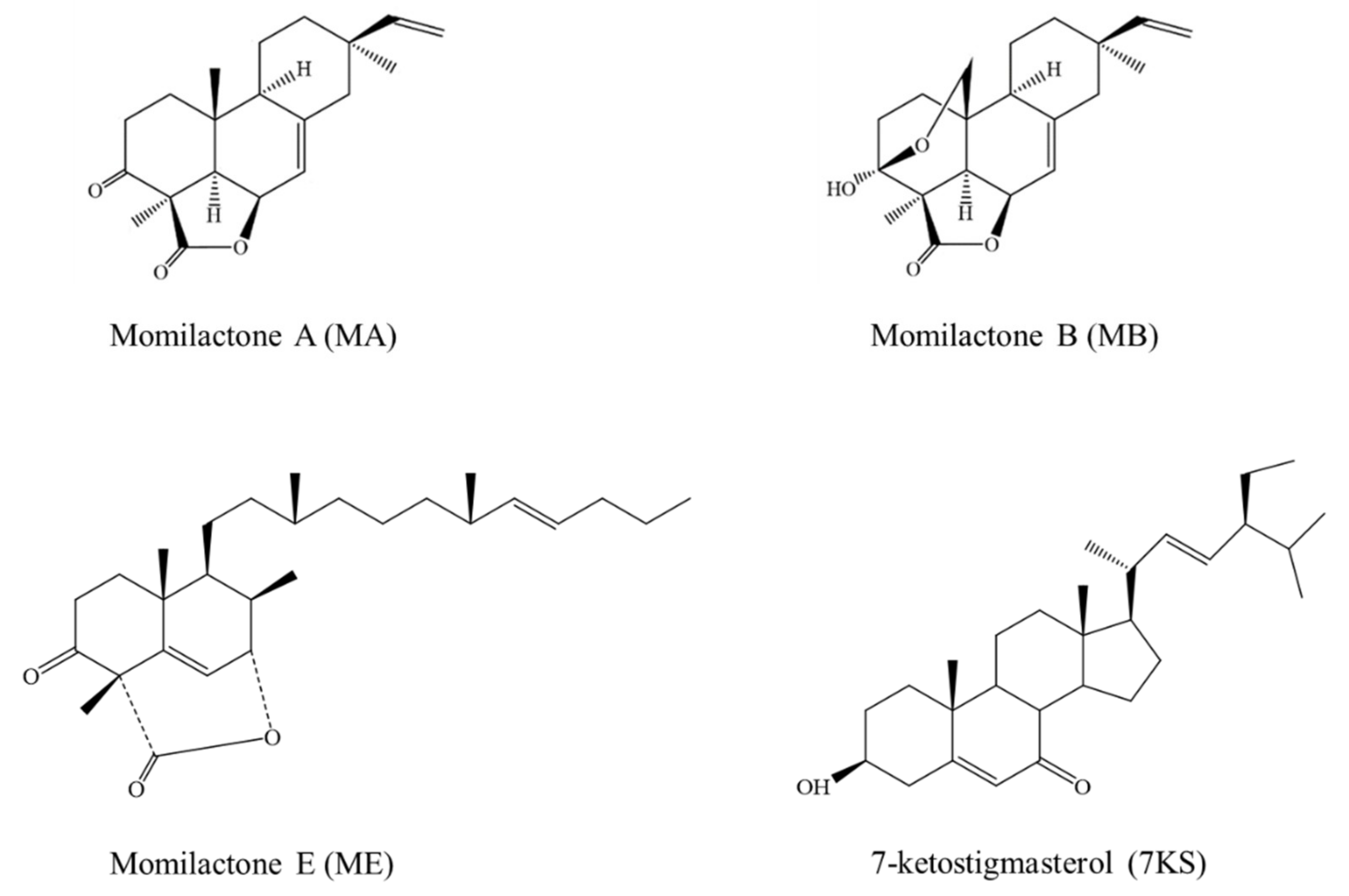
Inhibitory Activities of Momilactones A, B, E, and 7-Ketostigmasterol Isolated from Rice Husk on Paddy and Invasive Weeds




Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction and Isolation of Active Compounds from Rice Husk
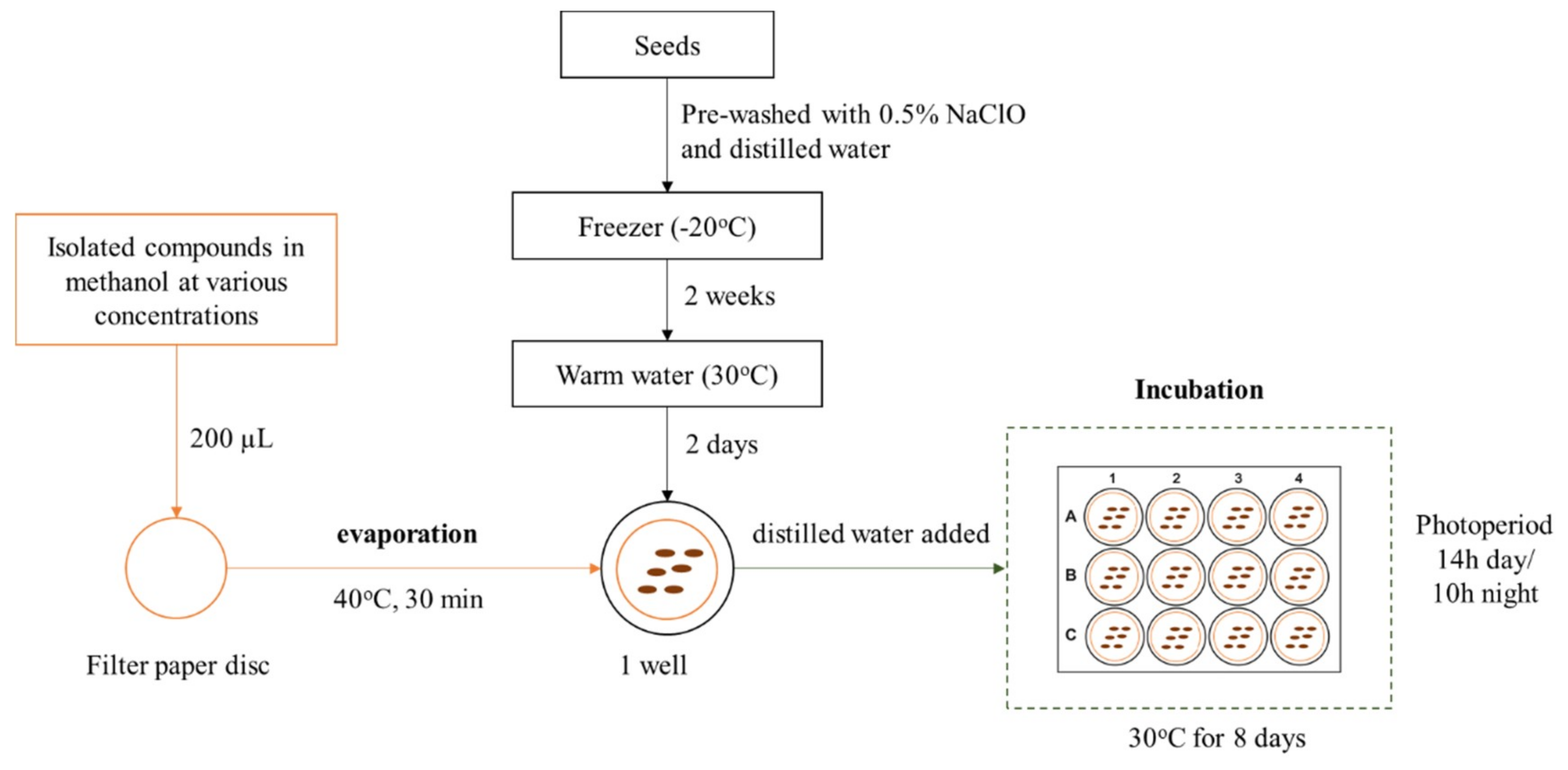
4.3. Allelopathic Bioassays
4.4. Quantification of Momilactones A and B by HPLC
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rice, E.L. Allelopathy—An Overview. In Chemically Mediated Interactions between Plants and Other Organisms. Recent Advances in Phytochemistry; Cooper-Driver, G.A., Swain, T., Conn, E.E., Eds.; Springer: Boston, MA, USA, 1985; Volume 19, pp. 81–99. [Google Scholar] [CrossRef]
- Xuan, T.D.; Elzaawely, A.A.; Deba, F.; Fukuta, M.; Tawata, S. Mimosine in Leucaena as a potent bio-herbicide. Agron. Sustain. Dev. 2006, 26, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Khanh, T.D.; Xuan, T.D.; Chung, I.M. Rice allelopathy and the possibility for weed management. Ann. Appl. Biol. 2007, 151, 325–339. [Google Scholar] [CrossRef]
- Xuan, T.D.; Tawata, S.; Khanh, T.D. Herbicidal activity of mimosine and its derivatives. In Herbicides—Advances in Research; Price, A., Kelton, J., Eds.; IntechOpen: Rijeka, Croatia, 2013; pp. 299–312. [Google Scholar] [CrossRef]
- Xuan, T.D.; Tsuzuki, E.; Tawata, S.; Khanh, T.D. Method to determine allelopathic potential of crop plants for weed control. Allelopathy J. 2004, 13, 149–164. [Google Scholar]
- Chung, I.M.; Ali, M.; Hahn, S.J.; Siddiqui, N.A.; Lim, Y.H.; Ahmad, A. Chemical constituents from the hulls of Oryza sativa with cytotoxic activity. Chem. Nat. Compd. 2005, 41, 182–189. [Google Scholar] [CrossRef]
- Esa, N.M.; Ling, T.B.; Peng, L.S. By-products of rice processing: An overview of health benefits and applications. J. Rice Res. 2013, 1, 107. [Google Scholar] [CrossRef]
- Khang, D.T.; Anh, L.H.; Ha, P.T.T.; Tuyen, P.T.; Quan, N.V.; Minh, L.T.; Quan, N.T.; Minh, T.N.; Xuan, T.D.; Khanh, T.D.; Trung, K.H. Allelopathic activity of dehulled rice and its allelochemicals on weed germination. Int. Lett. Nat. Sci. 2016, 58, 1–10. [Google Scholar] [CrossRef]
