Ecosystem Services, Physiology, and Biofuels Recalcitrance of Poplars Grown for Landfill Phytoremediation

 ,
,  ,
, 
Abstract
1. Introduction
2. Results
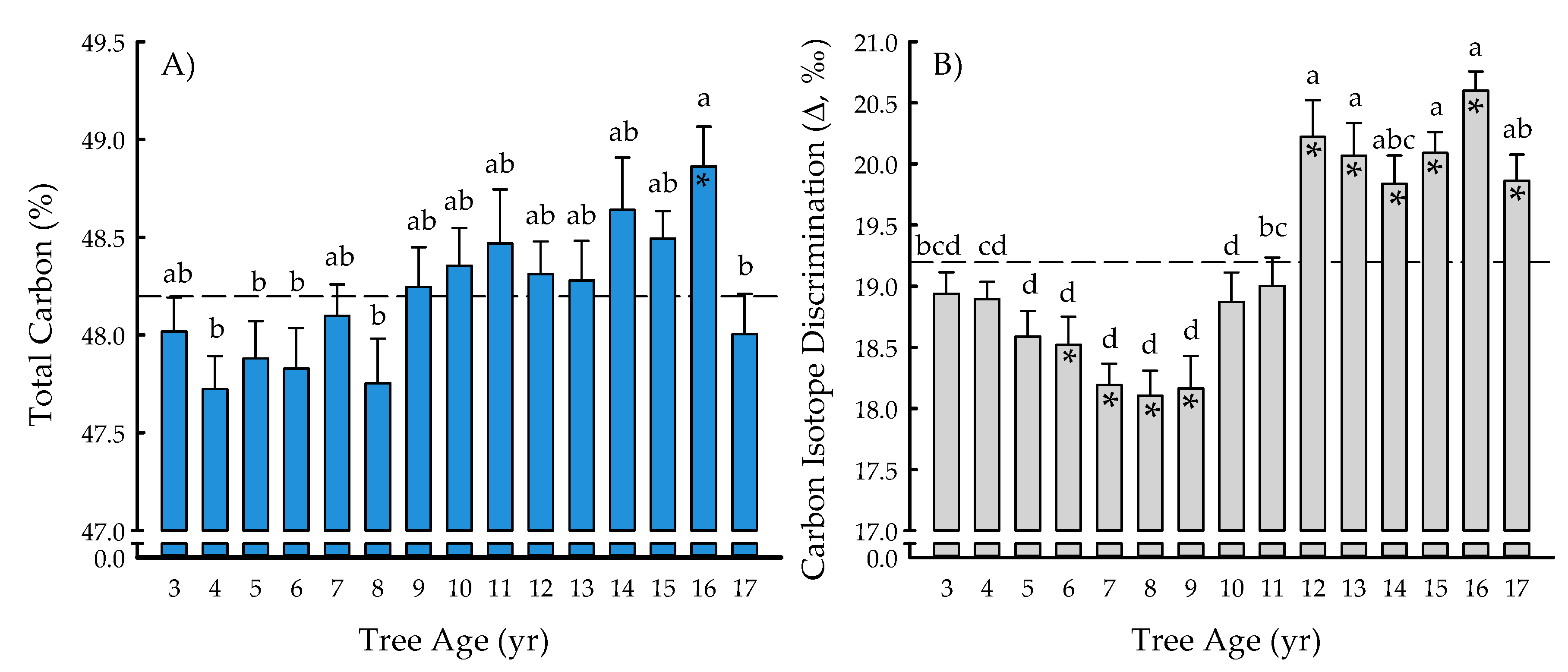
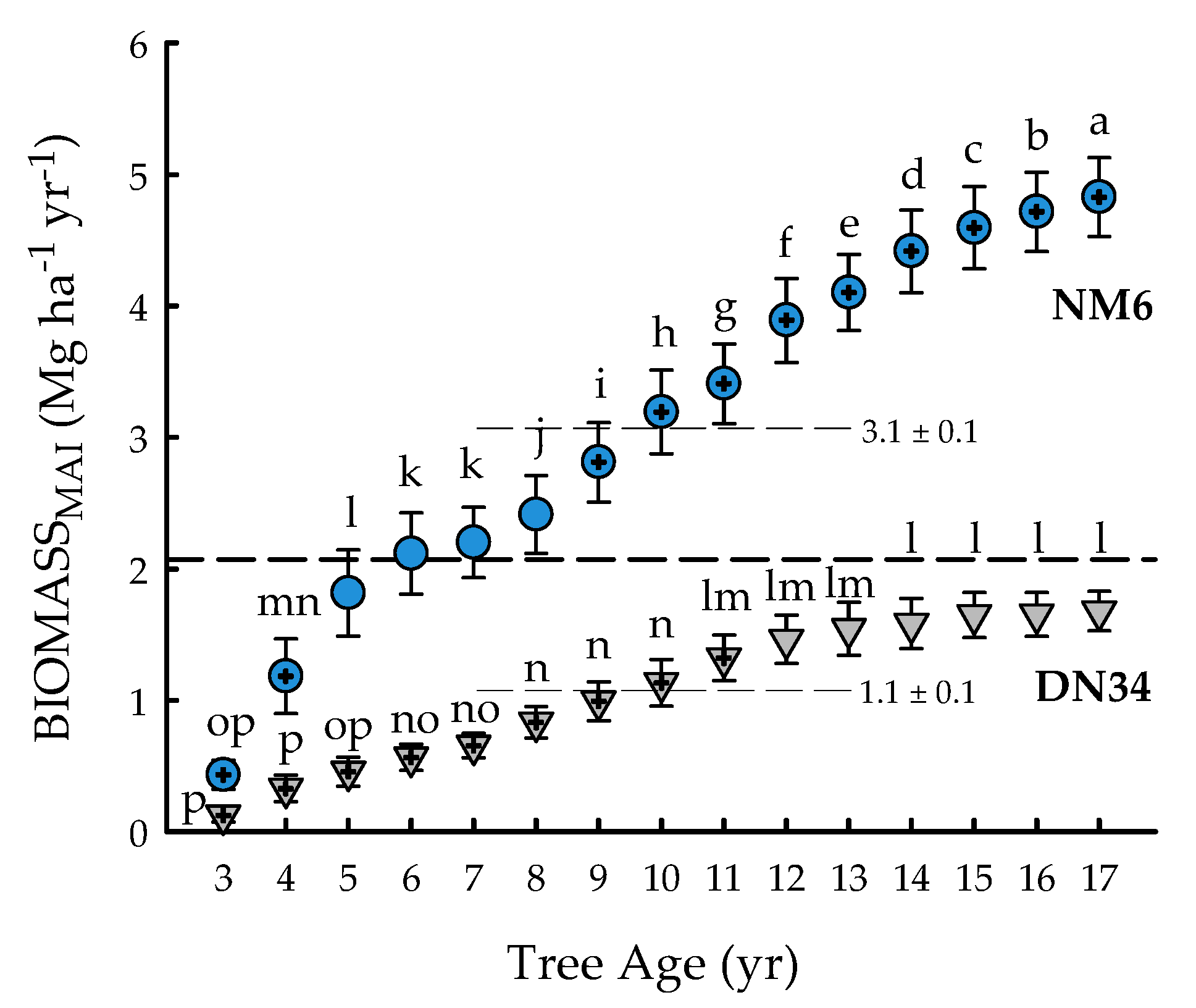
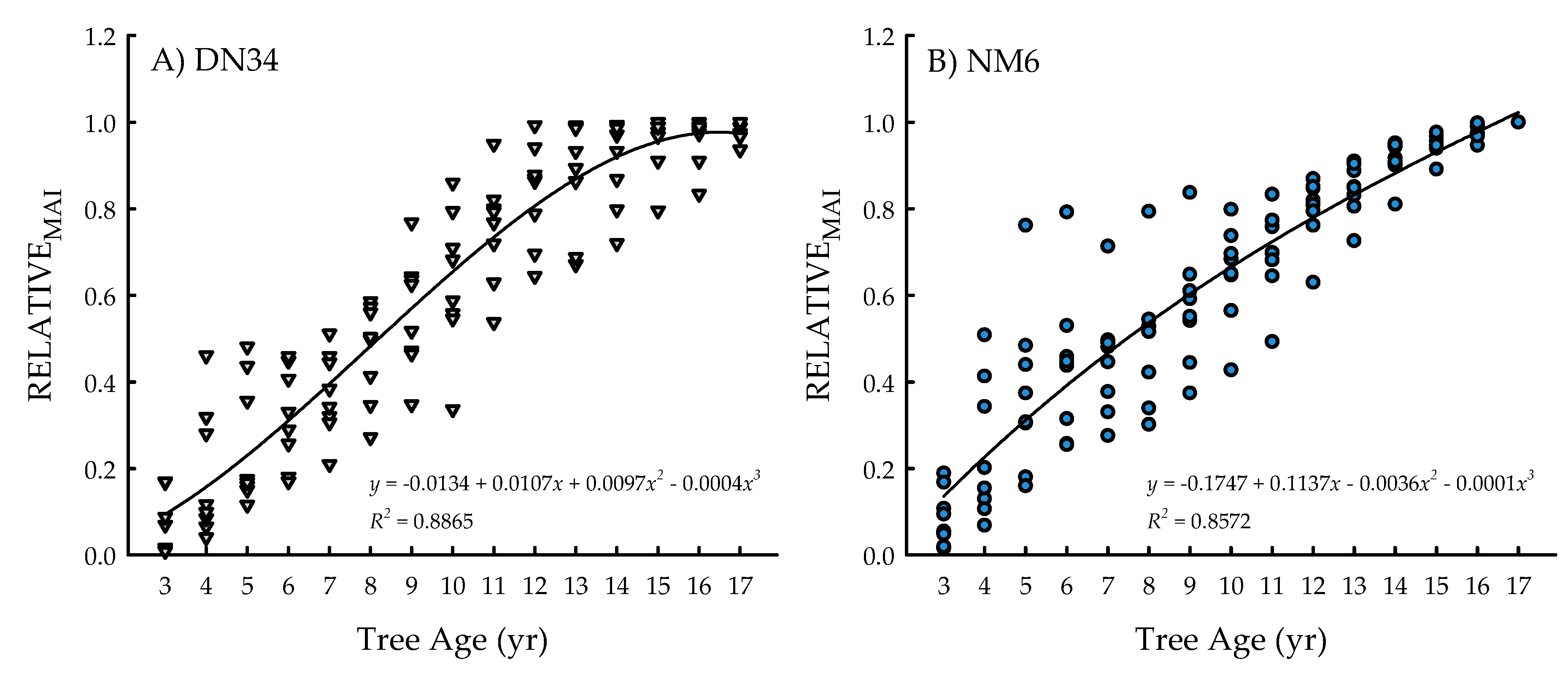
2.1. Ecosystem Services and Physiology
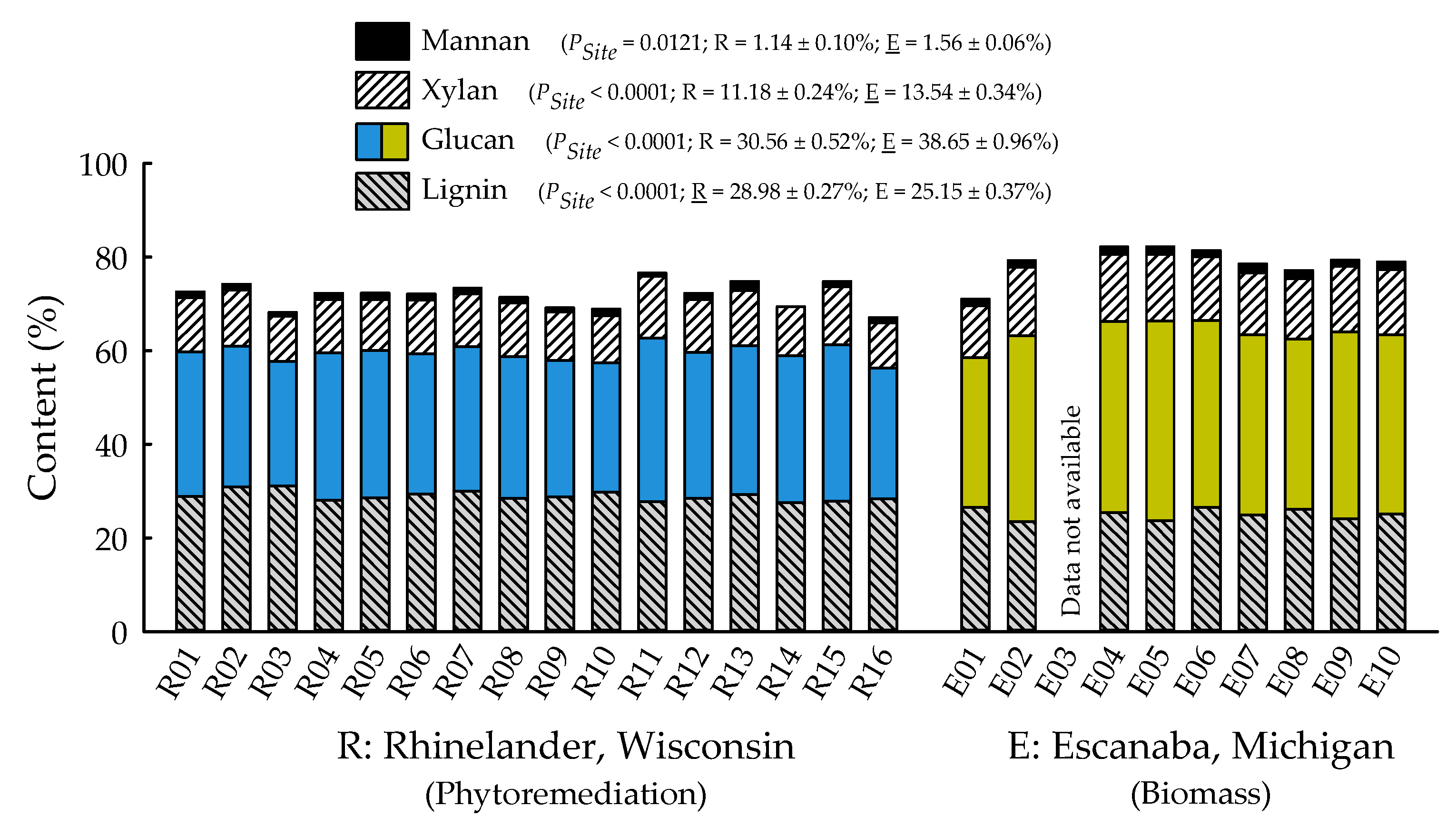
