16, 319–325. [Google Scholar] [CrossRef]
- Abawi, G.S.; Van Etten, H.D.; Mai, W.F. Phaseollin production induced by Pratylenchus penetrans in Phaseolus vulgaris. J. Nematol. 1971, 3, 301. [Google Scholar]
- Plowright, R.A.; Grayer, R.J.; Gill, J.R.; Rahman, M.L.; Harbornez, J.B. The induction of phenolic compounds in rice after infection by the stem nematode Ditylenchus angustus. Nematologica 1996, 42, 564–578. [Google Scholar] [CrossRef]
- Baldridge, G.D.; O’Neill, N.R.; Samac, D.A. Alfalfa (Medicago sativa L.) resistance to the root-lesion nematode, Pratylenchus penetrans: Defense-response gene mRNA and isoflavonoid phytoalexin levels in roots. Plant Mol. Biol. 1998, 38, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Soriano, I.R.; Asenstorfer, R.E.; Schmidt, O.; Riley, I.T. Inducible flavone in oats (Avena sativa) is a novel defense against plant-parasitic nematodes. Phytopathology 2004, 94, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.R.; Keen, N.T.; Thomason, I.J. Association of coumestans with the hypersensitivity of Lima bean roots to Pratylenchus scribneri. Physiol. Plant Pathol. 1977, 10, 105–116. [Google Scholar] [CrossRef]
- Collingborn, F.M.B.; Gowen, S.R.; Mueller-Harvey, I. Investigations into the biochemical basis for nematode resistance in roots of three Musa cultivars in response to Radopholus similis infection. J. Agric. Food Chem. 2000, 48, 5297–5301. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.J.; Niblack, T.L.; Krishnan, H.B. Infection by Heterodera glycines elevates isoflavonoid production and influenes soybean nodulation. J. Nematol. 1999, 31, 341–347. [Google Scholar] [PubMed]
- Brueske, C.H. Phenylalanine ammonia lyase activity in tomato roots infected and resistant to the root-knot nematode, Meloidogyne incognita. Physiol. Plant Pathol. 1980, 16, 409–414. [Google Scholar] [CrossRef]
- Edens, R.M.; Anand, S.C.; Bolla, R.I. Enzymes of the phenylpropanoid pathway in soybean infected with Meloidogyne incognita or Heterodera glycines. J. Nematol. 1995, 27, 292–303. [Google Scholar] [PubMed]
- Villeth, G.R.; Carmo, L.S.; Silva, L.P.; Fontes, W.; Grynberg, P.; Saraiva, M.; Mehta, A. Cowpea–Meloidogyne incognita interaction: Root proteomic analysis during early stages of nematode infection. Proteomics 2015, 15, 1746–1759. [Google Scholar] [CrossRef] [PubMed]
- Klink, V.P.; Hosseini, P.; Matsye, P.D.; Alkharouf, N.W.; Matthews, B.F. Syncytium gene expression in Glycine max[PI 88788] roots undergoing a resistant reaction to the parasitic nematode Heterodera glycines. Plant Physiol. Biochem. 2010, 48, 176–193. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, S.S.; Siddiqui, I.A.; Ali, N.A.; Ali, S.A.; Khan, G.H. Nematicidal and allelopathic responses of Lantana camara root extract. Phytopathol. Mediterr. 2003, 42, 71–78. [Google Scholar]
- Oliveira, J.T.; Araujo-Filho, J.H.; Grangeiro, T.B.; Gondim, D.M.; Segalin, J.; Pinto, P.M.; Vasconcelos, I.M. Enhanced synthesis of antioxidant enzymes, defense proteins and leghemoglobin in rhizobium-free cowpea roots after challenging with Meloidogyne incognita. Proteomes 2014, 2, 527. [Google Scholar] [CrossRef] [PubMed]
- Ithal, N.; Recknor, J.; Nettleton, D.; Hearne, L.; Maier, T.; Baum, T.J.; Mitchum, M.G. Parallel genome-wide expression profiling of host and pathogen during soybean cyst nematode infection of soybean. Mol. Plant-Microbe Interact. 2007, 20, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Hutangura, P.; Mathesius, U.; Jones, M.G.K.; Rolfe, B.G. Auxin induction is a trigger for root gall formation caused by root-knot nematodes in white clover and is associated with the activation of the flavonoid pathway. Funct. Plant Biol. 1999, 26, 221–231. [Google Scholar] [CrossRef]
