Gravity Signaling in Flowering Plant Roots



{kind=link}
Abstract
1. Introduction
2. Gravity Sensing in Roots
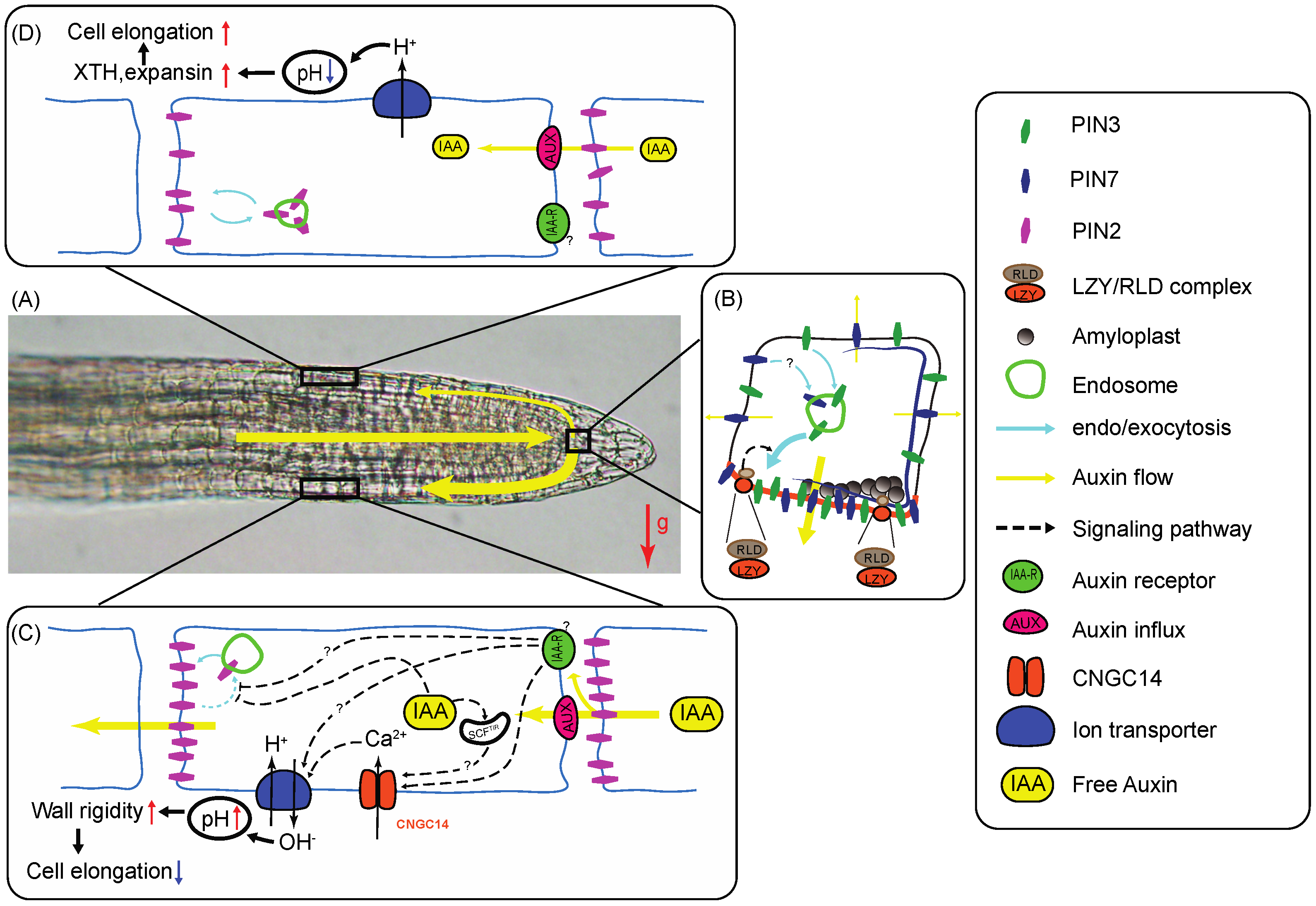
2.1. The Columella Cells Contribute to Root Gravity Sensing
2.2. Gravity Sensing in the Statocytes Involves Unknown Receptors
3. Gravity Signal Transduction in the Root Statocytes
3.1. Auxin Transport in Plant Tissues Involves Auxin Influx and Efflux Carriers
3.2. Gravity-Signal Transducers Contribute to Vesicular Trafficking and Plastid Function
4. Auxin Gradient Propagation from Root Cap to EZ
5. The Gravitropic Curvature Results from Differential Cell Elongation between Opposite Flanks of the Root EZ
6. The GSA of Lateral Roots Differs from That of Primary Roots
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Xiao, G.; Wang, X.; Zhang, X.; Friml, J. Evolution of fast root gravitropism in seed plants. Nat. Commun. 2019, 10, 3480–3490. [Google Scholar] [CrossRef] [PubMed]
- Barlow, P. Gravity perception in plants—A multiplicity of systems derived by evolution. Plant Cell Environ. 1995, 18, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Kobayashi, Y.; Hase, Y.; Watanabe, H. Positional effect of cell inactivation on root gravitropism using heavy-ion microbeams. J. Exp. Bot. 2002, 53, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Tsugeki, R.; Fedoroff, N. Genetic ablation of root cap cells in Arabidopsis. Proc. Natl. Acad. Sci. USA 1999, 96, 12941–12946. [Google Scholar] [CrossRef] [PubMed]
- Blancaflor, E.; Fasano, J.; Gilroy, S. Mapping the functional roles of cap cells in the response of Arabidopsis primary roots to gravity. Plant Physiol. 1998, 116, 213–222. [Google Scholar] [CrossRef]
- Leitz, G.; Kang, B.-H.; Schoenwaelder, M.; Staehelin, L. Statolith sedimentation kinetics and force transduction to the cortical endoplasmic reticulum in gravity-sensing Arabidopsis columella cells. Plant Cell 2009, 21, 843–860. [Google Scholar] [CrossRef]
- Haberlandt, G. Ueber die perzeption des geotropischen reizes. Ber. Dtsch. Bot. Ges. 1900, 18, 261–272. [Google Scholar]
- Nemec, B. Ueber die art der wahrnehmung des schwekraftreizes bei den pflanzen. Ber. Dtsch. Bot. Ges. 1900, 18, 241–245. [Google Scholar]
- Baldwin, K.; Strohm, A.; Masson, P. Gravity sensing and signal transduction in vascular plant primary roots. Am. J. Bot. 2013, 100, 126–142. [Google Scholar] [CrossRef]
- Kiss, J.; Hertel, R.; Sack, F. Amyloplasts are necessary for full gravitropic sensitivity in roots of Arabidopsis thaliana. Planta 1989, 177, 198–206. [Google Scholar] [CrossRef]
- Kiss, J.; Wright, J.; Caspar, T. Gravitropism in roots of intermediate-starch mutants of Arabidopsis. Physiol. Plant 1996, 97, 237–244. [Google Scholar] [CrossRef] [PubMed]
- MacCleery, S.; Kiss, J. Plastid sedimentation kinetics in roots of wild-type and starch-deficient mutants of Arabidopsis. Plant Physiol. 1999, 120, 183–192. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Band, L.; Wells, D.; Larrieu, A.; Sun, J.; Middleton, A.; French, A.; Brunoud, G.; Sato, E.; Wilson, M.; Péret, B.; et al. Root gravitropism is regulated by a transient lateral auxin gradient controlled by a tipping-point mechanism. Proc. Natl. Acad. Sci. USA 2012, 109, 4668–4673. [Google Scholar] [CrossRef] [PubMed]
- Fitzelle, K.; Kiss, J. Restoration of gravitropic sensitivity in starch-deficient mutants of Arabidopsis by hypergravity. J. Exp. Bot. 2001, 52, 265–275. [Google Scholar] [CrossRef]
- Vitha, S.; Yang, M.; Sack, F.; Kiss, J. Gravitropism in starch-excess mutant of Arabidopsis thaliana. Am. J. Bot. 2007, 94, 590–598. [Google Scholar] [CrossRef]
- Kuznetsov, O.; Hasenstein, K. Intracellular magnetophoresis of amyloplasts and induction of root curvature. Planta 1996, 198, 87–94. [Google Scholar] [CrossRef]