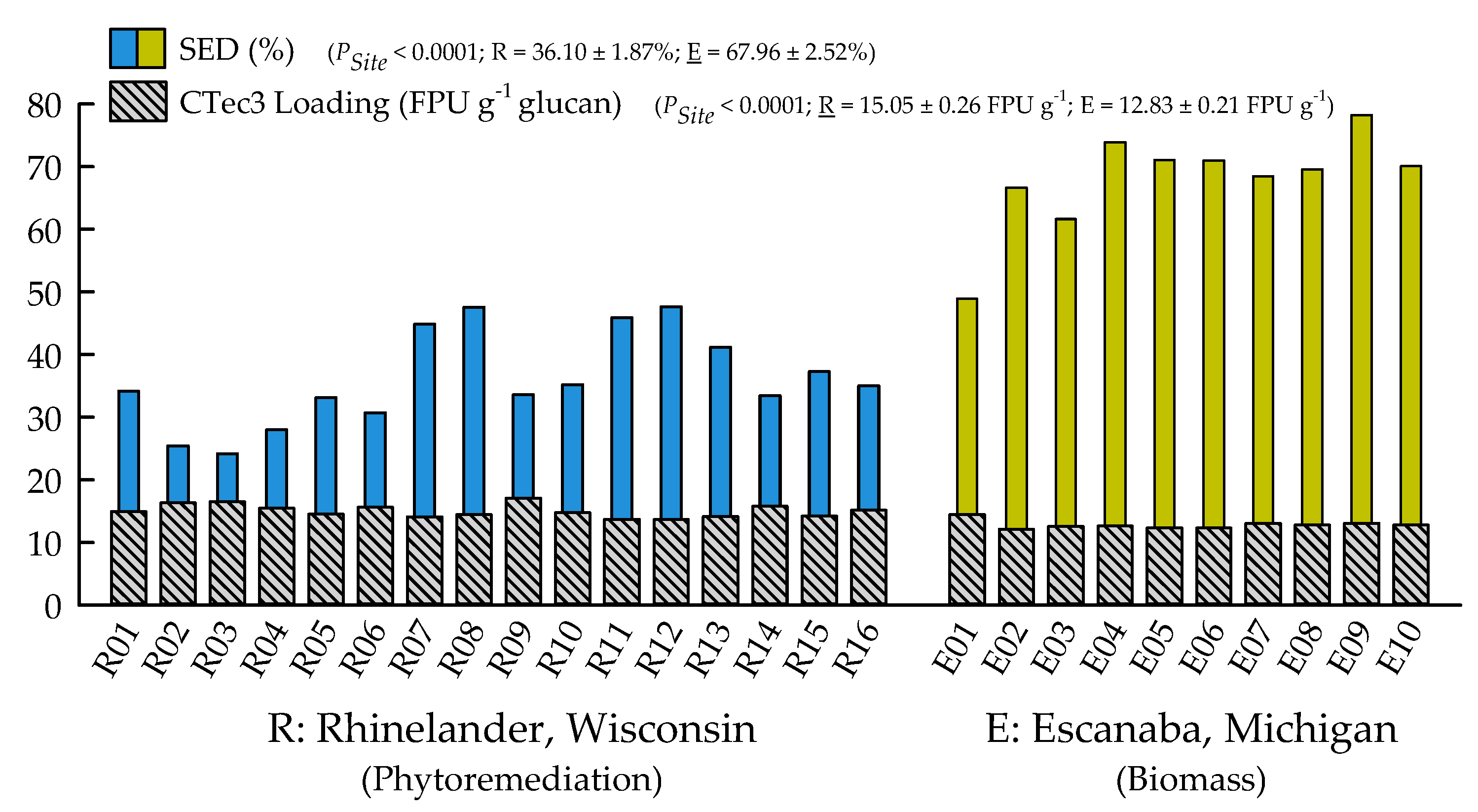
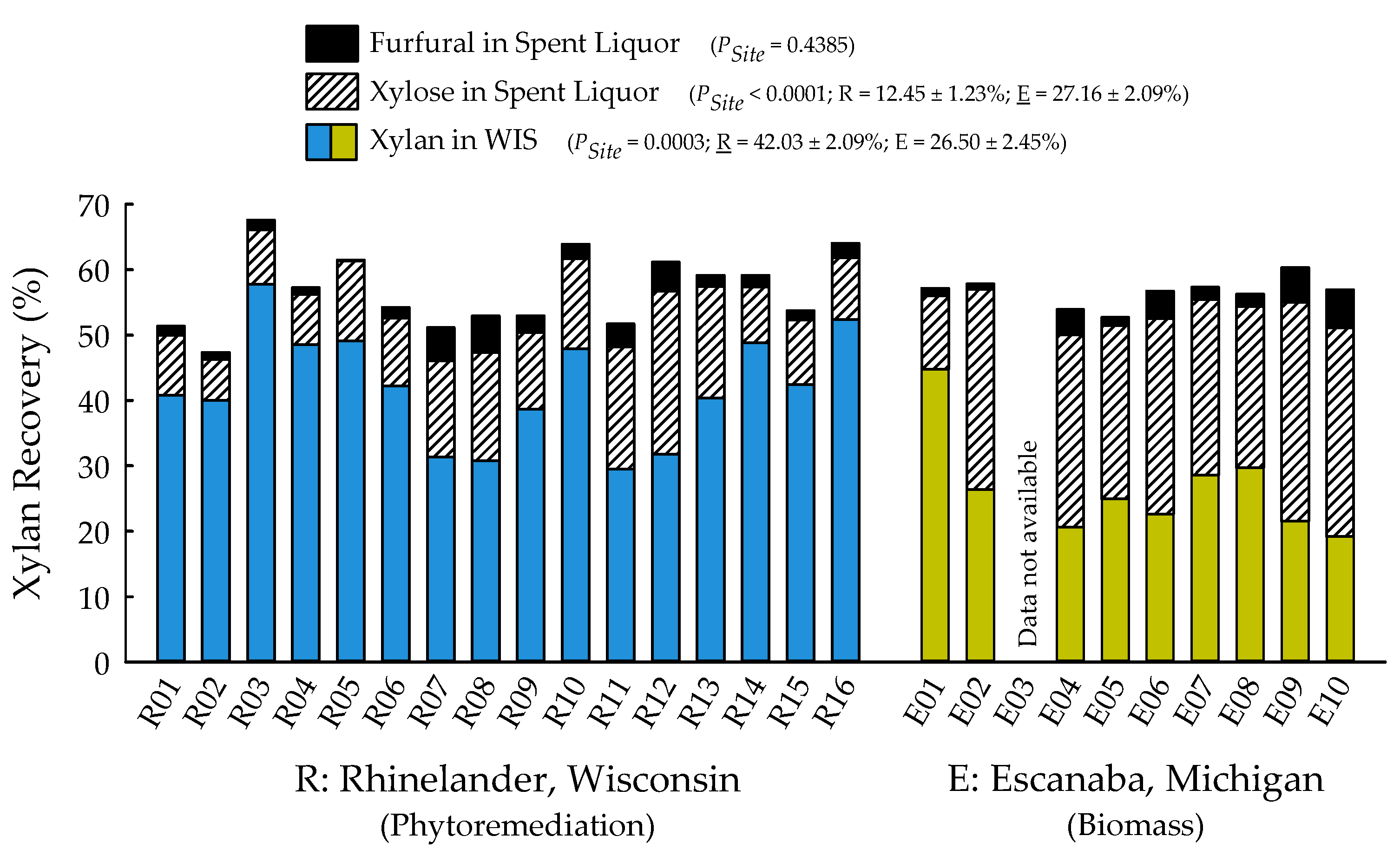
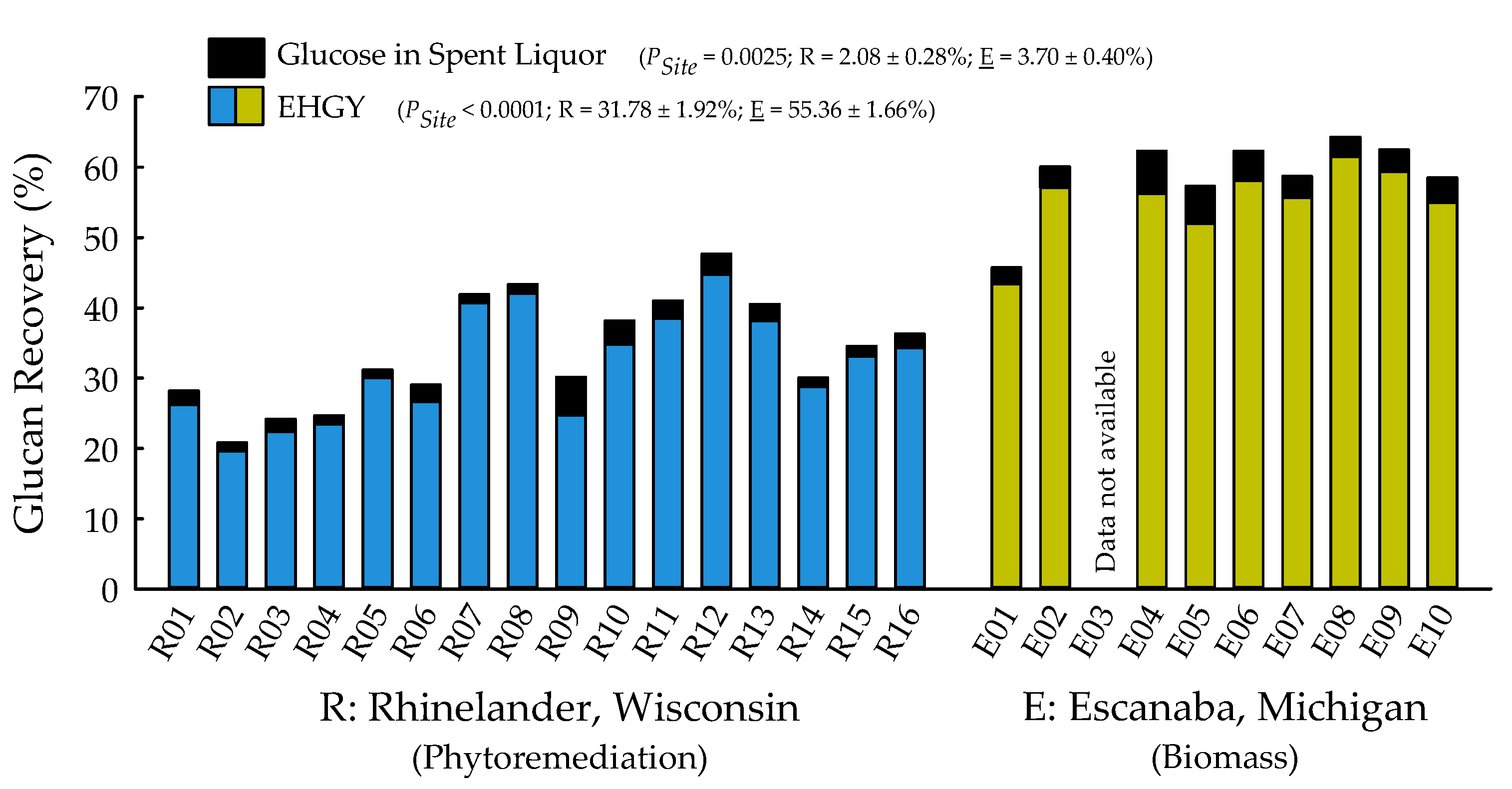
2.2. Biofuels Recalcitrance
3. Discussion
4. Materials and Methods
4.1. Clone and Site Selection
4.2. Field Sampling
4.3. Ring Width Measurements and Laboratory Sampling
4.4. Total Carbon and Wood Composition
4.5. Carbon Isotope (δ13C) Analyses and Discrimination (Δ) Calculations
4.6. Annual Biomass Estimates
4.7. Sulfite Pretreatment to Overcome Recalcitrance of Lignocelluloses (SPORL) Pretreatment
4.8. Enzymatic Hydrolysis
4.9. Chemical Composition Analyses
4.10. Experimental Design and Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carle, J. Trends & Perspectives in Poplar & Willow Cultivation: Global Synthesis. In Proceedings of the 25th Session of the FAO International Poplar Commission, Berlin, Germany, 12–16 September 2016. Available online: http://ipc25berlin2016.com/fileadmin/allgemein/pdf/veranstaltungen/IPC25_2016/Plenary_4/Carle_JB_Global_Synthesis_Keynote_Plenary_16_Sept_2016.pdf (accessed on 6 March 2020).