- Wasson, A.P.; Ramsay, K.; Jones, M.G.K.; Mathesius, U. Differing requirements for flavonoids during the formation of lateral roots, nodules and root knot nematode galls in Medicago truncatula. New Phytol. 2009, 183, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Smant, G.; Helder, J.; Goverse, A. Parallel adaptations and common host cell responses enabling feeding of obligate and facultative plant parasitic nematodes. Plant J. 2018, 93, 686–702. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, T.; Vieira, P.; Gheysen, G.; de Almeida-Engler, J. Nematode feeding sites: Unique organs in plant roots. Planta 2013, 238, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.F. The ultrastructure and histochemistry of a nematode-induced giant cell. J. Biophys. Biochem. Cytol. 1961, 11, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Palomares-Rius, J.E.; Escobar, C.; Vovlas, A.; Castillo, P. Anatomical alterations in plant tissues induced by plant-parasitic nematodes. Front. Plant Sci. 2017, 8, 1987. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, J.; Díaz-Manzano, F.E.; Barcala, M.; Arganda-Carreras, I.; de Almeida-Engler, J.; Engler, G.; Fenoll, C.; Escobar, C. Phenotyping nematode feeding sites: Three-dimensional reconstruction and volumetric measurements of giant cells induced by root-knot nematodes in Arabidopsis. New Phytol. 2015, 206, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.J.; Lilley, C.J.; Knox, P.; Urwin, P.E. Syncytia formed by adult female Heterodera schachtii in Arabidopsis thaliana roots have a distinct cell wall molecular architecture. New Phytol. 2012, 196, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Peer, W.A.; Bandyopadhyay, A.; Blakeslee, J.J.; Makam, S.N.; Chen, R.J.; Masson, P.H.; Murphy, A.S. Variation in expression and protein localization of the PIN family of auxin efflux facilitator proteins in flavonoid mutants with altered auxin transport in Arabidopsis thaliana. Plant Cell 2004, 16, 1898–1911. [Google Scholar] [CrossRef] [PubMed]
- Peer, W.A.; Murphy, A.S. Flavonoids and auxin transport: Modulators or regulators? Trends Plant Sci. 2007, 12, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Stenlid, G. The effects of flavonoid compounds on oxidative phosphorylation and on the enzymatic destruction of indoleacetic acid. Physiol. Plant. 1963, 16, 110–120. [Google Scholar] [CrossRef]
- Balasubramanian, M.; Rangaswami, G. Presence of indole compound in nematode galls. Nature 1962, 194, 774–775. [Google Scholar] [CrossRef]
- Karczmarek, A.; Overmars, H.; Helder, J.; Goverse, A. Feeding cell development by cyst and root-knot nematodes involves a similar early, local and transient activation of a specific auxin-inducible promoter element. Mol. Plant Pathol. 2004, 5, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.; Perrine-Walker, F.; Wasson, A.; Mathesius, U. The control of auxin transport in parasitic and symbiotic root–microbe interactions. Plants 2015, 4, 606. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, T.; Goverse, A.; Haegeman, A.; Warmerdam, S.; Wanjau, C.; Jahani, M.; Engler, G.; de Almeida Engler, J.; Gheysen, G. Redirection of auxin flow in Arabidopsis thaliana roots after infection by root-knot nematodes. J. Exp. Bot. 2016, 67, 4559–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunewald, W.; Cannoot, B.; Friml, J.; Gheysen, G. Parasitic nematodes modulate PIN-mediated auxin transport to facilitate infection. PLoS Pathog. 