1. Introduction
Tomato (
Solanum lycopersicum), which originated in sub-tropical areas [
1], is sensitive to chilling temperature [
2,
3]. However, its production areas usually experience sub-optimal temperatures (which range between two thresholds, a minimum (8–12 °C) and an optimum (25–27 °C)) [
4,
5,
6]. Sub-optimal temperatures affect the vegetative and fruit growth of tomato plants, which ultimately leads to decline in the yield [
1]. Hence, to improve tomato production and breeding under these conditions, it is necessary to understand the mechanisms underlying tomato responses to sub-optimal temperatures.
Plants can respond and tolerate to cold stress by changing several biochemical processes and traits features [
7]. These changes include alterations in the composition, structure, and function of the plasma membrane; synthesis of cryoprotectant molecules, including soluble sugars and low-molecular-weight nitrogenous compounds, such as proline; and increases in the scavenging activity of reactive oxygen species (ROS) [
8,
9]. Peroxidase (POD) is one of the most important antioxidative enzymes, and can alleviate oxidative damage by scavenging ROS under stress conditions [
10,
11]. Cold temperatures also induce cascades of alterations in metabolic pathways, including the phenylpropanoid pathway, and in the activities of antioxidant enzymes [
12]. As one of the most important inorganic osmotica in plants, potassium (K) plays key role in establishing the osmotic adjustment ability [
13,
14]. The available evidence suggest that K improves plant survival rate under cold stress by reducing ROS accumulation and increasing antioxidant levels [
15,
16]. A significant negative correlation has been found between the leaf K concentration and frost damage, and an adequate K supply can effectively increase the frost resistance of plants, whereas K deficiency can increase the susceptibility of plants to low temperatures [
17,
18,
19]. The K requirement of greenhouse tomato plants is very high for vegetative growth and fruit production [
20], but little is known regarding the K
+ uptake characteristics of two tomato cultivars differing in cold resistance under sub-optimal temperature conditions.
In plant roots, K
+ uptake from the outer environment is mainly mediated by K
+ channels and transporters [
21,
22,
23]. All abiotic stress results in disturbance to K
+ homeostasis and provokes a feedback control on K
+ channels and transporters expression and post-translational regulation of their activity, optimizing K
+ absorption and usage [
24,
25,
26]. However, few studies have focused on the expression of genes related to K
+ uptake and translocation during exposure to sub-optimal temperature stress. Although some studies have reported that low temperature decreases the uptake [
27] and influx [
26] of K
+, it is still unclear whether sub-optimal temperatures affect K
+ uptake and translocation by affecting the expression of genes coding for K
+ channels and transporters.
RNA sequencing (RNA-seq) is a critical method for profiling global gene expression in response to biotic and abiotic stress [
28,
29]. In tomato, transcriptome profiling has been used for comparative analysis of expression of defense related genes between two tomato cultivars with different resistances to salt, cold and disease stress [
30,
31,
32]. However, to the best of our knowledge, a transcriptome analysis of tomato under sub-optimal temperature conditions that focuses on the root system has not been reported yet.
In this study, we profiled differences in plant growth, K+ uptake, and gene expression changes by making comparison between one cold-sensitive and one cold-tolerant tomato cultivar exposed to sub-optimal temperature conditions. We hypothesized that (1) under sub-optimal temperature conditions, the plant growth rate (PGR) and the POD activity of roots in cold-tolerant cultivar would be higher than that in the cold-sensitive cultivar; and (2) the cold-tolerant cultivar would exhibit enhanced cold tolerance by regulating not only the expression of genes related to cold tolerance in roots, but also K+ uptake. As mentioned earlier, K+ uptake from outside is mainly mediated by K+ channels and transporters; thus, we also hypothesized that (3) sub-optimal temperatures could inhibit the expression of genes coding for K+ channels and transporters in roots, and that the inhibitory effect on cold-sensitive tomato would be stronger than that on cold-tolerant tomato.
3. Discussion
Decreasing the greenhouse temperatures affects different aspects of tomato growth and development [
1]. The comparison performed in this study showed that the phenotype of S708, including the IPH, and PGR was more significantly affected than that of T722 by sub-optimal temperature treatment (
Figure 1). The results described above suggest that the T722 cultivar was more tolerant to sub-optimal temperature than the S708 cultivar. Moreover, the T722 cultivar had a more developed root system than the S708 cultivar under sub-optimal temperature condition (
Figure 1c,d), which indicated that the T722 cultivar has a stronger nutrient uptake ability than the S708 cultivar [
34]. MDA content was used to evaluate the cell membrane damage [
35]. In our study, the MDA content was significantly higher in S708 than in T722 under sub-optimal temperature condition (
Figure 2a). This result suggested that the T722 suffered less membrane damage than S708 under sub-optimal temperature stress.