- Isebrands, J.G.; Aronsson, P.; Carlson, M.; Ceulemans, R.; Coleman, M.; Dickinson, N.; Dimitriou, J.; Doty, S.; Gardiner, E.; Heinsoo, K.; et al. Environmental applications of poplars and willows. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; Food and Agriculture Organization (FAO) of the United Nations: Rome, Italy; CAB International, Inc.: Boston, MA, USA, 2014; Chapter 6; pp. 258–336. ISBN 978-1-78064-108-9. [Google Scholar]
- Vance, E.D.; Maguire, D.A.; Zalesny, R.S., Jr. Research strategies for increasing productivity of intensively managed forest plantations. J. For. 2010, 108, 183–192. [Google Scholar] [CrossRef]
- Pilipović, A.; Orlović, S.; Rončević, S.; Nikolić, N.; Župunski, M.; Spasojević, J. Results of selection of poplars and willows for water and sediment phytoremediation. Agric. For. 2015, 61, 205–211. [Google Scholar] [CrossRef]
- Licht, L.A.; Isebrands, J.G. Linking phytoremediated pollutant removal to biomass economic opportunities. Biomass Bioenergy 2005, 28, 203–218. [Google Scholar] [CrossRef]
- Pilipović, A.; Zalesny, R.S., Jr.; Orlović, S.; Drekić, M.; Pekeć, S.; Katanić, M.; Poljaković-Pajnik, L. Growth and physiological responses of three poplar clones grown on soils artificially contaminated with heavy metals, diesel fuel, and herbicides. Int. J. Phytoremed. 2020, 22, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Headlee, W.L.; Gopalakrishnan, G.; Bauer, E.O.; Hall, R.B.; Hazel, D.W.; Isebrands, J.G.; Licht, L.A.; Negri, M.C.; Guthrie-Nichols, E.; et al. Ecosystem services of poplar at long-term phytoremediation sites in the Midwest and Southeast, United States. WIREs Energy Environ. 2019, e349. [Google Scholar] [CrossRef]
- Baldantoni, D.; Cicatelli, A.; Bellino, A.; Castiglione, S. Different behaviours in phytoremediation capacity of two heavy metal tolerant poplar clones in relation to iron and other trace elements. J. Environ. Manag. 2014, 146, 94–99. [Google Scholar] [CrossRef]
- Laureysens, I.; Blust, R.; De Temmerman, L.; Lemmens, C.; Ceulemans, R. Clonal variation in heavy metal accumulation and biomass production in a poplar coppice culture: I. seasonal variation in leaf, wood and bark concentrations. Environ. Pollut. 2004, 131, 485–494. [Google Scholar] [CrossRef]
- Pilipović, A.; Zalesny, R.S., Jr.; Rončević, S.; Nikolić, N.; Orlović, S.; Beljin, J.; Katanić, M. Growth, physiology, and phytoextraction potential of poplar and willow established in soils amended with heavy-metal contaminated, dredged river sediments. J. Environ. Manag. 2019, 239, 352–365. [Google Scholar] [CrossRef]
- Burken, J.G.; Schnoor, J.L. Distribution and volatilization of organic compounds following uptake by hybrid poplar trees. Int. J. Phytoremed. 1999, 1, 139–151. [Google Scholar] [CrossRef]
- Cook, R.L.; Landmeyer, J.E.; Atkinson, B.; Messier, J.-P.; Guthrie Nichols, E. Successful establishment of a phytoremediation system at a petroleum hydrocarbon contaminated shallow aquifer: Trends, trials, and tribulations. Int. J. Phytoremed. 2010, 12, 716–732. [Google Scholar] [CrossRef]
- Doty, S.L.; Freeman, J.L.; Cohu, C.M.; Burken, J.G.; Firrincieli, A.; Simon, A.; Khan, Z.; Isebrands, J.G.; Lukas, J.; Blaylock, M.J. Enhanced degradation of TCE on a superfund site using endophyte-assisted poplar tree phytoremediation. Environ. Sci. Technol. 2017, 51, 10050–10058. [Google Scholar] [CrossRef]
- Abichou, T.; Musagasa, J.; Yuan, L.; Chanton, J.; Tawfiq, K.; Rockwood, D.; Licht, L. Field performance of alternative landfill covers vegetated with cottonwood and eucalyptus trees. Int. J. Phytoremed. 2012, 14 (Suppl. 1), 47–60. [Google Scholar] [CrossRef]
- Zalesny, J.A.; Zalesny, R.S., Jr.; Wiese, A.H.; Hall, R.B. Choosing tree genotypes for phytoremediation of landfill leachate using phyto-recurrent selection. Int. J. Phytoremed. 2007, 9, 513–530. [Google Scholar] [CrossRef] [PubMed]
- Zupančič-Justin, M.; Pajk, N.; Zupanc, V.; Zupančič, M. Phytoremediation of landfill leachate and compost wastewater by irrigation of Populus and Salix: Biomass and growth response. Waste Manag. 2010, 30, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Riemenschneider, D.E.; Berguson, W.E.; Dickmann, D.I.; Hall, R.B.; Isebrands, J.G.; Mohn, C.A.; Stanosz, G.R.; Tuskan, G.A. Poplar breeding and testing strategies in the north-central U.S.: Demonstration of potential yield and consideration of future research needs. For. Chron. 2001, 77, 245–253. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Hall, R.B.; Zalesny, J.A.; Berguson, W.E.; McMahon, B.G.; Stanosz, G.R. Biomass and genotype × environment interactions of Populus energy crops in the Midwestern United States. BioEnergy Res. 2009, 2, 106–122. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment (MEA). Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005; Available online: https://www.millenniumassessment.org/documents/document.356.aspx.pdf (accessed on 1 September 2020).