- Kuznetsov, O.; Hasenstein, K. Magnetophoretic induction of curvature in coleoptiles and hypocotyls. J. Exp. Bot. 1997, 48, 1951–1957. [Google Scholar] [CrossRef]
- Zheng, Z.; Zou, J.; Li, H.; Xue, S.; Wang, Y.; Le, J. Microrheological insights into the dynamics of amyloplasts in root gravity-sensing cells. Mol. Plant 2014, 8, 660–663. [Google Scholar] [CrossRef][Green Version]
- Hou, G.; Mohamalawari, D.; Blancaflor, E. Enhanced gravitropism of roots with a disrupted cap actin cytoskeleton. Plant Physiol. 2003, 131, 1360–1373. [Google Scholar] [CrossRef][Green Version]
- Yamamoto, K.; Pyke, K.; Kiss, J. Reduced gravitropism in inflorescence stems and hypocotyls, but not in roots, of Arabidopsis mutants with large plastids. Physiol. Plant 2002, 114, 627–636. [Google Scholar] [CrossRef]
- Okamoto, K.; Ueda, H.; Shimada, T.; Tamura, K.; Kato, T.; Tasaka, M.; Morita, M.; Hara-Nishimura, I. Regulation of organ straightening and plant posture by an actin-myosin XI cytoskeleton. Nat. Plants 2015, 1, 15031. [Google Scholar] [CrossRef] [PubMed]
- De Bang, L.; Paez-Garcia, A.; Cannon, A.; Chin, S.; Kolape, J.; Liao, F.; Sparks, J.; Jiang, Q.; Blancaflor, E. Brassinosteroids inhibit autotropic root straightening by modifying filamentous-actin organization and dynamics. Front. Plant. Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.; Edelmann, R.; Wood, P. Gravitropism of hypocotyls of wild-type and starch-deficient Arabidopsis seedlings in spaceflight studies. Planta 1999, 209, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Wolverton, C.; Mullen, J.; Ishikawa, H.; Evans, M. Root gravitropism in response to a signal originating outside of the cap. Planta 2002, 215, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, S.; Barlow, P.; Volkmann, D.; Baluska, F. Actin turnover-mediated gravity response in maize root apices. Gravitropism of decapped roots implicates gravisensing outside of the root cap. Plant Signal. Behav. 2006, 1, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Staves, M.; Wayne, R.; Leopold, A. Hydrostatic pressure mimics gravitational pressure in characean cells. Protoplasma 1992, 168, 141–152. [Google Scholar] [CrossRef]
- Staves, M.; Wayne, R.; Leopold, A. The effect of external medium on the gravitropic curvature of rice (Oryza sativa, Poaceae) roots. Am. J. Bot. 1997, 84, 1522–1529. [Google Scholar] [CrossRef]
- Soga, K. Resistance of plants to gravitational force. J. Plant Res. 2013, 126, 589–596. [Google Scholar] [CrossRef]
- Herranz, R.; Vandenbrink, J.; Villacampa, A.; Manzano, A.; Poehlman, W.; Feltus, F.; Kiss, J.; Medina, F. RNAseq analysis of the response of Arabidopsis thaliana to fractional gravity under blue-light stimulation during spaceflight. Front. Plant Sci. 2019, 10, 1529. [Google Scholar] [CrossRef]
- Sievers, A.; Buchen, B.; Volkmann, D.; Hejnowicz, Z. Role of the cytoskeleton in gravity perception. In The Cytoskeletal Basis of Plant Growth and Form; Lloyd, C., Ed.; Acedemic Press: London, UK, 1991; pp. 169–182. [Google Scholar]
- Pickard, B.; Ding, J. The mechanosensory calcium-selective ion channel: Key component of a plasmalemmal control centre? Aust. J. Plant Physiol. 1993, 20, 439–459. [Google Scholar] [CrossRef]
- Hamilton, E.; Schlegel, A.; Haswell, E. United in diversity: Mechanosensitive ion channels in plants. Annu. Rev. Plant Biol. 2015, 66, 113–137. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, H.; Toyota, M.; Furuichi, T.; Sokabe, M. Calcium mobilizations in response to changes in the gravity vector in Arabidopsis seedlings. Possible cellular mechanisms. Plant Signal. Behav. 2014, 9, e29099. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hattori, T.; Otomi, Y.; Nakajima, Y.; Soga, K.; Wakabayashi, K.; Iida, H.; Hoson, T. MCA1 and MCA2 are involved in the response to hypergravity in Arabidopsis hypocotyls. Plants 2020, 9, 590. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, W.; Trewavas, A. Calcium in gravitropism: A re-examination. Planta 1997, 203, S85–S90. [Google Scholar] [CrossRef]
