Distribution and Redistribution of 109Cd and 65Zn in the Heavy Metal Hyperaccumulator Solanum nigrum L.: Influence of Cadmium and Zinc Concentrations in the Root Medium



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
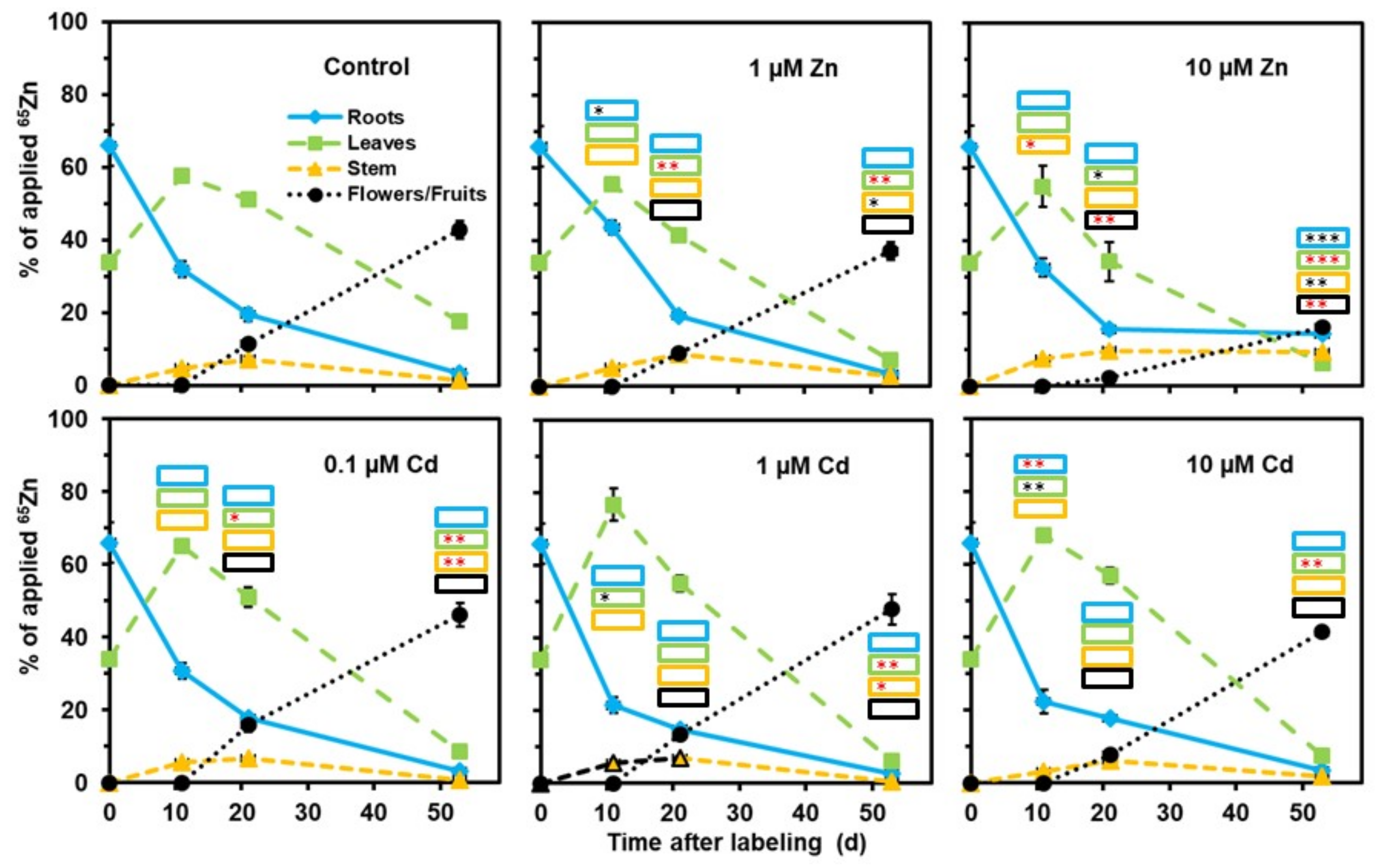
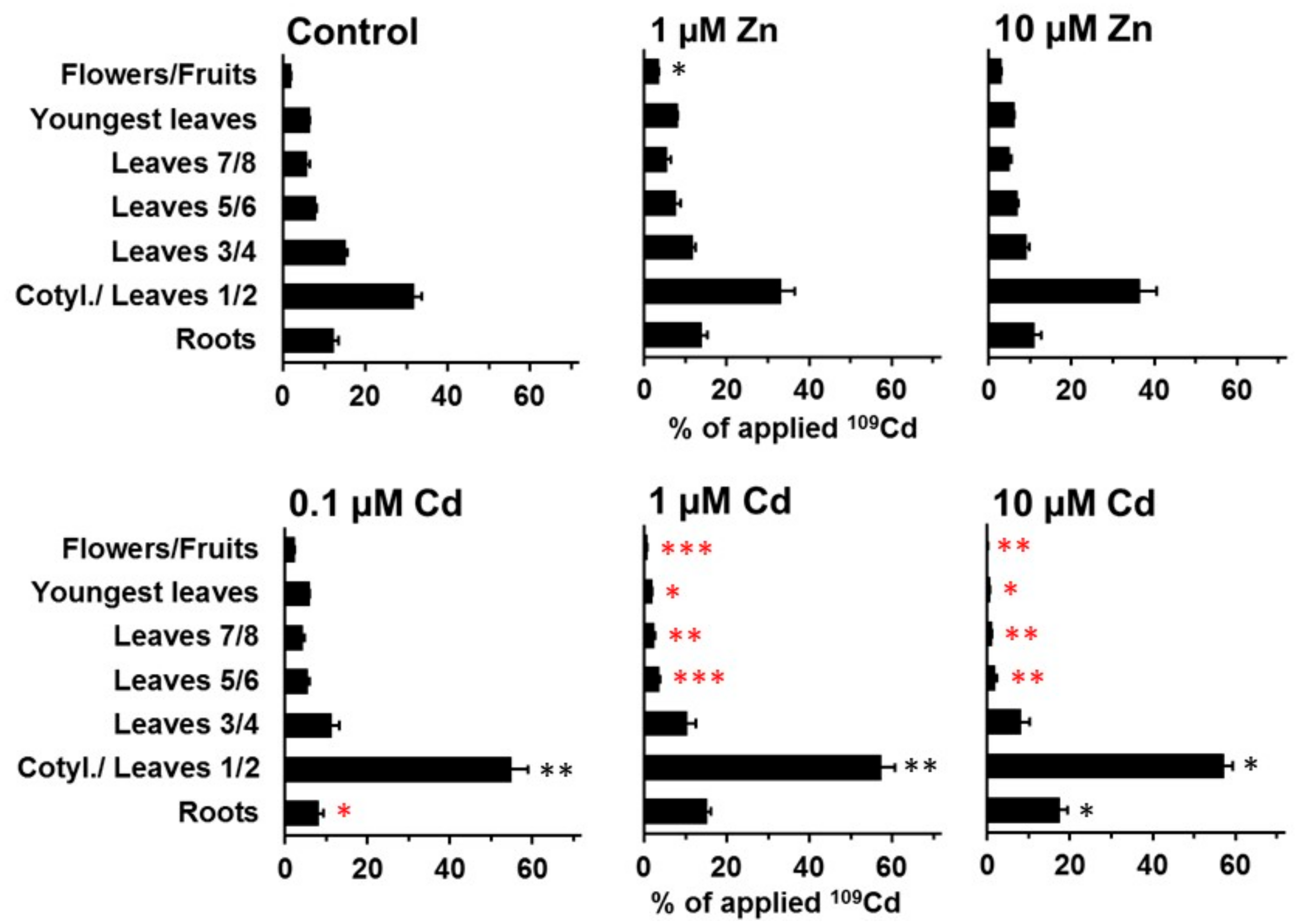
2. Results
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grotz, N.; Guerinot, M.L. Molecular aspects of Cu, Fe and Zn homeostasis in plants. Biochim. Biophys. Acta-Mol. Cell Res. 2006, 1763, 595–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rascio, N.; Vecchia, F.D.; La Rocca, N.; Barbato, R.; Pagliano, C.; Raviolo, M.; Gonnelli, C.; Gabbrielli, R. Metal accumulation and damage in rice (cv. Vialone nano) seedlings exposed to cadmium. Environ. Exp. Bot. 2008, 62, 267–278. [Google Scholar] [CrossRef]
- Bouis, H.E.; Welch, R.M. Biofortification-A sustainable agricultural strategy for reducing micronutrient malnutrition in the global south. Crop Sci. 2010, 50, S20–S32. [Google Scholar] [CrossRef]
- Page, V.; Feller, U. Heavy Metals in Crop Plants: Transport and Redistribution Processes on the Whole Plant Level. Agronomy 2015, 5, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Pence, N.S.; Larsen, P.B.; Ebbs, S.D.; Letham, D.L.D.; Lasat, M.M.; Garvin, D.F.; Eide, D.; Kochian, L.V. The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator Thlaspi caerulescens. Proc. Natl. Acad. Sci. USA 2000, 97, 4956–4960. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Fen g, Y.; He, Z.L.; Stoffella, P.J. Molecular mechanisms of heavy metal hyperaccumulation and phytoremediation. J. Trace Elem. Med. Biol. 2005, 18, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Kramer, U. Metal hyperaccumulation in plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Wei, S.; Anders, I.; Feller, U. Selective uptake, distribution, and redistribution of Cd-109, Co-57, Zn-65, Ni-63, and Cs-134 via xylem and phloem in the heavy metal hyperaccumulator Solanum nigrum L. Environ. Sci. Pollut. Res. 2014, 21, 7624–7630. [Google Scholar] [CrossRef]