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; Headlee, W.L.; Bañuelos, G.S.; Hass, A. Ecosystem services of woody crop production systems. BioEnergy Res. 2016, 9, 465–491. [Google Scholar] [CrossRef]
- Burges, A.; Alkorta, I.; Epelde, L.; Garbisu, C. From phytoremediation of soil contaminants to phytomanagement of ecosystem services in metal contaminated sites. Int. J. Phytoremed. 2018, 20, 384–397. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Richardson, J. Poplars and Willows: Trees for Society and the Environment; Food and Agriculture Organization (FAO) of the United Nations: Rome, Italy; CAB International, Inc.: Boston, MA, USA, 2014; ISBN 978-1-78064-108-9. [Google Scholar]
- Zalesny, R.S., Jr.; Headlee, W.L. Developing woody crops for the enhancement of ecosystem services under changing climates in the North Central United States. J. For. Exp. Sci. 2015, 31, 78–90. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Bauer, E.O. Selecting and utilizing Populus and Salix for landfill covers: Implications for leachate irrigation. Int. J. Phytoremed. 2007, 9, 497–511. [Google Scholar] [CrossRef]
- Allen, S.; Hall, R.; Rosier, P. Transpiration by two poplar varieties grown as coppice for biomass production. Tree Physiol. 1999, 19, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Vose, J.; Swank, W.; Harvey, G.; Clinton, B.; Sobek, C. Leaf water relations and sapflow in eastern cottonwood (Populus deltoides Bartr.) trees planted for phytoremediation of a groundwater pollutant. Int. J. Phytoremed. 2000, 2, 53–73. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Wiese, A.H.; Bauer, E.O.; Riemenschneider, D.E. Sapflow of hybrid poplar (Populus nigra L. × P. maximowiczii A. Henry ‘NM6′) during phytoremediation of landfill leachate. Biomass Bioenergy 2006, 30, 784–793. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Berndes, G.; Dimitriou, I.; Fritsche, U.; Miller, C.; Eisenbies, M.; Ghezehei, S.; Hazel, D.; Headlee, W.L.; Mola-Yudego, B.; et al. Positive water linkages of producing short rotation poplars and willows for bioenergy and phytotechnologies. WIREs Energy Environ. 2019, 8, e345. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Austral J. Plant Physiol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Hubick, K.T.; Condon, A.G.; Richards, R.A. Carbon isotope fractionation and plant water-use efficiency. In Stable Isotopes in Ecological Research; Rundel, P.W., Ehleringer, J.R., Nagy, K.A., Eds.; Springer: New York, NY, USA, 1989; pp. 21–40. [Google Scholar] [CrossRef]
- Radwanski, D.; Gallagher, F.; Vanderlinden, D.W.; Schafer, K.V.R. Photosynthesis and aboveground carbon allocation of two co-occurring poplar species in an urban brownfield. Environ. Pollut. 2017, 223, 497–506. [Google Scholar] [CrossRef]
- Martin, P.J.; Stephens, W. Willow growth in response to nutrients and moisture on a clay landfill cap soil. II: Water use. Biores. Technol. 2006, 97, 449–458. [Google Scholar] [CrossRef][Green Version]
- Schifman, L.A.; Stella, J.C.; Volk, T.A.; Teece, M.A. Carbon isotope variation in shrub willow (Salix spp.) ring-wood as an indicator of long-term water status, growth and survival. Biomass Bioenergy 2012, 36, 316–326. [Google Scholar] [CrossRef]
- Shrive, S.C.; McBride, R.A.; Gillespe, T.J. Physiological and spectral responses of sugar maple (Acer saccharinum Marsh.) to MSW leachate spray irrigation. Waste Manag. Res. 1990, 8, 3–19. [Google Scholar] [CrossRef]
- Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef]
- Bert, V.; Neub, S.; Zdanevitch, I.; Friesl-Hanl, W.; Collet, S.; Gaucher, R.; Puschenreiter, M.; Müller, I.; Kumpiene, J. How to manage plant biomass originated from phytotechnologies? Gathering perceptions from end-users. Int. J. Phytoremed. 2017, 19, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.-F.; Coleman, M.D.; Gesch, R.; Jaradat, A.; Mitchell, R.; Reicosky, D.; Wilhelm, W.W. Biomass-bioenergy crops in the United States: A changing paradigm. Am. J. Plant Sci. Biotechnol. 2007, 1, 1–28. [Google Scholar]
- Zhu, J.Y.; Pan, X.; Zalesny, R.S., Jr. Pretreatment of woody biomass for biofuel production: Energy efficiency, technologies, and recalcitrance. Appl. Microbiol. Biotechnol. 2010, 87, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gu, F.; Zhu, J.Y.; Zalesny, R.S., Jr. Using a combined hydrolysis factor to optimize high titer ethanol production from sulfite-pretreated poplar without detoxification. Bioresour. Technol. 2015, 186, 223–231. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, L.; Gleisner, R.; Zhu, J.Y. Co-producing bioethanol and furfural from poplar using acid hydrotropic fractionation (AHF). Fuel 2019, 254, 115572. [Google Scholar] [CrossRef]
- Wang, Z.J.; Zhu, J.Y.; Zalesny, R.S., Jr.; Chen, K.F. Ethanol production potential from poplar wood through enzymatic saccharification and fermentation. Fuel 2012, 95, 606–614. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Stanturf, J.A.; Gardiner, E.S.; Bañuelos, G.S.; Hallett, R.A.; Hass, A.; Stange, C.M.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; et al. Environmental technologies of woody crop production systems. BioEnergy Res. 2016, 9, 492–506. [Google Scholar] [CrossRef]
- Cunningham, S.D.; Ow, D.W. Promises and prospects of phytoremediation. Plant Physiol. 1996, 110, 715–719. [Google Scholar] [CrossRef]
- Mirck, J.; Isebrands, J.G.; Verwijst, T.; Ledin, S. Development of short-rotation willow coppice systems for environmental purposes in Sweden. Biomass Bioenergy 2005, 28, 219–228. [Google Scholar] [CrossRef]
- Gopalakrishnan, G.; Negri, M.C.; Wang, M.; Wu, M.; Snyder, S.W.; Lafreniere, L. Biofuels, land, and water: A systems approach to sustainability. Environ. Sci. Technol. 2009, 43, 6094–6100. [Google Scholar] [CrossRef][Green Version]
- Miller, R.O. Growth variation among hybrid poplar varieties in Michigan, USA and the implications for commercial biomass production. BioEnergy Res. 2018, 11, 816–825. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Bauer, E.O.; Hall, R.B.; Zalesny, J.A.; Kunzman, J.; Rog, C.J.; Riemenschneider, D.E. Clonal variation in survival and growth of hybrid poplar and willow in an in situ trial on soils heavily contaminated with petroleum hydrocarbons. Int. J. Phytoremed. 2005, 7, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Zalesny, R.S., Jr.; Bauer, E.O. Genotypic variability and stability of poplars and willows grown on nitrate-contaminated soils. Int. J. Phytoremed. 2019, 21, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; LeBoldus, J.M. The infection biology of Sphaerulina musiva: Clues to understanding a forest pathogen. PLoS ONE 2014, 9, e103477. [Google Scholar] [CrossRef]
- Gebrekirstos, A.; van Noordwijk, M.; Neufeldt, H.; Mitlohner, R. Relationships of stable carbon isotopes, plant water potential and growth: An approach to assess water use efficiency and growth strategies of dry land agroforestry species. Trees 2011, 25, 95–102. [Google Scholar] [CrossRef]
- Voltas, J.; Serrano, L.; Hernández, M.; Pemán, J. Carbon isotope discrimination, gas exchange and stem growth of four Euramerican hybrid poplars under different watering regimes. New Forests 2006, 31, 435–451. [Google Scholar] [CrossRef]
- Yin, C.; Wang, X.; Duan, B.; Luo, J.; Li, C. Early growth, dry matter allocation and water use efficiency of two sympatric Populus species as affected by water stress. Environ. Exp. Bot. 2005, 53, 315–322. [Google Scholar] [CrossRef]
- Dillen, S.; Marron, N.; Koch, B.; Ceulemans, R. Genetic variation of stomatal traits and carbon isotope discrimination in two hybrid poplar families (Populus deltoides ‘S9-2′ × P. nigra ‘Ghoy’ and P. deltoides ‘S9-2′ × P. trichocarpa ‘V24′). Annals Bot. 2008, 102, 399–407. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Delmotte, F.M.; Villar, M.; Delay, D.; Boudouresque, E.; Petit, J.-M.; Marron, N.; Bréchet, C.; Brignolas, F. Productivity, leaf traits and carbon isotope discrimination in 29 Populus deltoides × P. nigra clones. New Phytol. 2005, 167, 53–62. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.-M.; Barbaroux, C.; Le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides × P. nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Maier, C.A.; Burley, J.; Cook, R.; Ghezehei, S.B.; Hazel, D.W.; Nichols, E.G. Tree water use, water use efficiency, and carbon isotope discrimination in relation to growth potential in Populus deltoides and hybrids under field conditions. Forests 2019, 10, 993. [Google Scholar] [CrossRef]
- Verlinden, M.S.; Fichot, R.; Broeckx, L.S.; Vanholme, B.; Boerjan, W.; Ceulemans, R. Carbon isotope compositions (δ13C) of leaf, wood and holocellulose differ among genotypes of poplar and between previous land uses in a short-rotation biomass plantation. Plant Cell Environ. 2015, 38, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Orlović, S.; Galović, G.; Zorić, M.; Kovačević, B.; Pilipović, A.; Zoran, G. Evaluation of interspecific DNA variability in poplars using AFLP and SSR markers. Afr. J. Biotechnol. 2009, 8, 5241–5247. Available online: https://www.ajol.info/index.php/ajb/article/view/65955 (accessed on 25 August 2020).