- Lu, Y.-T.; Feldman, L. Light-regulated root gravitropism: A role for, and characterization of, a calcium/calmodulin-dependent protein kinase homolog. Planta 1997, 203, S91–S97. [Google Scholar] [CrossRef]
- Monshausen, G.; Miller, N.; Murphy, A.; Gilroy, S. Dynamics of auxin-dependent Ca2+ and pH signaling in root growth revealed by integrating high-resolution imaging with automated computer vision-based analysis. Plant J. 2011, 65, 309–318. [Google Scholar] [CrossRef]
- Perera, I.; Hung, C.-Y.; Brady, S.; Muday, G.; Boss, W. A universal role for inositol 1,4,5-trisphosphate-mediated signaling in plant gravitropism. Plant Physiol. 2006, 140, 746–760. [Google Scholar] [CrossRef]
- Andreeva, Z.; Barton, D.; Armour, W.; Li, M.; Liao, L.; McKellar, H.; Pethybridge, K.; Marc, J. Inhibition of phospholipase C disrupts cytoskeletal organization and gravitropic growth in Arabidopsis roots. Planta 2010, 232, 1263–1279. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, W.; Chen, X.; Xue, H. The role of Arabidopsis 5PTase13 in root gravitropism through modulation of vesicle trafficking. Cell Res. 2009, 19, 1191–1204. [Google Scholar] [CrossRef]
- Munnik, T.; Nielsen, E. Green light for polyphosphoinositide signals in plants. Curr. Opin. Plant Biol. 2011, 14, 489–497. [Google Scholar] [CrossRef]
- Plieth, C.; Trewavas, A. Reorientation of seedlings in the Earth’s gravitational field induces cytosolic calcium transients. Plant Physiol. 2002, 129, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Furuichi, T.; Tatsumi, H.; Sokabe, M. Cytoplasmic calcium increases in response to changes in the gravity vector in hypocotyls and petioles of Arabidopsis seedlings. Plant Physiol. 2008, 146, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Bizet, F.; Pereda-Loth, V.; Chauvet, H.; Gérard, J.; Eche, B.; Girousse, C.; Courtade, M.; Perbal, G.; Legué, V. Both gravistimulation onset and removal trigger an increase of cytoplasmic free calcium in statocytes of roots grown in microgravity. Sci. Rep. 2018, 8, 11442. [Google Scholar] [CrossRef] [PubMed]
- Chauvet, H.; Pouliquen, O.; Forterre, Y.; Legué, V.; Moulia, B. Inclination, not force, is sensed by plants during shoot gravitropism. Sci. Rep. 2016, 6, 35431. [Google Scholar] [CrossRef]
- Limbach, C.; Hauslage, J.; Schäfer, C.; Braun, M. How to activate a plant gravireceptor. Early mechanisms of gravity sensing studied in Characean rhizoids during parabolic flights. Plant Physiol. 2005, 139, 1030–1040. [Google Scholar] [CrossRef]
- Strohm, A.; Barrett-Wilt, G.; Masson, P. A functional TOC complex contributes to gravitty signal transduction in Arabidopsis. Front. Plant Sci. 2014, 5, 148. [Google Scholar] [CrossRef] [PubMed]
- Bérut, A.; Chauvet, H.; Legué, V.; Moulia, B.; Pouliquen, O.; Forterre, Y. Gravisensors in plant cells behave like an active granular liquid. Proc. Natl. Acad. Sci. USA 2018, 115, 5123–5128. [Google Scholar] [CrossRef]
- Pouliquen, O.; Forterre, Y.; Bérut, A.; Chauvet, H.; Bizet, F.; Legué, V.; Moulia, B. A new scenario for gravity detection in plants: The position sensor hypothesis. Phys. Biol. 2017, 14, 035005. [Google Scholar] [CrossRef]
- Raven, J. Transport of indoleacetic acid in plant cells in relation to pH and electrical potential gradients, and its significance for polar IAA transport. New Phytol. 1975, 74, 163–172. [Google Scholar] [CrossRef]
- Bennett, M.; Marchant, A.; Green, H.; May, S.; Ward, S.; Millner, P.; Walker, A.; Schulz, B.; Feldmann, K. Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 1996, 273, 948–950. [Google Scholar] [CrossRef]