2009, 5, e1000266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, M.; Rubery, P.H. Naturally occurring auxin transport regulators. Science 1988, 241, 346. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Han, K.; Heller, W.; Albert, A.; Dobrev, P.I.; Zažímalová, E.; Schäffner, A.R. Kaempferol 3-O-rhamnoside-7-O-rhamnoside is an endogenous flavonol inhibitor of polar auxin transport in Arabidopsis shoots. New Phytol. 2014, 201, 466–475. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Engler, J.; De Vleesschauwer, V.; Burssens, S.; Celenza, J.L.; Inzé, D.; Van Montagu, M.; Engler, G.; Gheysen, G. Molecular markers and cell cycle inhibitors show the importance of cell cycle progression in nematode-induced galls and syncytia. Plant Cell 1999, 11, 793–807. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Engler, J.; Gheysen, G. Nematode-induced endoreduplication in plant host cells: Why and how? Mol. Plant-Microbe Interact. 2012, 26, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Siddique, S.; Grundler, F.M.W. Metabolism in nematode feeding sites. In Advances in Botanical Research; Escobar, C., Fenoll, C., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 73, Chapter 5; pp. 119–138. [Google Scholar]
- De Almeida Engler, J.; Kyndt, T.; Vieira, P.; Van Cappelle, E.; Boudolf, V.; Sanchez, V.; Gheysen, G. CCS52 and DEL1 genes are key components of the endocycle in nematode-induced feeding sites. Plant J. 2012, 72, 185–198. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Engler, J.; Vieira, P.; Rodiuc, N.; Grossi de Sa, M.F.; Engler, G. The plant cell cycle machinery: Usurped and modulated by plant-parasitic nematodes. In Advances in Botanical Research; Escobar, C., Fenoll, C., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 73, Chapter 4; pp. 91–118. [Google Scholar]
- Li, Y.; Duan, S.; Jia, H.; Bai, C.; Zhang, L.; Wang, Z. Flavonoids from tartary buckwheat induce G2/M cell cycle arrest and apoptosis in human hepatoma HepG2 cells. Acta Biochim. Biophys. Sin. 2014, 46, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida Engler, J.; Rodiuc, N.; Smertenko, A.; Abad, P. Plant actin cytoskeleton re-modeling by plant parasitic nematodes. Plant Signal. Behav. 2010, 5, 213–217. [Google Scholar] [CrossRef]
- De Almeida Engler, J.; Van Poucke, K.; Karimi, M.; De Groodt, R.; Gheysen, G.; Engler, G.; Gheysen, G. Dynamic cytoskeleton rearrangements in giant cells and syncytia of nematode-infected roots. Plant J. 2004, 38, 12–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhl, M.; Tietze, S.; Sokoll, A.; Madathil, S.; Pfennig, F.; Apostolakis, J.; Fahmy, K.; Gutzeit, H.O. Flavonoids affect actin functions in cytoplasm and nucleus. Biophys. J. 2007, 93, 2767–2780. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Name of Enzyme/Gene/Metabolite | Flavonoid Accumulation Site | Suggested Flavonoid Function | Host Studied | Nematode Studied | Reference |
|---|---|---|---|---|---|
| Glyceollin | Roots | Glyceollin I and III accumulated more in the resistant cultivar compared with the susceptible cultivar. | Soybean, Glycine max | Soybean cyst nematode, Heterodera glycines | [64] |
| Stele in roots | Glyceollin was associated with the incompatible interaction between the resistant cultivar and M. incognita: accumulation was localised in the stele of resistant roots, high concentrations of glyceollin in resistant cultivar and glyceollin inhibited the motility of M. incognita. | Soybean, Glycine max | Root-knot nematodes, Meloidogyne incognita. and M. javanica | [65] | |