- Kato, T.; Kabuto, C.; Sasaki, N.; Tsunagawa, M.; Aizawa, H.; Fujita, K.; Kato, Y.; Kitahara, Y.; Takahashi, N. Momilactones, growth inhibitors from rice, Oryza sativa L. Tetrahedron Lett. 1973, 14, 3861–3864. [Google Scholar] [CrossRef]
- Berendji, S.; Asghari, J.B.; Matin, A.A. Allelopathic potential of rice (Oryza sativa) varieties on seedling growth of barnyardgrass (Echinochloa crus-galli). J. Plant Interact. 2008, 3, 175–180. [Google Scholar] [CrossRef]
- Minh, T.N.; Xuan, T.D.; Ahmad, A.; Elzaawely, A.A.; Teschke, R.; Van, T.M. Efficacy from different extractions for chemical profile and biological activities of rice husk. Sustainability 2018, 10, 1356. [Google Scholar] [CrossRef]
- Minh, T.N.; Xuan, T.D.; Ahmad, A.; Elzaawely, A.A.; Teschke, R.; Van, T.M. Momilactones A and B: Optimization of yields from isolation and purification. Separations 2018, 5, 28. [Google Scholar] [CrossRef]
- Quan, N.V.; Hoang-Dung, T.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 2019, 24, 482. [Google Scholar] [CrossRef] [PubMed]
- Quan, N.V.; Xuan, T.D.; Hoang-Dung, T.; Ahmad, A.; Khanh, T.D.; Dat, T.D. Contribution of momilactones A and B to diabetes inhibitory potential of rice bran: Evidence from in vitro assays. Saudi Pharm. J. 2019, in press. [Google Scholar] [CrossRef]
- Tsunakawa, M.; Ohba, A.; Sasaki, N.; Kabuto, C. Momilactone C, a minor constituent of growth inhibitors in rice husk. Chem. Lett. 1976, 1157–1158. [Google Scholar] [CrossRef]
- Cho, J.; Cha, B.; Min Lee, S.; Shrestha, S.; Jeong, R.; Sung Lee, D.; Kim, Y.; Lee, D.; Kang, H.; Kim, J.; Baek, N. Diterpenes from the roots of Oryza sativa L. and their inhibition activity on NO production in LPS-stimulated RAW264.7 macrophages. Chem. Biodivers. 2015, 12, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Xuan, T.D.; Minh, T.N.; Siddiqui, N.A.; Quan, N.V. Comparative extraction and simple isolation improvement techniques of active constituents’ momilactone A and B from rice husks of Oryza sativa by HPLC analysis and column chromatography. Saudi Pharm. J. 2019, 27, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Alemany, L.; Laparra, J.M.; Barberá, R.; Alegría, A. Evaluation of the cytotoxic effect of 7keto-stigmasterol and 7keto-cholesterol in human intestinal (Caco-2) cells. Food Chem. Toxicol. 