- Noh, B.; Murphy, A.; Spalding, E. Multidrug resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development. Plant Cell 2001, 13, 2441–2454. [Google Scholar]
- Geisler, M.; Blakeslee, J.; Bouchard, R.; Lee, O.; Vincenzetti, V.; Bandyopadhyay, A.; Titapiwatanakun, B.; Peer, W.; Bailly, A.; Richards, E.; et al. Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1. Plant J. 2005, 44, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Mellor, N.; Voß, U.; Janes, G.; Bennett, M.; Wells, D.; Band, L. Auxin fluxes through plasmodesmata modify root-tip auxin distribution. Development 2020, 147, dev181669. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Wisniewska, J.; Benkova, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Kleine-Vehn, J.; Ding, Z.; Jones, A.; Tasaka, M.; Morita, M.; Friml, J. Gravity-induced PIN transcytosis for polarization of auxin fluxes in gravity-sensing root cells. Proc. Natl. Acad. Sci. USA 2010, 107, 22344–22349. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Lee, S.-H.; Cho, H.-T. Functional Identification of the phosphorylation sites of Arabidopsis PIN-FORMED3 for Its subcellular localization and biological role. Plant J. 2012, 71, 810–823. [Google Scholar] [CrossRef]
- Friml, J.; Yang, X.; Michniewicz, M.; Weijers, D.; Quint, A.; Tietz, O.; Benjamins, R.; Ouwerkerk, P.; Ljung, K.; Sandberg, G.; et al. A PINOID-Dependent Binary Switch in Apical-Basal PIN Polar Targeting Directs Auxin Efflux. Science 2004, 306, 862–865. [Google Scholar] [CrossRef]
- Michniewicz, M.; Zago, M.; Abas, L.; Weijers, D.; Schweighofer, A.; Meskiene, I.; Heisler, M.; Ohno, C.; Zhang, J.; Huang, F.; et al. Antagonistic regulation of PIN phosphorylation by PP2A and PINOID directs auxin flux. Cell 2007, 130, 1044–1056. [Google Scholar] [CrossRef]
- Kleine-Vehn, J.; Huang, F.; Naramoto, S.; Zhang, J.; Michniewicz, M.; Offringa, R.; Friml, J. PIN auxin efflux carrier activity is regulated by PINOID kinase-mediated recruitment into GNOM-independent trafficking in Arabidopsis. Plant Cell 2009, 21, 3839–3849. [Google Scholar] [CrossRef]
- Zourelidou, M.; Absmanner, B.; Weller, B.; Barbosa, I.; Willige, B.; Fastner, A.; Streit, V.; Port, S.; Colcombet, J.; de la Fuente van Bentem, S.; et al. Auxin efflux by PIN-FORMED proteins is activated by two different protein kinases, D6 PROTEIN KINASE and PINOID. eLife 2014, e02860. [Google Scholar] [CrossRef] [PubMed]
- Rakusová, H.; Gallego-Bartolomé, J.; Vanstraelen, M.; Robert, H.; Alabadí, D.; Blázquez, M.; Benková, E.; Friml, J. Polarization of PIN3-dependent auxin transport for hypocotyl gravitropic response in Arabidopsis thaliana. Plant J. 2011, 67, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Grones, P.; Abas, M.; Hajný, J.; Jones, A.; Waidmann, S.; Kleine-Vehn, J.; Friml, J. PID/WAG-mediated phosphorylation of the Arabidopsis PIN3 auxin transporter mediates polarity switches during gravitropism. Sci. Rep. 2018, 8, 10279. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.; Masson, P. ARL2, ARG1 and PIN3 define a gravity signal transduction pathway in root statocytes. Plant J. 2008, 53, 380–392. [Google Scholar] [CrossRef]
- Abas, L.; Benjamins, R.; Malenica, N.; Paciorek, T.; Wirniewska, J.; Moulinier-Anzola, J.; Sieberer, T.; Friml, J.; Luschnig, C. Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root gravitropism. Nat. Cell Biol. 2006, 8, 249–256. [Google Scholar] [CrossRef]
- Fasano, J.; Swanson, S.; Blancaflor, E.; Dowd, P.; Kao, T.; Gilroy, S. Changes in root cap pH are required for the gravity response of the Arabidopsis root. Plant Cell 2001, 13, 907–921. [Google Scholar] [CrossRef]
