1. Introduction
Soybean (
Glycine max) is one of the most important crops grown throughout the world, and is a primary source of plant oil and protein for the human diet [
1]. In China, the soybean yield increases slowly in the past 50 years. Moreover, China imports more than 80% of the soybeans needed for the total domestic use, hence it is priority to increase the domestic production of soybean to make country self-sufficient [
2]. In this context, development of high yielding soybean cultivars is the key objective in ongoing plant breeding programmes. There is a positive correlation between plant height (PH) and pod number per plant, but taller plants are also more prone to lodging, which affects seed yield and quality. However, PH is an important agronomic trait that is closely related with seed yield in soybean [
3,
4], thereby making PH an important trait in soybean breeding program. However, PH is a complex quantitative trait controlled by multiple major or minor genes/QTLs, and is highly affected by the environment and other interactions [
5,
6]. Although, both conventional breeding and marker-assisted selection (MAS) have been successfully used to breed for PH. The latter is a more effective breeding method especially for environment-sensitive traits, and can be used to select plants at earlier seedling stages for those traits that are expressed late in the maturity stage [
7,
8]. Therefore, unrevealing the genetic basis of PH will greatly facilitates the implementation of MAS in breeding high-yielding soybean varieties.
In the past three decades, numerous studies have been carried out for elucidating the genetic control and QTLs composition of PH in soybean [
9,
10,
11,
12]. Till date, more than 200 QTLs were documented in SoyBase [
13], that are distributed on all 20 chromosomes of soybean, and have been detected in different populations and environments. Due to lack of high-throughput markers, the genetic maps used in most of the previous studies for QTL detection were based on low-throughput markers, such as restriction fragment length polymorphism (RFLP), amplified fragment length polymorphism (AFLP) and simple sequence repeat (SSR). As a result, most of these QTLs were mapped in large genomic regions, and have not been effectively used in MAS to breed varieties with ideal PH. Compared to low-throughput marker, the single nucleotide polymorphisms (SNPs) are the most abundant genetic polymorphism present in the plant genome and have high density as well as are evenly distributed across the whole genome. With the rapid advances in sequencing technologies and the completion of the whole genome sequencing of soybean
cv. Williams 82 [
14], SNP markers have become the marker of choice and has been now routinely used for high-density genetic map construction in soybean [
15]. These high-density linkage maps have greatly assisted in increasing QTL mapping resolution as well as provided convenience for QTL fine mapping and MAS for complex traits. For example, Li et al. [
16] used a soybean genetic map with an average distance of 0.72 cM between adjacent markers to detect QTLs for fatty acids and the results shown that more than 90% of QTL intervals were smaller than 5.0 cM. Similarly, Zhang et al. [
17] reported that the confidence interval of QTLs for tolerance to low-phosphorus stress in soybean could be significantly reduced by using a high-density genetic map.
Although some of the quantitative traits are controlled by only few major QTLs/genes, while the phenotypic variation for most of the complex traits is governed by many factors [
18]. In addition, two important factors i.e., QTL by QTL (epistatic) and QTL by environments interactions, contribute significantly to phenotypic variation of a complex trait [
19]. When the additive and epistatic effects as well as QTL by environment interaction effects associated with the target trait are reckoned in the QTL mapping model, the precision of QTL mapping would be greatly improved [
20]. Therefore, these factors cannot only be considered as the main obstacles to dissect the genetic architecture of complex traits, but also affect the accuracy of breeding value estimation and thus hindering the efficiency of breeding programs. Hence, these factors should be considered while dissecting the genetic basis of complex traits, and their use to improve plant performance. In recent years, epistatic and QTL by environment interaction effects have been considered in several crop species including soybean for QTL mapping [
3,
21,
22,
23,
24]. However, to our knowledge most of the previous studies on QTL mapping for PH in soybean focused mostly main-effect QTLs mapping [
25,
26,
27]. Therefore, efforts are required to study such QTL interaction effects for precise breeding of ideal PH in soybean.
In addition to the minor effect QTLs, PH was reported to be regulated by two major growth habit genes,
Dt1 and
Dt2 [
28,
29,
30]. In the Chinese Jang-Huai River Valley, soybean is always planted in early June and harvested at early October. In this region, soybean varieties have both determinate and indeterminate stem growth habits.