- Demirezen, D.; Aksoy, A. Heavy metal levels in vegetables in Turkey are within safe limits for Cu, Zn, Ni and exceeded for Cd and Pb. J. Food Qual. 2006, 29, 252–265. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Stasinos, S.; Nasopoulou, C.; Tsikrika, C.; Zabetakis, I. The bioaccumulation and physiological effects of heavy metals in carrots, onions, and potatoes and dietary implications for Cr and Ni: A review. J. Food Sci. 2014, 79, R765–R780. [Google Scholar] [CrossRef] [PubMed]
- Kramer, U.; Chardonnens, A.N. The use of transgenic plants in the bioremediation of soils contaminated with trace elements. Appl. Microbiol. Biotechnol. 2001, 55, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.H.; Zhou, Q.X.; Koval, P.V. Flowering stage characteristics of cadmium hyperaccumulator Solanum nigrum L. and their significance to phytoremediation. Sci. Total Environ. 2006, 369, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Memon, A.R.; Schroder, P. Implications of metal accumulation mechanisms to phytoremediation. Environ. Sci. Pollut. Res. 2009, 16, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Herren, T.; Feller, U. Transport of cadmium via xylem and phloem in maturing wheat shoots: Comparison with the translocation of zinc, strontium and rubidium. Ann. Bot. 1997, 80, 623–628. [Google Scholar] [CrossRef]
- Riesen, O.; Feller, U. Redistribution of nickel, cobalt, manganese, zinc and cadmium via the phloem in young and maturing wheat. J. Plant Nutr. 2005, 28, 421–430. [Google Scholar] [CrossRef]
- Mari, S.; Gendre, D.; Pianelli, K.; Ouerdane, L.; Lobinski, R.; Briat, J.-F. Root-to-shoot long-distance circulation of nicotianamine and nicotianamine-nickel chelates in the metal hyperaccumulator Thlaspi Caerulescens. J. Exp. Bot. 2006, 57, 4111–4122. [Google Scholar] [CrossRef]