2012, 50, 3106–3113. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Alfonso-García, A.; Alegría, A.; Barberá, R.; Cilla, A. 7keto-stigmasterol and 7keto-cholesterol induce differential proteome changes to intestinal epitelial (Caco-2) cells. Food Chem. Toxicol. 2015, 84, 29–36. [Google Scholar] [CrossRef]
- Marinho, R.d.S.S.; Ramos, C.J.B.; Leite, J.P.G.; Teixeira, V.L.; Paixão, I.C.N.d.P.; Belo, C.A.D.; Pereira, A.B.; Pinto, A.M.V. Antiviral activity of 7-keto-stigmasterol obtained from green Antarctic algae Prasiola crispa against equine herpesvirus 1. J. Appl. Phycol. 2017, 29, 555–562. [Google Scholar] [CrossRef]
- Khanh, T.D.; Cong, L.C.; Chung, I.M.; Xuan, T.D.; Tawata, S. Variation of weed-suppressing potential of Vietnamese rice cultivars against barnyardgrass (Echinochloa crus-galli) in laboratory, greenhouse and field screenings. J. Plant Interact. 2009, 4, 209–218. [Google Scholar] [CrossRef]
- Fujii, Y. Toxic chemicals from invasive alien plants. In Plant Toxins. Toxinology; Gopalakrishnakone, P., Carlini, C., Ligabue-Braun, R., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 25–36. [Google Scholar] [CrossRef]
- Kato, T.; Tsunakawa, M.; Sasaki, N.; Aizawa, H.; Fujita, K.; Kitahara, Y.; Takahashi, N. Growth and germination inhibitors in rice husks. Phytochemistry 1977, 16, 45–48. [Google Scholar] [CrossRef]
- Cartwright, D.W.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Isolation and characterization of two phytoalexins from rice as momilactones A and B. Phytochemistry 1981, 20, 535–537. [Google Scholar] [CrossRef]
- Fukuta, M.; Xuan, T.D.; Deba, F.; Tawata, S.; Khanh, T.D.; Chung, I.M. Comparative efficacies in vitro of antibacterial, fungicidal, antioxidant, and herbicidal activities of momilactones A and B. J. Plant Interact. 2007, 2, 245–251. [Google Scholar] [CrossRef]
- Xuan, T.D.; Shinkichi, T.; Khanh, T.D.; Chung, I.M. Biological control of weeds and plant pathogens in paddy rice by exploiting plant allelopathy: An overview. Crop Prot. 2005, 24, 197–206. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanh, T.D.; Xuan, T.D.; Chung, I.M.; Tawata, S. Allelochemicals of barnyardgrass-infested soil and their activities on crops and weeds. Weed Biol. Manag. 2008, 8, 267–275. [Google Scholar] [CrossRef]