However, very little is known about the genetics of PH in the summer planting soybean genotypes grown in this region. By keeping the above into consideration, the present study has used three parents, M8206 (M), Tongshan (T) and Zhengyang (Z) with different growth habits to develop two related RIL populations viz., MT and ZM sharing a common indeterminate parent M8206. The objective of our study was to estimate the main-additive effects, epistatic effects and QTL by environment interaction effects of QTLs by utilizing the high-density linkage maps as well as phenotypic data collected from three different environments of two RIL populations to get detailed understanding of genetic basis for PH in summer planting soybean. This will further assist to better understand the implications of these QTLs while breeding for ideal PH through MAS method.
3. Discussion
Plant height (PH) is an important agronomic trait that is closely related to crop yield and quality in soybean [
4,
12]. Hence, developing soybean cultivars with desirable PH is an important objective of soybean breeders. However, to develop such soybean cultivars, it is imperative to have a detailed understanding of the genetic mechanism as well as genetic elements associated with PH. Plant height is a quantitative trait regulated by multiple genes, and is affected by environmental conditions [
26]. Although, over the past decades many QTLs related to soybean PH have been reported, and there are more than 200 QTLs documented for PH in the USDA Soybean Genome Database [
13]. However, most of these QTLs were not stable as well as confirmed due to small sized mapping population and low-density genetic map, and hence have not been useful for breeding PH in soybean. The quality of genetic maps has a great influence on the accuracy of QTL detection; high-density genetic map aided in the identification of more recombination events in a population as well as increased QTL mapping accuracy [
31,
32].
Moreover, little is known about the genetics of PH in summer planting soybean grown in the Chinese Jiang-Huai River Valley [
22]. Previous studies have shown that genetic polymorphisms between two soybean types are often very low [
33], and QTL detection is often limited in a single mapping population [
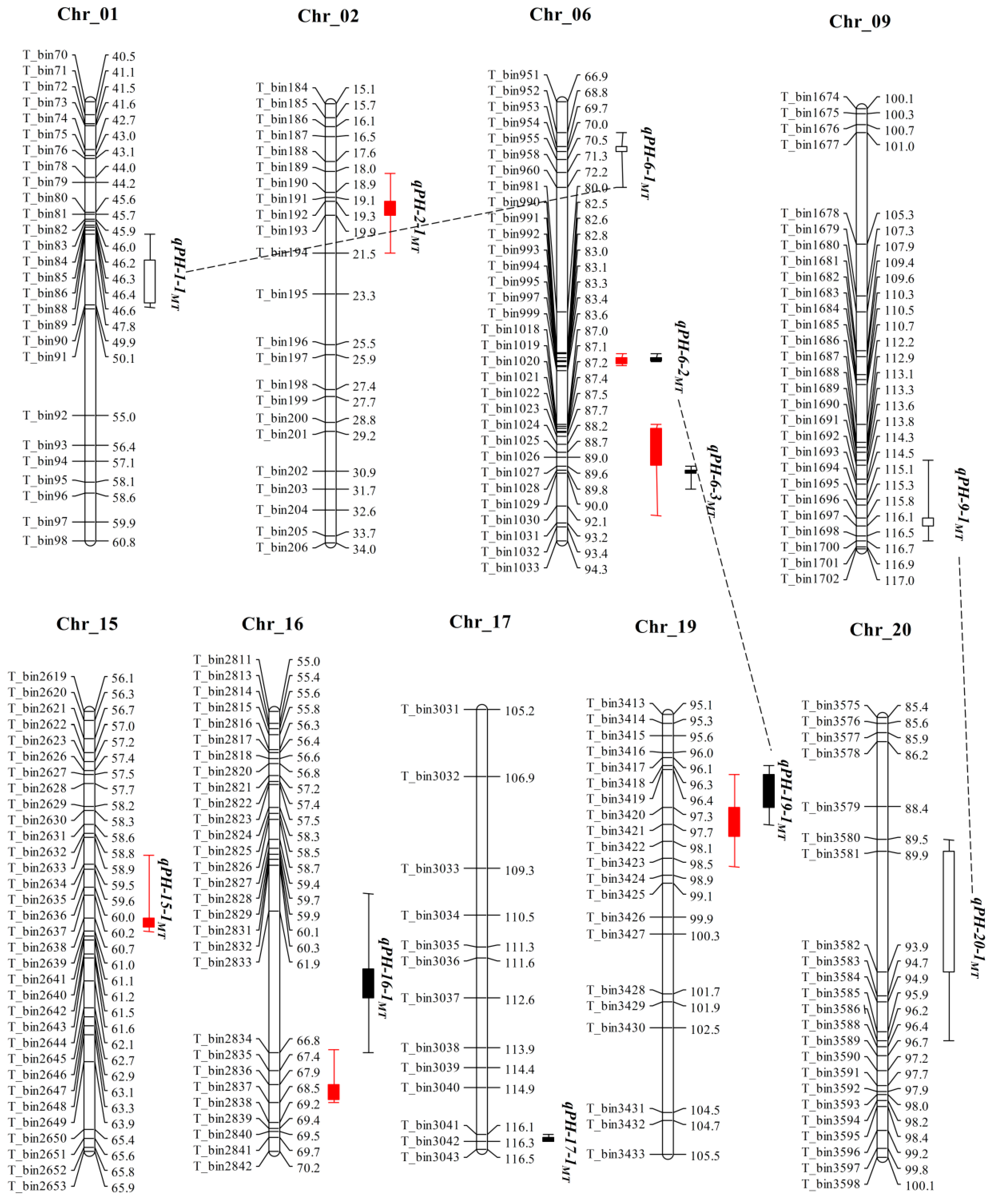
34]. This suggests that more diverse germplasm are needed for revealing the genetic basis of PH in this specific geographical region. In this regard, the present study utilized high-density linkage maps of two RIL populations viz., MT and ZM derived from diverse parents, and evaluated in multiple environments to identify the main-effect and epistatic effect QTLs as well as their interaction with the environment for PH in soybean. The results of our study showed that there were different QTLs in the two populations and most of QTLs were mapped in a small confidence interval (less than 3 cM), that greatly adds to the growing knowledge of the genetic control of PH (
Table 2 and
Table 3).