- Page, V.; Weisskopf, L.; Feller, U. Heavy metals in white lupin: Uptake, root-to-shoot transfer and redistribution within the plant. New Phytol. 2006, 171, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; et al. OsHMA3, a P-1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef]
- Page, V.; Feller, U. Selective transport of zinc, manganese, nickel, cobalt and cadmium in the root system and transfer to the leaves in young wheat plants. Ann. Bot. 2005, 96, 425–434. [Google Scholar] [CrossRef]
- Feller, U.; Anders, I.; Wei, S. Effects of PEG-Induced Water Deficit in Solanum nigrum on Zn and Ni Uptake and Translocation in Split Root Systems. Plants 2015, 4, 284–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, V.; Feller, U. Light intensity selectively influences the distribution and further redistribution of macroand micronutrients in hydroponically grown wheat (Triticum aestivum L.). J. Plant. Nutr. 2016, 39, 428–437. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, C.; Chai, S.; Shuai, W.; Sha, L.; Zhang, H.; Kang, H.; Fan, X.; Zeng, J.; Zhou, Y.; et al. Ammonium N influences the uptakes, translocations, subcellular distributions and chemical forms of Cd and Zn to mediate the Cd/Zn interactions in dwarf polish wheat (Triticum polonicum L.) seedlings. Chemosphere 2018, 193, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Cheng, Y.R.; Shuai, W.D.; Zeng, J.; Kang, H.Y.; Fan, X.; Sha, L.N.; Zhang, H.Q.; Wang, Y.; Zhou, Y.H. Nitrate N influences the accumulations and subcellular distributions of Cd and Zn to mediate the Cd/Zn interactions in dwarf Polish Wheat (Triticum polonicum L.) seedlings. Soil Sci. Plant Nutr. 2018, 137–147. [Google Scholar] [CrossRef]
- Simek, I.; Tuma, I.; Feller, U. Accumulation, distribution and redistribution of 109Cd and 65Zn in wheat (Triticum aestivum L.) and bean (Phaseolus vulgaris L.) plants on nutrient media with elevated zinc or cadmium. J. Elem. 2019, 24, 1129–1141. [Google Scholar] [CrossRef]