Low temperature may lead to osmotic stress because low temperature impairs water absorption and transport [
36]. K
+ plays a key role in the establishment of a plant’s ability to adjust to osmotic changes and in improving the cold tolerance of plants [
14]. However, K
+ uptake characteristics of tomatoes in response to sub-optimal temperatures are still not very clear. Our results showed that the whole plant K
+ amount and the net K
+ uptake rate were significantly higher in T722 than in S708 under sub-optimal temperature condition (
Table 2 and
Figure 3a). Many studies have indicated that there was a significant positive correlation between the K
+ amount and plant resistance to abiotic stress [
17,
18,
19]. A sufficient K supply can reduce the formation of ROS in plant cells and alleviate the damage to plants exposed to chilling or freezing stress [
15]. Therefore, we deduced that the higher K
+ amount observed in T722 might contribute to the defense of the plants against sub-optimal temperature stress. Moreover, we found that sub-optimal temperature induced a reduction in the K
+ transportation ratio in both cultivars during the 5–10 days period (
Figure 3b). This reduction might have caused the K
+ content in roots to significantly increase and that in stems to significantly decrease in both cultivars at 5 DAT (
Table 2).
To investigate the differences in gene expression between the T722 and S708 cultivars in response to sub-optimal temperature, we performed a comparative transcriptome analysis. The overall number of DEGs in cold-tolerant cultivar was higher than that in cold-sensitive cultivar at 5 DAT (
Figure 5). This result is different from the previous study [
35], and this might be related to the characteristics of the cultivar used in experiment. Different cultivars showed different cold responses at the transcriptional level. The DEGs may reveal the difference in the response to cold stress between the S708 and T722 cultivars. GO enrichment analysis is helpful for highlighting the main biological processes in response to stress environment [
37]. In our study, we found that the genes involved in ROS homeostasis and hormone metabolic were differentially expressed between S708 and T722 under sub-optimal temperature condition. These results showed that hormone and ROS may plant key roles in regulating gene expression in response to sub-optimal temperature stress.
The cultivar-specific DEGs in T722 were considered crucial to its higher cold tolerance. Among them, most DEGs were involved in the “plant hormone signal transduction” and “phenylpropanoid biosynthesis” pathways. Plant hormones are known to regulate plant growth and development and thus allow the adaptation of plants to abiotic stress [
38]. Our results showed that 11 genes in the significantly enriched KEGG pathway, “plant hormone signal transduction” were induced only in T722 at 5 DAT (
Table 4). Among these genes, two auxin-related genes, including Solyc09g065850.2, which encodes indole-3-acetic acid (IAA)-induced protein IAA3, were up-regulated only in T722. In
Arabidopsis, mutation of this gene can affect auxin-dependent root growth and lateral root formation [
39]. Moreover, Solyc04g078470.2, which encodes a homologue of
Arabidopsis CYCD3, responds to cytokinins and is required for proper secondary root thickening [
40]. These results might be linked to the stronger root system in T722 than in S708 under sub-optimal temperature stress. Solyc08g008600.2, involved in the jasmonic acid (JA) signaling pathway and encodes a transcription factor MYC2, which positively regulates the formation of lateral roots [
41] and tolerance to oxidative stress in
Arabidopsis [
42]. This gene was strongly induced by sub-optimal temperature only in T722, which suggested that its up-regulation might contribute to cold tolerance in tomato. This result is consistent with the findings obtained in previous studies [
15], which showed that JA might act as a positive regulator in response to cold stress in tomato. Three genes (Solyc01g106640.2, Solyc10g085960.1 and Solyc09g007020.1) involved in the salicylic acid (SA) signaling pathway that encode the pathogenesis-related (PR) protein PR1 were more strongly induced by sub-optimal temperature in T722 than in S708 (
Table 4). PR proteins such as β-1,3-glucanase exhibit antifreeze activity and even prevent the formation of intracellular ice [
43]. These results showed that the plant hormones IAA, SA and JA might play their role in regulating root growth and in resisting sub-optimal temperature in T722.