The QTLs associated with PH has been mapped on all 20 linkage groups/chromosomes of soybean [
13]. For cross validation and improving the accuracy of QTL mapping results, we used two different methods for QTL mapping including CIM and MCIM. A total of 8 and 12 additive effect QTLs were detected in MT and ZM populations (including the QTLs were detected in sub-populations), respectively (
Table 2,
Table 3 and
Table 5). Most of QTLs detected in our study overlapped with the loci available on SoyBase database [
13], and majority of the previously reported QTLs were identified in small physical genomic regions (
Table 2,
Table 3 and
Table 5), suggesting the importance of using high-resolution genetic map for QTL detection. In particular, the novel QTL,
QPH-17 was identified by comparing the mapping results of both CIM and MCIM methods and in MT and ZM populations (
Table 2,
Table 3 and
Table 5), indicating somehow distinct genetic architecture of MT and ZM populations and suggests the need to use more germplasm for revealing the complex genetic basis of PH in soybean. Finally, a continuous distribution and transgressive segregation were observed for PH in both RIL populations in all environments, which were consistent with the result of previous studies [
11,
12,
22], and implied that the existence of different allelic variations in the loci associated with plant height in the two parents of each RIL population. In fact, the results of this study also show that the positive alleles of the QTLs for PH were derived from both determinate and indeterminate parents in case of both MT and ZM populations. Some lines can pyramid different alleles and the additive effects of the QTLs controlling PH from both parents that make lines with a taller or smaller PH than their parents. Thus, we can develop cultivars with desirable PH by pyramiding different favorable alleles if we know the QTLs associated with PH in summer planting soybean, so it is important to note that not only the higher phenotype parent contributes positive alleles, but also the contribution of positive alleles by lower phenotype parent cannot be disregarded; and similar observation was made by Wang and Guan [
35], Miao et al. [
36] and Miao et al. [
37]. However, the soybean stem growth habit has a great influence on plant height. It is usually better to use the taller determinate cultivars as parents to develop a determinate line with larger plant height, but, we can also use the different QTLs associated with PH that independent of the
Dt1 locus to rapidly develop determinate or indeterminate stem growth habit cultivars. In the present study, the QTLs associated with PH that dependent and independent of the
Dt1 locus were detected. In particular, some QTLs could be detected in different environments and methods. Therefore, our findings provide important information for future soybean breeding programs.