- Eckenwalder, J.E. Natural intersectional hybridization between North American species of Populus (Salicaceae) in sections Aigeiros and Tacamahaca. II. Taxonomy. Can. J. Bot. 1984, 62, 325–335. [Google Scholar] [CrossRef]
- Nelson, N.D.; Berguson, W.E.; McMahon, B.G.; Cai, M.; Buchman, D.J. (2018) Growth performance and stability of hybrid poplar clones in simultaneous tests on six sites. Biomass Bioenergy 2018, 118, 115–125. [Google Scholar] [CrossRef]
- Nelson, N.D.; Berguson, W.E.; McMahon, B.G.; Meilan, R.; Smart, L.B.; Gouker, F.E.; Bloese, P.; Miller, R.; Volk, T.A.; Cai, M.; et al. Discovery of geographically robust hybrid poplar clones. Silvae Genet. 2019, 68, 101–110. [Google Scholar] [CrossRef]
- Headlee, W.L.; Zalesny, R.S., Jr.; Hall, R.B.; Bauer, E.O.; Bender, B.; Birr, B.A.; Miller, R.O.; Randall, J.A.; Wiese, A.H. Specific gravity of hybrid poplars in the north-central region, USA: Within-tree variability and site × genotype effects. Forests 2013, 4, 251–269. [Google Scholar] [CrossRef]
- Stanturf, J.A.; van Oosten, C.; Netzer, D.A.; Coleman, M.D.; Portwood, C.J. Ecology and silviculture of poplar plantations. Part A. In Poplar Culture in North America; Dickmann, D.I., Isebrands, J.G., Eckenwalder, J.G., Richardson, J., Eds.; NRC Research Press, National Research Council of Canada: Ottawa, ON, Canada, 2001; Chapter 5; pp. 153–206. ISBN 978-0-66018-145-5. [Google Scholar]
- Hjelm, K.; Rytter, L. The influence of soil conditions, with focus on soil acidity, on the establishment of poplar (Populus spp.). New Forests 2016, 47, 731–750. [Google Scholar] [CrossRef]
- Cabane, M.; Afif, D.; Hawkins, S. Lignins and abiotic stresses. Adv. Bot. Res. 2012, 61, 219–262. [Google Scholar] [CrossRef]
- Waliszewska, B.; Mleczek, M.; Zborowska, M.; Goliński, P.; Rutkowski, P.; Szentner, K. Changes in the chemical composition and the structure of cellulose and lignin in elm wood exposed to various forms of arsenic. Cellulose 2019, 26, 6303–6315. [Google Scholar] [CrossRef]
- Hausman, J.F.; Evers, D.; Thiellement, H.; Jouve, L. Compared responses of poplar cuttings and in vitro raised shoots to short-term chilling treatments. Plant Cell Rep. 2000, 19, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Klašjna, B.; Kopitović, S.; Orlović, S. Variability of some wood properties of eastern cottonwood (Populus deltoides Bartr.) clones. Wood Sci. Technol. 2003, 37, 331–337. [Google Scholar] [CrossRef]
- Sannigrahi, P.; Ragauskas, A.J.; Tuskan, G.A. Poplar as a feedstock for biofuels: A review of compositional characteristics. Biofuels Bioprod. Bioref. 2010, 4, 209–226. [Google Scholar] [CrossRef]
- Zhou, G.; Taylor, G.; Polle, A. FTIR-ATR-based prediction and modelling of lignin and energy contents reveals independent intra-specific variation of these traits in bioenergy poplars. Plant Methods 2011, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bauddh, K.; Singh, B.; Korstad, J. Phytoremediation Potential of Bioenergy Plants; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Studer, M.H.; DeMartini, J.D.; Davis, M.F.; Sykes, R.W.; Davison, B.; Keller, M.; Tuskan, G.A.; Wyman, C.E. Lignin content in natural Populus variants affects sugar release. Proc. Natl. Acad. Sci. USA 2011, 108, 6300–6305. [Google Scholar] [CrossRef]
- Chen, L.; Dou, J.; Ma, Q.; Li, N.; Wu, R.; Bian, H.; Yelle, D.J.; Vuorinen, T.; Fu, S.; Pan, X.; et al. Rapid and near-complete dissolution of wood lignin at ≤80 °C using a recyclable acid hydrotrope. Sci. Adv. 2017, 3, e1701735. [Google Scholar] [CrossRef]
- Sawada, D.; Kalluri, U.C.; O’Neill, H.; Urban, V.; Langan, P.; Davison, B.; Pingali, S.V. Tension wood structure and morphology conducive for better enzymatic digestion. Biotechnol. Biofuels 2018, 11, 44. [Google Scholar] [CrossRef]
- Leu, S.Y.; Zhu, J.Y. Substrate-related factors affecting enzymatic saccharification of lignocelluloses: Our recent understanding. BioEnergy Res. 2013, 6, 405–415. [Google Scholar] [CrossRef]
- Yu, Q.; Zhuang, X.; Wang, W.; Qi, W.; Wang, Q.; Tan, X.; Kong, X.; Yuan, Z. Hemicellulose and lignin removal to improve the enzymatic digestibility and ethanol production. Biomass Bioenergy 2016, 94, 105–109. [Google Scholar] [CrossRef]
- Volk, T.A.; Verwijst, T.; Tharakan, P.J.; Abrahamson, L.P.; White, E.H. Growing fuel: A sustainability assessment of willow biomass crops. Front. Ecol. Environ. 2004, 2, 411–418. [Google Scholar] [CrossRef]
- Ray, M.J.; Brereton, N.J.; Shield, I.; Karp, A.; Murphy, R.J. Variation in cell wall composition and accessibility in relation to biofuel potential of short rotation coppice willows. BioEnergy Res. 2012, 5, 685–698. [Google Scholar] [CrossRef]
- Bose, S.K.; Francis, R.C.; Govender, M.; Bush, T.; Spark, A. Lignin content versus syringyl to guaiacyl ratio amongst poplars. Bioresour. Technol. 2009, 100, 1628–1633. [Google Scholar] [CrossRef] [PubMed]
- Davison, B.H.; Drescher, S.R.; Tuskan, G.A.; Davis, M.F.; Nghiem, N.P. Variation of S/G ratio and lignin content in a Populus family influences the release of xylose by dilute acid hydrolysis. Appl. Biochem. Biotechnol. 2006, 130, 427–435. [Google Scholar] [CrossRef]
- Zalesny, R.S., Jr.; Stange, C.M.; Birr, B.A. Survival, height growth, and phytoextraction potential of hybrid poplar and Russian Olive (Elaeagnus Angustifolia L.) established on soils varying in salinity in North Dakota, USA. Forests 2019, 10, 672–691. [Google Scholar] [CrossRef]
- ANKOM Technology. Fiber Analyzer A200; ANKOM Technology: Macedon, NY, USA, 2020; Available online: https://www.ankom.com/technical-support/fiber-analyzer-a200 (accessed on 1 September 2020).