Further results showed that 12 peroxidase related genes involved in the significantly enriched KEGG pathway “phenylpropanoid biosynthesis” were significantly induced only in T722 at 5 DAT (
Table 5). This result is consistent with the results of POD activity which significantly increased in T722 under sub-optimal temperature (
Figure 2b). ROS are produced by plants upon exposure to low-temperature stress [
44,
45], and peroxidase is one of the most important antioxidative enzymes, and can alleviate oxidative damage by scavenging ROS under stress conditions [
10,
11]. Moreover, we found that three genes encoding mitogen-activated protein kinase (MAPK) (Solyc12g005360.1, Solyc02g090980.1 and Solyc05g008020.2) were induced only in T722 during exposure to sub-optimal temperatures (
Table S6). MAPK can enhance
Nicotiana tabacum total peroxidase (POD) activity, thereby inducing less accumulation of H
2O
2 and alleviation of ROS-mediated injuries [
46], which might at least partly explain the large number of peroxidase related genes that were up-regulated in T722. K
+ efflux is an important physiological phenomenon, that is usually detected almost instantaneously after the application of a stress factor and lasts from a few minutes to several hours [
47]. Our results showed that the T722 cultivar had a lower K
+ efflux rate than S708 cultivar at 5 hours and 5 days after treatment (
Figure 4), which is consistent with the previous results reported by Chen et al. (2005) [
48], who indicated that the cold-tolerant tomato root system has a strong ability to maintain K
+. K
+ efflux is mainly induced by stresses and ROS. The present results suggested that the sub-optimal temperature treatment induced the expression of genes coding for MAPK and peroxidase in T722, and increased the POD activity, which might contribute to ROS scavenging and reduced loss of K
+.
It worth noting that almost the same number of peroxidase-related genes involved in the “phenylpropanoid biosynthesis” pathway were repressed in T722 (
Table S5). Peroxidases are involved in lignification, cell elongation, stress defense and seed germination. The diverse peroxidase activities facilitate opposing reactions in plants, such as generation/scavenging of ROS and loosening/stiffening of the cell wall [
49]. Similar results were also found in cytochrome P450 pathway; there were lots of up- and down-regulated glutathione S transferases (GSTs) involved in cytochrome P450 pathway, and these genes play roles in detoxifying oxidative-stress metabolites [
50]. The expression of defense genes has negative effects on plant growth, which to some degree counterbalances their positive effects [
34]. So we deduced that the up- and down-regulated defense related genes in T722 might help to maintain energy balance. Moreover, our results showed that eight sulfur metabolism-related genes were up-regulated in both cultivars. Sulfur assimilation is a platform for the biosynthesis of sulfur-containing defense compounds (SDCs), which are crucial for the survival of plants under biotic and abiotic stress [
51].