- Furutani, M.; Hirano, Y.; Nishimura, T.; Nakamura, M.; Taniguchi, M.; Suzuki, K.; Oshida, R.; Kondo, C.; Sun, S.; Kato, K.; et al. Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Shih, H.; DePew, C.; Miller, N.; Monshausen, G. The cyclic nucleotide-gated channel CNGC14 regulates root gravitropism in Arabidopsis thaliana. Curr. Biol. 2015, 25, 3119–3125. [Google Scholar] [CrossRef]
- Su, S.-H.; Gibbs, N.; Jancewicz, A.; Masson, P. Review: Molecular mechanisms of root gravitropism. Curr. Biol. 2017, 27, R964–R972. [Google Scholar] [CrossRef]
- Sedbrook, J.; Chen, R.; Masson, P. ARG1 (Altered Response to Gravity) encodes a DnaJ-like protein that potentially interacts with the cytoskeleton. Proc. Natl. Acad. Sci. USA 1999, 96, 1140–1145. [Google Scholar] [CrossRef]
- Chen, R.; Hilson, P.; Sedbrook, J.; Rosen, E.; Caspar, T.; Masson, P. The Arabidopsis thaliana AGRAVITROPIC 1 gene encodes a component of the polar-auxin-transport efflux carrier. Proc. Natl. Acad. Sci. USA 1998, 95, 15112–15117. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Rosen, E.; Boonsirichai, K.; Poff, K.; Masson, P. The ARG1-LIKE2 (ARL2) gene of Arabidopsis thaliana functions in a gravity signal transduction pathway that is genetically distinct from the PGM pathway. Plant Physiol. 2003, 133, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Boonsirichai, K.; Sedbrook, J.; Chen, R.; Gilroy, S.; Masson, P. ARG1 is a peripheral membrane protein that modulates gravity-induced cytoplasmic alkalinization and lateral auxin transport in plant statocytes. Plant Cell 2003, 15, 2612–2625. [Google Scholar] [CrossRef]
- Stanga, J.; Boonsirichai, K.; Sedbrook, J.; Otegui, M.; Masson, P. A role for the TOC complex in Arabidopsis root gravitropism. Plant Physiol. 2009, 149, 1896–1905. [Google Scholar] [CrossRef] [PubMed]
- Young, L.-S.; Harrison, B.; Murthy, N.; Moffatt, B.; Gilroy, S.; Masson, P. Adenosine kinase modulates root gravitropism and cap morphogenesis in Arabidopsis thaliana. Plant Physiol. 2006, 142, 564–573. [Google Scholar] [CrossRef]
- Abe, K.; Takahashi, H.; Suge, H. Graviresponding sites in shoots of normal and ‘lazy’ rice seedlings. Physiol. Plant 1994, 92, 371–374. [Google Scholar] [CrossRef]
- Abe, K.; Takahashi, H.; Suge, H. Localization of cells containing sedimented amyloplasts in the shoots of normal and lazy rice seedlings. Biol. Sci. Space 1994, 8, 221–225. [Google Scholar] [CrossRef]
- Godbole, R.; Takahashi, H.; Hertel, R. The lazy mutation in rice affects a step between statoliths and gravity-induced lateral auxin transport. Plant Biol. 1999, 1, 379–381. [Google Scholar] [CrossRef]
- Ge, L.; Chen, R. Negative gravitropism in plant roots. Nat. Plants 2016, 2, 16155. [Google Scholar] [CrossRef]
- Yoshihara, T.; Spalding, E. LAZY genes mediate the effects of gravity on auxin gradients and plant architecture. Plant Phyisol. 2017, 175, 959–969. [Google Scholar] [CrossRef]
- Taniguchi, M.; Furutani, M.; Nishimura, T.; Nakamura, M.; Fushita, T.; Iijima, K.; Baba, K.; Tanaka, H.; Toyota, M.; Tasaka, M.; et al. The Arabidopsis LAZY1 family plays a key role in gravity signaling within statocytes and in branch angle control of roots and shoots. Plant Cell 2017, 29, 1984–1999. [Google Scholar] [CrossRef] [PubMed]
- Guseman, J.; Webb, K.; Srinivasan, C.; Dardick, C. DRO1 influences root system architecture in Arabidopsis and Prunus species. Plant J. 2017, 89, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Nishimura, T.; Morita, M. Bridging the gap between amyloplasts and directional auxin transport in plant gravitropism. Curr. Opin. Plant Biol. 2019, 52, 54–60. [Google Scholar] [CrossRef]