It has been demonstrated that epistatic and QTL by environment interaction effects are the two important genetic factors that makes large contribution to the phenotypic variation observed in complex traits; and the knowledge of those interaction effects are important for understanding the genetic mechanism of complex traits [
19,
38]. Previous studies revealed that PH of soybean is significantly affected by environment [
11,
22]. Moreover, knowledge on specific QTL by environment interactions can guide for the search of varieties adapted to particular environments. The QTLs with greater additive effects are often more stable in multiple environments and populations [
39,
40]. For example,
QPH-19 (additive effect: 12.9) identified in multiple environments and populations; however,
qPH-5-1ZM (additive effect: 2.5) was detected in only one specific environment and population (ZM only) (
Table 3). The genetic architecture of PH also includes epistatic interactions between QTLs [
22]. Hence, ignoring inter-genic interaction will lead to overestimation of individual QTL effects and underestimation of genetic variance [
41]. This in turn could result substantial drop in the genetic response to MAS, particularly at late generations [
42]. In this study, six pairs of digeneic epistatic QTLs pairs were identified for PH in both populations, and explained a total of 3.5% and 6.5% of the phenotypic variation in MT and ZM populations, respectively (
Table 4). Out of these six pairs, four epistatic QTL pairs did not display additive effects alone, suggesting that these loci might serve as modifying genes that interact with other genes to affect the phenotype of plant height. All six pairs have significant AA but only two QTL pairs viz.,
qPH-6-2MT &
qPH-19-1MT/
QPH-19 and
qPH-2-2ZM &
qPH-4-1ZM possess significant AAE interaction effects. However, the total phenotypic variation explained by six epistatic pairs through AA effects was about 10%, and by AAE was 2.7%. Together, the effects of epistatic and environmental interactions (including AA, AE and AAE) could explain 13.9% and 14.7% of phenotypic variation in MT and ZM populations, respectively. Therefore, the results showed that epistatic and environmental interactions are important for understanding genetic basis of PH in soybean, suggesting that these effects should be considered in QTL mapping program, and will increase the accuracy of phenotypic value prediction in MAS.
In plant breeding, stability of QTL is essential for their use in marker-assisted breeding. Four pairs of QTLs viz.,
qPH-2-1MT &
qPH-2-1ZM (
QPH-2),
qPH-6-3MT &
qPH-6-2ZM (
QPH-6),
qPH-17-1MT &
qPH-17-1ZM (
QPH-17) and
qPH-19-1MT &
qPH-19-1ZM (
QPH-19) were detected in different populations in the same or overlapping physical position, could be considered the same locus. Out of these stable QTLs,
QPH-6 and
QPH-19 have
R2 > 10%, and were considered as the major and stable QTLs. However,
QPH-6 and
QPH-19 were identified in the physical interval that overlap with the previously identified QTLs
viz.,
Plant height 20-3 and
Plant height1-1, respectively as reported earlier by Gai et al. [
43] and Mansur et al. [
44]. The
QPH-17 was detected at the physical position of about 40–42 Mb and did not overlap with any of the previously reported PH QTLs, therefore, this stable QTL is been reported for the first time. The
QPH-2 was identified in the physical interval that overlaps with the previously identified QTLs
Plant height 13-1 [
45]. In addition to
QPH-2,
QPH-6,
QPH-17 and
QPH-19, some QTLs were identified in a single population through both mapping methods as well as multiple environments. For instance,
qPH-6-2MT was identified in MT population in three environments and overlap with the
Plant height 13-2 [
45]. The QTLs viz.,
qPH-9-1ZM and
qPH-10-1ZM were detected in two environments in ZM population, were adjacent or overlapping with QTLs as reported by Kim et al. [
46] and Wang et al. [
6]. Therefore, our results showed the reliability of QTL mapping in the present study, those QTLs can be used as important targets to identify the candidate genes and MAS in future studies.
It is of great interest for both theoretical study and practical breeding program to identify the actual candidate gene underlying the QTL region. Most of the earlier QTL mapping studies on PH did not mine for candidate genes [
11,
22,
26], and till date only few/limited genes related to PH have been isolated from soybean [
27,
29,
30]. Hence, based on the available information in current literature and gene annotation, the present study predicted some of the possible candidate genes for PH that underlies the major and stable QTLs in this study. For example,
Dt1 (
Glyma19g22160) gene lying within the physical genomic interval of
QPH-19 was predicted the candidate gene. Previous study has revealed that
Dt1 controls soybean growth habits with a significant impact on PH [
29]. In addition to the growth habit genes, the maturity and flowering time genes also have a great impact on plant height [
22,
47,
48,
49]. The physical intervals of
qPH-6-2MT,
qPH-10-1ZM and
qPH-16-1ZM contain
E1 [
48],
E2 [
50] and
E9 [
51] gene, respectively, and these genes might be the important candidate genes for these QTLs. However, the PH in soybean is known to be controlled by multiple genes. Some genes which have functions related to growth hormones or have functions that are directly or indirectly related to vegetative growth in stable QTLs were predicted as possible candidate genes. A total of 84 and 125 model genes were mined from the physical regions of the two other stable QTLs viz.,
QPH-6 and
QPH-17, and out of these 10 and 9 were considered as possible candidate genes based on the gene annotation and available literature (
Table S2). However, it needs further verification and functional validation to prove their actual role in the regulation of PH in soybean.



 ,
, 




{kind=link}
{kind=link}
{kind=link}