| Leaves | Glyceollin accumulated at sufficiently high concentrations at infection sites to result in a localised hypersensitive response. It inhibited nematode motility and respiration as well as plant tissue death via inhibition of mitochondrial electron transport system. | In vitro system | Root-knot nematodes, Meloidogyne incognita, and M. javanica | [66] | |
| Phaseollin | Hypocotyl and root | Phaseollin found only in P. penetrans infected tissue. The survival of P. penetrans juveniles incubated for 16 h in 47 µg/mL of phaseollin solution was unaffected. | Common bean, Phaseolus vulgaris | Root-lesion nematode, Pratylenchus penetrans | [67] |
| Sakuranetin | Leaf | Present only in resistant cultivars—suggested to be involved in defense | Rice | Stem nematode, Ditylenchus angustus | [68] |
| Roots, meristems, leaves | Isoflavonoid and pterocarpan (conjugate) accumulation correlated with nematode resistance. Medicarpin inhibited P. penetrans in a concentration dependent manner. | White clover, Trifolium repens | Stem nematode, Ditylenchus dipsaci | [47] |
| Lucerne, Medicago sativa | Stem nematode, Ditylenchus dipsaci | [52] | |||
| Lucerne, Medicago sativa | Root-lesion nematode, Pratylenchus penetrans | [69] | |||
| Roots and shoots during P. neglectus and H. avenae infection | Flavonoids possibly acted as broad defense compounds—induced in methyl jasmonate and nematode-treated plants. Plants treated with root extracts from methyl jasmonate-induced plants had reduced infection. | Oats, Avena sativa | Root lesion nematode, Pratylenchus neglectus, Cereal cyst nematode, Heterodera avenae, Stem nematode, Ditylenchus dipsaci | [70] |
| Roots | Coumesterol and psoralidin accumulated in roots and were localised at lesion sites caused by nematodes only in lima bean. Coumesterol significantly inhibited nematode motility at 10–15 µg/mL concentrations. | Lima bean, Phaseolus lunatus and snap bean, P. vulgaris | Root-lesion nematode, Pratylenchus scribneri | [71] |
| Adult female extracts | The yellow coloration in G. rostochiensis and G. pallida is attributed to flavonoid quercetagetin, which was present in pathotypes with yellow color and absent in paler pathotypes. | N/A | Potato cyst nematodes, Globodera rostochiensis and G. pallida | [48] |
| Roots | Flavan-3,4-diols and condensed tannins accumulated after nematode infection. | Banana, Musa | Burrowing nematode, Radopholus similis | [72] |
| Roots | Daidzein and genistein increased in susceptible Sussex cultivar at two and four days post inoculation. Isoflavonoid production was enhanced in nematode infected plants in susceptible Sussex and resistant Hartwig cultivar at two and three days post inoculation. | Soybean, Glycine max | Soybean cyst nematode, Heterodera glycines | [73] |
| Several compounds from the chalcone, flavone, flavanone, isoflavonoid and flavonol pathways. | Purified compounds and plant extracts. | Kaempferol, quercetin and myricetin repelled R. similis and M. incognita juveniles at 60–84 µg/mL. Luteolin, daidzein and genistein, repelled R. similis at 100–142 μg/mL. Kaempferol, quercetin, myricetin, rutin and quercitrin inhibited 13–41% of M. incognita juveniles after 48 h of incubation. Naringenin and hesperetin, apigenin, daidzein, and kaempferol reduced egg hatching in R. similis up to 21%. | N/A | Burrowing nematode, Radopholus similis, root-lesion nematode, Pratylenchus penetrans and root-knot nematode, Meloidogyne incognita | [41] |
| Purified compounds and marigold, Tagetes patula L. flower extracts | Patuletin killed 100% of nematodes at various dilutions after 72 h, whereas patulitrin killed 10–50% and quercetin killed 70–80% of nematodes. Rutin at 0.5–1% killed all nematodes within 24 h. | N/A | Corn cyst nematode, Heterodera zeae | [42] |