- Li, P.; Wang, Y.; Qian, Q.; Fu, Z.; Wang, M.; Zeng, D.; Li, B.; Wang, X.; Li, J. LAZY1 controls rice shoot gravitropism through regulating polar auxin transport. Cell Res. 2007, 17, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Jiang, C.; Chen, X.; Zhang, T.; Ding, L.; Song, W.; Luo, H.; Lai, J.; Liu, R.; Chen, H.; et al. Maize LAZY1 mediates shoot gravitropism and inflorescence development through regulating auxin transport, auxin signaling and light response. Plant Physiol. 2013. [Google Scholar] [CrossRef]
- Yoshihara, T.; Spalding, E.; Iino, M. AtLAZY1 is a signaling component required for gravitropism of the Arabidopsis thaliana inflorescence. Plant J. 2013, 74, 267–279. [Google Scholar] [CrossRef]
- Li, Z.; Liang, Y.; Yuan, Y.; Wang, L.; Meng, X.; Xiong, G.; Zhou, J.; Cai, Y.; Han, N.; Hua, L.; et al. OsBRXL4 regulates shoot gravitropism and rice tiller angle through affecting LAZY1 nuclear localization. Mol. Plant 2019, 12, 1143–1156. [Google Scholar] [CrossRef]
- Ashraf, A.; Rehman, O.; Muzammil, S.; Léon, J.; Naz, A.; Rasool, F.; Ali, G.; Zafar, Y.; Khan, M. Evolution of Deeper Rooting 1-like homeologs in wheat entails the C-terminus mutations as well as gain and loss of auxin-response elements. PLoS ONE 2019, 14, e0214145. [Google Scholar] [CrossRef]
- Yoshihara, T.; Spalding, E. Switching the direction of stem gravitropism by altering two amino acids in AtLAZY1. Plant Phyisol. 2020, 182, 1039–1051. [Google Scholar] [CrossRef]
- Pereira-Leal, J.; Seabra, M. Evolution of the Rab family of small GTP-binding proteins. J. Mol. Biol. 2001, 313, 889–891. [Google Scholar] [CrossRef]
- Mayers, J.; Hu, T.; Wang, C.; Cárdenas, J.; Tan, Y.; Pan, J.; Bednarek, S. SCD1 and SCD2 form a complex that functions with the exocyst and RabE1 in exocytosis and cytokinesis. Plant Cell 2017, 29, 2610–2665. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Wen, Q.; Yu, R.; Han, X.; Deng, X.; Chen, H. Light modulates the gravitropic responses through organ-specific PIFs and HY5 regulation of LAZY4 expression in Arabidopsis. Proc. Nat. Acad. Sci. USA 2020, 117, 18840–18848. [Google Scholar] [CrossRef] [PubMed]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Barbosa, I.; Zourelidou, M.; Willige, B.; Weller, B.; Schwechheimer, C. D6 PROTEIN KINASE activates auxin transport-dependent growth and PIN-FORMED phosphorylation at the plasma membrane. Dev. Cell 2014, 29, 674–685. [Google Scholar] [CrossRef]
- Dhonukshe, P.; Huang, F.; Galvan-Ampudia, C.; Mähönen, A.; Kleine-Vehn, J.; Xu, J.; Quint, A.; Prasad, K.; Friml, J.; Scheres, B.; et al. Plasma membrane-bound AGC3 kinases phosphorylate PIN auxin carriers at TPRXS(N/S) motifs to direct apical PIN recycling. Development 2010, 137, 3245–3255. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Nagawa, S.; Chen, J.; Cao, L.; Chen, X.; Xu, T.; Li, H.; Dhonukshe, P.; Yamamuro, C.; Friml, J.; et al. A ROP GTPase-dependent auxin signaling pathway regulates the subcellular distribution of PIN2 in Arabidopsis roots. Curr. Biol. 2012, 22, 1319–1325. [Google Scholar] [CrossRef]
- Li, G.; Xue, H. Arabidopsis PLD{zeta}2 regulates vesicle trafficking and is required for auxin response. Plant Cell 2007, 19, 281–295. [Google Scholar] [CrossRef]
- Whitford, R.; Fernandez, A.; Tejos, R.; Pérez, A.; Kleine-Vehn, J.; Vanneste, S.; Drozdzecki, A.; Leitner, J.; Abas, L.; Aerts, M.; et al. GOLVEN secretory peptides regulate auxin carrier turnover during plant gravitropic responses. Dev. Cell 2012, 22, 678–685. [Google Scholar] [CrossRef]
- Retzer, K.; Akhmanova, M.; Konstantinova, N.; Malínská, K.; Leitner, J.; Petrásek, J.; Luschnig, C. Brassinosteroid signaling delimits root gravitropism via sorting of the Arabidopsis PIN2 auxin transporter. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Dharmasiri, S.; Swarup, R.; Mockaitis, K.; Dharmasiri, N.; Singh, S.; Kowalchyk, M.; Marchant, A.; Mills, S.; Sandberg, G.; Bennett, M.; et al. AXR4 is required for localization of the auxin influx facilitator AUX1. Science 2006, 312, 1218–1220. [Google Scholar] [CrossRef]