To clarify the change of K
+ content in response to sub-optimal temperatures, the expression of genes coding for K
+ uptake and translocation was analyzed (
Table 6). The results showed that Solyc12g005670.1, the K
+ transporter SIHAK5, and a homologue of the
O. sativa K
+ transporter HAK26 (Solyc06g051830.1), was repressed in both cultivars at 5 DAT. Moreover, under sub-optimal temperature conditions, a homologue of the
Arabidopsis K
+ channel AKT1 (Solyc09g005220.1) was repressed only in S708. Similarly, the expression of another gene coding for
S. lycopersicum K
+ channel LKT1 (Solyc12g006850.1) was down-regulated 2.11-fold in S708 and 1.44-fold in T722 (
Table 6). The Shaker type K
+ channel LKT1 in tomato belongs to the
Arabidopsis AKT1 subfamily, and HAK5 belongs to the KT/HAK/KUP transporter family, which plays an important role in K
+ uptake [
52,
53,
54]. Then, we verified the expression of these genes by qRT-PCR, and they were consistent with the RNA-seq data, despite the differential expression folds (
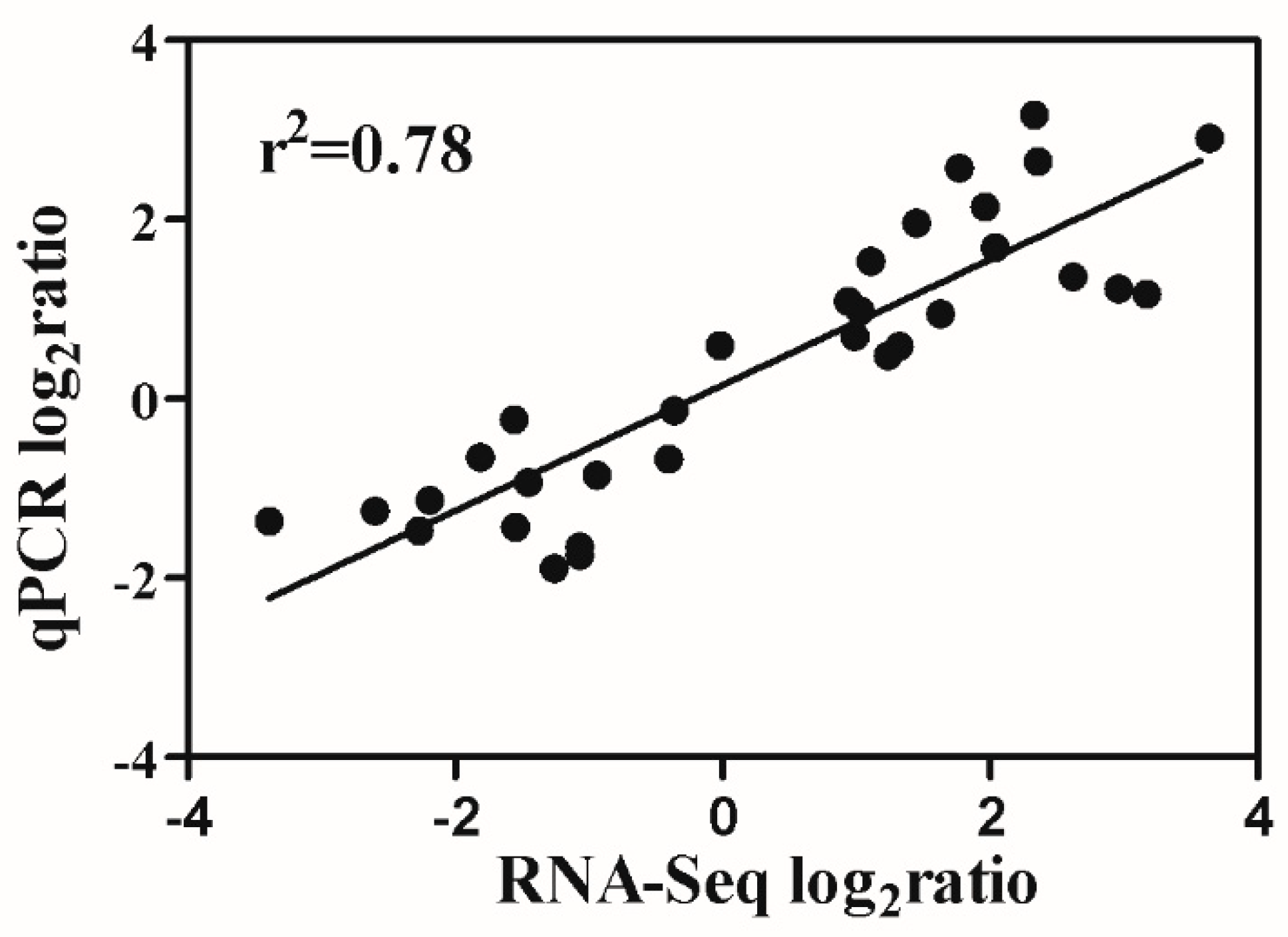
Figure 6).
The phosphorylation of K
+ channels and transporters is the most important mechanism for the regulation of their function [
22]. Several studies have demonstrated that the CBL-CIPK complex can regulate not only the expression of
AKT1, but also the expression of
AtHAK5 [
55,
56,
57,
58]. Our results show that a homologue of the
O. sativa calcineurin B-like protein CBL (Solyc03g083320.2) and two homologues of the
O. sativa CBL-interacting protein kinase CIPK (Solyc09g042660.2 and Solyc03g006110.2) were induced only in T722 (
Table 6). Our findings suggest that sub-optimal temperatures might repress K
+ net uptake rate and K
+ influx by inhibiting the expressions of gene coding for K
+ channel and transporter and the CBL-CIPK pathway may be more active in T722 during sub-optimal temperature stress. Moreover, we found that Solyc08g007060.2, which encodes a homologue of the
Arabidopsis nitrate transporter NPF7.3/NRT1.5, was inhibited in both cultivars by the sub-optimal temperature treatment (
Table 6). The encoded protein, which can regulate K
+ translocation to the shoots in
Arabidopsis [
59,
60], was repressed by sub-optimal temperature. That may explain the decreased K
+ transportation ratio in both cultivars during the 5–10 days after treatment (
Figure 3b). In addition, there are close couplings between aquaporins and K-channel transporters in water uptake of roots [
20] and osmoregulation [
61]. Our transcriptome and qPCR analysis showed that sub-optimal temperature inhibits not only genes related to K
+ uptake and translocation, but also aquaporins (
Table S6 and
Figure 6), which may be another reason for the increased K
+ accumulation in roots under sub-optimal temperature conditions. Extensive studies have shown the roles of K
+ uptake and transport related genes in resistance to salt and low potassium stress [
22,
62,
63], and the roles of these genes in the regulation of cold resistance should be analyzed in further studies.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}