- Edelstein, M.; Ben-Hur, M. Heavy metals and metalloids: Sources, risks and strategies to reduce their accumulation in horticultural crops. Sci. Hort. 2018, 234, 431–444. [Google Scholar] [CrossRef]
- Luo, L.; Shen, Y.; Wang, X.; Chu, B.; Xu, T.; Liu, Y.; Zeng, Y.; Liu, J. Phytoavailability, bioaccumulation, and human health risks of metal(loid) elements in an agroecosystem near a lead-zinc mine. Environ. Sci. Pollut. Res. 2018, 25, 24111–24124. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, N.P.; Walsh, K.B.; Baker, A.J.M. Detection and quantification of ligands involved in nickel detoxification in a herbaceous Ni hyperaccumulator Stackhousia tryonii Bailey. J. Exp. Bot. 2005, 56, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Sagardoy, R.; Morales, F.; Rellen-Alvarez, R.; Abadia, A.; Abadia, J.; Lopez-Millan, A.F. Carboxylate metabolism in sugar beet plants grown with excess Zn. J. Plant Physiol. 2011, 168, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Stolt, J.P.; Sneller, F.E.C.; Bryngellson, T.; Lundborg, T.; Schat, H. Phytochelatin and cadmium accumulation in wheat. Environ. Exp. Bot. 2003, 49, 21–28. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feller, U.; Anders, I.; Wei, S. Distribution and Redistribution of 109Cd and 65Zn in the Heavy Metal Hyperaccumulator Solanum nigrum L.: Influence of Cadmium and Zinc Concentrations in the Root Medium. Plants 2019, 8, 340. https://doi.org/10.3390/plants8090340
Feller U, Anders I, Wei S. Distribution and Redistribution of 109Cd and 65Zn in the Heavy Metal Hyperaccumulator Solanum nigrum L.: Influence of Cadmium and Zinc Concentrations in the Root Medium. Plants. 2019; 8(9):340. https://doi.org/10.3390/plants8090340
Chicago/Turabian StyleFeller, Urs, Iwona Anders, and Shuhe Wei. 2019. "Distribution and Redistribution of 109Cd and 65Zn in the Heavy Metal Hyperaccumulator Solanum nigrum L.: Influence of Cadmium and Zinc Concentrations in the Root Medium" Plants 8, no. 9: 340. https://doi.org/10.3390/plants8090340