- Swarup, R.; Kramer, E.; Perry, P.; Knox, K.; Leyser, H.; Haseloff, J.; Beemster, G.; Bhalerao, R.; Bennett, M. Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal. Nat. Cell Biol. 2005, 7, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Murphy, A. Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J. 2009, 59, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.; Miller, N.; Splitt, B.; Wu, G.; Spalding, E. Separating the roles of acropetal and basipetal auxin transport on gravitropism with mutations in two Arabidopsis multidrug resistance-like ABC transporter genes. Plant Cell 2007, 19, 1838–1850. [Google Scholar] [CrossRef] [PubMed]
- Mullen, J.; Ishikawa, H.; Evans, M. Analysis of changes in relative elemental growth rate patterns in the elongation zone of Arabidopsis roots upon gravistimulation. Planta 1998, 206, 598–603. [Google Scholar] [CrossRef]
- Dindas, J.; Scherzer, S.; Roelfsema, M.; von Meyer, K.; Müller, H.; Al-Rasheid, K.; Palme, K.; Dietrich, P.; Becker, D.; Bennett, M.; et al. AUX1-mediated root hair auxin influx governs SCFTIR1/AFB-type Ca2+ signaling. Nat. Commun. 2018, 9, 1174. [Google Scholar] [CrossRef]
- Terrile, M.; París, R.; Calderón-Villalobos, L.; Iglesias, M.; Lamattina, L.; Estelle, M.; Casalongué, C. Nitric oxide influences auxin signaling through S-nitrosylation of the Arabidopsis TRANSPORT INHIBITOR RESPONSE 1 auxin receptor. Plant J. 2012, 70, 492–500. [Google Scholar] [CrossRef]
- Krieger, G.; Shkolnik, D.; Miller, G.; Fromm, H. Reactive oxygen species tune root tropic responses. Plant Phyisol. 2016, 172, 1209–1220. [Google Scholar] [CrossRef]
- Cosgrove, D. Loosening of Plant Cell Walls by Expansins. Nature 2000, 407, 321–326. [Google Scholar] [CrossRef]
- Bai, H.; Murali, B.; Barber, K.; Wolverton, C. Low phosphate alters lateral root setpoint angle and gravitropism. Am. J. Bot. 2013, 100, 175–182. [Google Scholar] [CrossRef]
- Kiss, J.; Miller, K.; Ogden, L.; Roth, K. Phototropism and gravitropism in lateral roots of Arabidopsis. Plant Cell Physiol. 2002, 43, 35–43. [Google Scholar] [CrossRef]
- Guyomarc’h, S.; Léran, S.; Auzon-Cape, M.; Perrine-Walker, F.; Lucas, M.; Laplaze, L. Early development and gravitropic response of lateral roots in Arabidopsis thaliana. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Rosquete, M.; von Wangenheim, D.; Marhavý, P.; Barbez, E.; Stelzer, E.; Benková, E.; Maizel, A.; Kleine-Vehn, J. An auxin transport mechanism restricts positive orthogravitropism in lateral roots. Curr. Biol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhry, S.; Del Bianco, M.; Kieffer, M.; Kepinski, S. Auxin controls gravitropic setpoint angle in higher plant lateral branches. Curr. Biol. 2013, 23, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Chen, R. Negative gravitropic response of roots directs auxin flow to control root gravitropism. Plant Cell Environ. 2019, 42, 2372–2383. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, N.; Kanbe, Y.; Nakamura, M.; Mori, A.; Morita, M. Gravity-sensing tissues for gravitropism are required for “anti-gravitropic” phenotypes of lzy multiple mutants in Arabidopsis. Plants 2020, 9, 615. [Google Scholar] [CrossRef] [PubMed]
- Waidmann, S.; Ruiz Rosquete, M.; Schöller, M.; Sarkel, E.; Lindner, H.; LaRue, T.; Petřík, I.; Dünser, K.; Martopawiro, S.; Sasidharan, R.; et al. Cytokinin functions as an asymmetric and anti-gravitropic signal in lateral roots. Nat. Commun. 2019, 10, 3540. [Google Scholar] [CrossRef]
- Yu, B.; Lin, Z.; Li, H.; Li, X.; Li, J.; Wang, Y.; Zhang, X.; Zhu, Z.; Zhai, W.; Wang, X.; et al. TAC1, a major quantitative trait locus controlling tiller angle in rice. Plant J. 2007, 52, 891–898. [Google Scholar] [CrossRef]