- Fischer, N.H.; Quijano, L. Allelopathic agents from common weeds. In The Chemistry of Allelopathy; Thompson, A.C., Ed.; ACS Symposium Series 268–American Chemical Society: Washington, DC, USA, 1985; pp. 133–147. [Google Scholar] [CrossRef]
- Walck, J.L.; Baskin, J.M.; Baskin, C.C. A comparative study of the seed germination biology of a narrow endemic and two geographically-widespread species of Solidago (Asteraceae). 1. Germination phenology and effect of cold stratification on germination. Seed Sci. Res. 1997, 7, 47–58. [Google Scholar] [CrossRef]
- Meyer, A.H.; Schmid, B. Seed dynamics and seedling establishment invading perennial Solidago altissima under different experimental treatments. J. Ecol. 1999, 87, 28–41. [Google Scholar] [CrossRef]
- Sakata, Y.; Kaneko, S.; Hayano, A.; Inoue-Murayama, M.; Ohgushi, T.; Isagi, Y. Isolation and characterization of microsatellite loci in the invasive herb Solidago altissima (Asteraceae). Appl. Plant Sci. 2013, 1, 1200313. [Google Scholar] [CrossRef]
- Heath, J.J.; Kessler, A.; Woebbe, E.; Cipollini, D.; Stireman, J.O. Exploring plant defense theory in tall goldenrod, Solidago altissima. New Phytol. 2014, 202, 1357–1370. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample | L. sativa | E. crus-galli | S. altissima |
|---|---|---|---|
| MA | nd | 229.67 ± 1.20 b | 119.80 ± 11.50 b |
| MB | 178.46 ± 0.03 b | 31.88 ± 0.18 a | 20.36 ± 0.91 a |
| MAB | 327.20 ± 27.50 a | 28.26 ± 3.05 a | 23.97 ± 1.89 a |
| ME | nd | nd | 1162.50 ± 14.10 d |
| 7KS | nd | nd | nd |
| ME-7KS | nd | nd | 358.30 ± 8.39 c |
| pHA | nd | nd | 1074.20 ± 66.60 d |
| Sample | L. sativa | E. crus-galli | ||
|---|---|---|---|---|
| RL | SL | RL | SL | |
| MA | 348.00 ± 11.30 b | 388.20 ± 10.70 b | 147.60 ± 0.37 b | 123.87 ± 1.78 b |
| MB | 6.49 ± 0.04 a | 58.64 ± 0.99 a | 4.00 ± 0.03 a | 4.46 ± 0.05 a |
| MAB | 5.63 ± 0.05 a | 63.77 ± 0.64 a | 5.13 ± 0.05 a | 4.47 ± 0.05 a |
| pHA | 820.57 ± 9.8 c | 1453.30 ± 48.10 c | 936.66 ± 4.49 c | 1464.6 ± 40.30 c |