| (E)-chalcone | Purified compound | (E)-chalcone killed nematodes at 33 μM within 24 h and completely inhibited egg hatching at <10 μM within 15 days. | N/A | Potato cyst nematodes, Globodera rostochiensis and G. pallida | [46] |
| Phenylalanine ammonia lyase (PAL) | Roots | Plants grown at 27 °C had optimal PAL activity, which enabled the plants to synthesise phenylpropanoids used in nematode defense, as opposed to 32 °C, which inhibited enzyme activity. | Tomato, Lycopersi- cum esculentum | Root-knot nematode, Meloidogyne incognita | [74] |
| Roots | Gene expression levels generally induced by nematodes and higher in resistant plants. | Alfalfa, Medicago sativa | Root-lesion nematode, Pratylenchus penetrans | [69] |
| Soybean, Glycine max | Heterodera glycines and Meloidogyne incognita | [75] | |||
| Cowpea. Vigna unguiculata L. Walp | Meloidogyne incognita | [76] | |||
| Soybean, Glycine max genotype PI 88788 | Soybean cyst nematode, Heterodera glycines population NL1-RHg/HG-type 7 | [77] | |||
Flavonoid pathways:
| Roots | M. incognita reproduction was significantly higher in tobacco mutant with higher anthocyanidin content. M. incognita reproduction in Arabidopsis tt mutants and wild-type plants were similar. | Tobacco, Nicotiana tabacum and Arabidopsis thaliana | Root-knot nematode, M. incognita | [63] |
| Root extracts | Root extracts inhibited nematode motility, reduced nematode egg hatching and reduced gall numbers. | Lantana camara L. | Root-knot nematode, Meloidogyne javanica | [78] |
| Chalcone isomerase Auxin-induced protein | Roots | Chalcone isomerase protein as well as an auxin-induced protein were increased at 4, 5, and 6 days post inoculation. | Cowpea. Vigna unguiculata L. Walp | Root-knot nematode, Meloidogyne incognita | [79] |
| Root tissue | Upregulation of flavonoid synthesis and PIN 2 genes in nematode infected roots. | Soybean, Glycine max L. Merr. cv. Williams 82 | Soybean cyst nematode, Heterodera glycines Ichinohe | [80] |
| Root tissue | CHS1::gusA, CHS2::gusA and CHS3::gusA expressions overlapped with GH3::gusA expression at 48 h, 72 h and 120 h post inoculation. | White clover, Trifolium repens cv. Haifa | Root-knot nematode, Meloidogyne javanica | [81] |
| N/A | Flavonoid deficiency in tt (transparent testa) mutant lines of single and double tt4, tt5, and tt6 did not reduce the number of adult females, with several lines producing more female nematodes. | Arabidopsis thaliana | Sugar beet nematode, Heterodera schachtii | [62] |
| Chalcone synthase (silencing by RNA interference) | Root tissue | Flavonoid deficiency did not affect gall numbers. Flavonoid deficient roots had shorter galls and less pericycle cell division compared to roots with flavonoids. | Barrel medic, Medicago truncatula | Root-knot nematode, Meloidogyne javanica | [82] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chin, S.; Behm, C.A.; Mathesius, U. Functions of Flavonoids in Plant–Nematode Interactions. Plants 2018, 7, 85. https://doi.org/10.3390/plants7040085
Chin S, Behm CA, Mathesius U. Functions of Flavonoids in Plant–Nematode Interactions. Plants. 2018; 7(4):85. https://doi.org/10.3390/plants7040085
Chicago/Turabian StyleChin, Sabrina, Carolyn A. Behm, and Ulrike Mathesius. 2018. "Functions of Flavonoids in Plant–Nematode Interactions" Plants 7, no. 4: 85. https://doi.org/10.3390/plants7040085
APA StyleChin, S., Behm, C. A., & Mathesius, U. (2018). Functions of Flavonoids in Plant–Nematode Interactions. Plants, 7(4), 85. https://doi.org/10.3390/plants7040085