- Dardick, C.; Callahan, A.; Horn, R.; Ruiz, K.; Zhebentyayeva, T.; Hollender, C.; Whitaker, M.; Abbott, A.; Scorza, R. PpeTAC1 promotes the horizontal growth of branches in peach trees and is a member of a functionally conserved gene family found in diverse plants species. Plant J. 2013, 75, 618–630. [Google Scholar] [CrossRef]
- Waite, J.; Dardick, C. TILLER ANGLE CONTROL 1 modulates plant architecture in response to photosynthetic signals. J. Exp. Bot. 2018, 69, 4935–4944. [Google Scholar] [CrossRef]
- Ruiz Rosquete, M.; Waidmann, S.; Kleine-Vehn, J. PIN7 auxin carrier has a preferential role in terminating radial root expansion in Arabidopsis thaliana. Int. J. Mol. Sci. 2018, 19, 1238. [Google Scholar] [CrossRef]
- Takahashi, N.; Yamazaki, Y.; Kobayashi, A.; Higashitani, A.; Takahashi, H. Hydrotropism interacts with gravitropism by degrading amyloplasts in seedling roots of Arabidopsis and radish. Plant Physiol. 2003, 132, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Shin, J.; Lee, S.; Kweon, H.; Maloof, J.; Choi, G. Phytochromes inhibit hypocotyl negative gravitropism by regulating the development of endodermal amyloplasts through phytochrome-interacting factors. Proc. Natl. Acad. Sci. USA 2011, 108, 1729–1744. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Jeong, J.; Kim, J.; Lee, N.; Eon, M.; Sangil, K.; Sun, L.; Kim, C.; Choi, G. PIF1 regulates plastid development by repressing photosynthetic genes in the endodermis. Mol. Plant 2016, 9, 1415–1427. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Kim, H.-S.; Park, J.; Cho, H.; Jeon, J. PIN-mediated polar auxin transport facilitates root–obstacle avoidance. New Phytol. 2020, 225, 1285–1296. [Google Scholar] [CrossRef]
- Porterfield, D.; Musgrave, M. The tropic response of plant roots to oxygen: Oxytropism in Pisum sativum L. Planta 1998, 20, 1–6. [Google Scholar] [CrossRef]
- Kiss, J.; Correll, M.; Mullen, J.; Hangarter, R.; Edelmann, R. Root phototropism: How light and gravity interact in shaping plant form. Gravit. Space Biol. Bull. 2003, 16, 55–60. [Google Scholar] [PubMed]
- Choi, W.; Barker, R.; Kim, S.; Swanson, S.; Gilroy, S. Variation in the transcriptome of different ecotypes of Arabidopsis thaliana reveals signatures of oxidative stress in plant responses to spaceflight. Am. J. Bot. 2019, 106, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Sng, N.; Zupanska, A.; Krishnamurthy, A.; Schultz, E.; Ferl, R. Genetic dissection of the Arabidopsis spaceflight transcriptome: Are some responses dispensable for the physiological adaptation of plants to spaceflight? PLoS ONE 2017, 12, e0180186. [Google Scholar] [CrossRef]
- Kwon, T.; Sparks, J.; Nakashima, J.; Allen, S.; Tang, Y.; Blancaflor, E. Transcriptional response of Arabidopsis seedlings during spaceflight reveals peroxidase and cell wall remodeling genes associated with root hair development. Am. J. Bot. 2015, 102, 21–35. [Google Scholar] [CrossRef]
- Basu, P.; Kruse, C.; Luesse, D.; Wyatt, S. Growth in spaceflight hardware results in alterations to the transcriptome and proteome. Life Sci. Space Res. 2017, 15, 88–96. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, S.-H.; Keith, M.A.; Masson, P.H. Gravity Signaling in Flowering Plant Roots. Plants 2020, 9, 1290. https://doi.org/10.3390/plants9101290
Su S-H, Keith MA, Masson PH. Gravity Signaling in Flowering Plant Roots. Plants. 2020; 9(10):1290. https://doi.org/10.3390/plants9101290
Chicago/Turabian StyleSu, Shih-Heng, Marie A. Keith, and Patrick H. Masson. 2020. "Gravity Signaling in Flowering Plant Roots" Plants 9, no. 10: 1290. https://doi.org/10.3390/plants9101290
APA StyleSu, S.-H., Keith, M. A., & Masson, P. H. (2020). Gravity Signaling in Flowering Plant Roots. Plants, 9(10), 1290. https://doi.org/10.3390/plants9101290