| Sample | Concentration (µg/mL) | L. sativa | E. crus-galli | S. altissima | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| GR | RL | SL | GR | RL | SL | GR | ||||
| Methanol | (8/8) a 0.0 | (1.95) b 0.0 | (0.76) c 0.0 | (8/8) 0.0 | (4.53) 0.0 | (3.03) 0.0 | (13/20) 0.0 | |||
| MA | 400 | 0.0 | 52.38 | 50.67 | 78.57 | 95.96 | 96.70 | 86.75 | ||
| MB | 100.00 | 94.71 | 84.59 | 100.00 | 100.00 | 100.00 | 100.00 | |||
| MAB | 58.33 | 91.82 | 82.82 | 100.00 | 100.00 | 100.00 | 100.00 | |||
| ME | 1000 | 0.0 | +92.68 | 6.61 | 0.0 | +16.48 | +8.01 | 39.53 | ||
| 7KS | 0.0 | +60.93 | 13.65 | nd | nd | nd | nd | |||
| ME-7KS | 0.0 | +93.00 | 5.71 | 0.0 | 13.33 | 29.52 | 94.66 | |||
| pHA | 0.0 | 63.14 | 37.01 | 0.0 | 58.46 | 36.12 | 44.66 | |||
| Parameters | This Study | Previous Study [13] |
|---|---|---|
| Sep-Pak C18 cartridge | Used | Not use |
| HPLC column | 10 µm, 150 mm × 4.6 mm i.d. | 10 µm, 250 mm × 4.6 mm i.d. |
| Flow rate | 0.5 mL/min | 0.4 mL/min |
| LOD | 0.05 ng/mL (MA), 0.48 ng/mL (MB) | 0.43 ng/mL (MA), 0.18 ng/mL (MB) |
| LOQ | 0.14 ng/mL (MA), 1.46 ng/mL (MB) | 1.31 ng/mL (MA), 0.54 ng/mL (MB) |
| Quantity of MA | 51.96 µg/mL | 16.44 µg/mL |
| Quantity of MB | 42.33 µg/mL | 9.24 µg/mL |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, N.V.; Xuan, T.D.; Tran, H.-D.; Dieu Thuy, N.T. Inhibitory Activities of Momilactones A, B, E, and 7-Ketostigmasterol Isolated from Rice Husk on Paddy and Invasive Weeds. Plants 2019, 8, 159. https://doi.org/10.3390/plants8060159
Quan NV, Xuan TD, Tran H-D, Dieu Thuy NT. Inhibitory Activities of Momilactones A, B, E, and 7-Ketostigmasterol Isolated from Rice Husk on Paddy and Invasive Weeds. Plants. 2019; 8(6):159. https://doi.org/10.3390/plants8060159
Chicago/Turabian StyleQuan, Nguyen Van, Tran Dang Xuan, Hoang-Dung Tran, and Nguyen Thi Dieu Thuy. 2019. "Inhibitory Activities of Momilactones A, B, E, and 7-Ketostigmasterol Isolated from Rice Husk on Paddy and Invasive Weeds" Plants 8, no. 6: 159. https://doi.org/10.3390/plants8060159
APA StyleQuan, N. V., Xuan, T. D., Tran, H. -D., & Dieu Thuy, N. T. (2019). Inhibitory Activities of Momilactones A, B, E, and 7-Ketostigmasterol Isolated from Rice Husk on Paddy and Invasive Weeds. Plants, 8(6), 159. https://doi.org/10.3390/plants8060159