- Bond-Lamberty, B.; Wang, C.; Gower, S.T. Aboveground and belowground biomass and sapwood area allometric equations for six boreal tree species of northern Manitoba. Can. J. For. Res. 2002, 32, 1441–1450. [Google Scholar] [CrossRef]
- Headlee, W.L.; Zalesny, R.S., Jr. Allometric relationships for aboveground woody biomass differ among hybrid poplar genomic groups and clones in the North-Central USA. BioEnergy Res. 2019, 12, 966–976. [Google Scholar] [CrossRef]
- Luo, X.; Gleisner, R.; Tian, S.; Negron, J.; Horn, E.; Pan, X.J.; Zhu, J.Y. Evaluation of mountain beetle infested lodgepole pine for cellulosic ethanol production by SPORL pretreatment. Ind. Eng. Chem. Res. 2010, 49, 8258–8266. [Google Scholar] [CrossRef]
- Dence, C.W. The determination of lignin. In Methods in Lignin Chemistry; Lin, S.Y., Dence, C.W., Eds.; Springer: Berlin, Germany, 1992; pp. 33–61. [Google Scholar] [CrossRef]
- Gu, F.; Gilles, W.; Gleisner, R.; Zhu, J.Y. Fermentative high titer ethanol production from a Douglas-fir forest residue without detoxification using SPORL: High SO2 loading at a low temperature. Ind. Biotechnol. 2016, 12, 168–175. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | Age | Clone × Age | |
|---|---|---|---|
| Total carbon (%) | 0.3401 | 0.0023 | 0.4764 |
| Carbon isotope discrimination (Δ, ‰) | 0.9986 | <0.0001 | 0.1440 |
| BIOMASSMAI (Mg ha−1 yr−1) | <0.0001 | <0.0001 | <0.0001 |
| RELATIVEMAI | 0.6116 | <0.0001 | 0.0305 |
| Clone | Stage | Clone × Stage | |
| Hemicellulose (%) | 0.0995 | <0.0001 | 0.1809 |
| Cellulose (%) | 0.5900 | 0.0010 | 0.0013 |
| Lignin (%) | 0.0468 | 0.2193 | 0.6066 |
| Site | Clone | Site × Clone | |
| Mannan (%) | 0.0121 | 0.8283 | 0.1919 |
| Xylan (%) | <0.0001 | 0.9529 | 0.9341 |
| Glucan (%) | <0.0001 | 0.4084 | 0.8779 |
| Lignin (%) | <0.0001 | 0.5497 | 0.1504 |
| Substrate enzymatic digestibility (SED) (%) | <0.0001 | 0.1422 | 0.8719 |
| CTec3 loading (FPU g−1 glucan) | <0.0001 | 0.3324 | 0.8638 |
| Furfural in spent liquor (%) | 0.4385 | 0.7268 | 0.9538 |
| Xylose in spent liquor (%) | <0.0001 | 0.2598 | 0.7119 |
| Xylan in water insoluble solids (WIS) (%) | 0.0003 | 0.5373 | 0.7516 |
| Enzymatic hydrolysis glucose yield (EHGY) (%) | <0.0001 | 0.1619 | 0.5599 |
| Glucose in spent liquor (%) | 0.0025 | 0.1899 | 0.3148 |
| DN34 | NM6 | p-Value | |
|---|---|---|---|
| Height (m) | 14.8 ± 0.5 | 16.7 ± 0.7 | 0.0588 |
| Diameter (cm) | 12.0 ± 0.4 | 18.7 ± 0.4 | <0.0001 |
| BIOMASSMAI (Mg ha−1 yr−1), leafless | 1.4 ± 0.1 | 4.7 ± 0.4 | <0.0001 |
| BIOMASSMAI (Mg ha−1 yr−1), total | 1.5 ± 0.1 | 5.0 ± 0.4 | <0.0001 |
| Total carbon (%) | 48.3 ± 0.2 | 48.1 ± 0.2 | 0.5100 |
| CARBONMAI (Mg ha−1 yr−1), leafless | 0.7 ± 0.1 | 2.3 ± 0.2 | <0.0001 |
| CARBONMAI (Mg ha−1 yr−1), total | 0.7 ± 0.1 | 2.4 ± 0.2 | <0.0001 |
| Specific gravity | 0.363 ± 0.003 | 0.320 ± 0.007 | 0.0001 |
| Bole moisture (%) | 54.0 ± 0.3 | 59.0 ± 0.8 | <0.0001 |
| Branch moisture (%) | 53.8 ± 0.9 | 52.7 ± 0.6 | 0.3334 |
| Leaf | Bole | Branch | |||||||
|---|---|---|---|---|---|---|---|---|---|
| DN34 | NM6 | p-Value | DN34 | NM6 | p-Value | DN34 | NM6 | p-Value | |
| Total nitrogen (N) | 2.62 ± 0.12 | 2.33 ± 0.10 | 0.0777 | 0.28 ± 0.01 | 0.26 ± 0.02 | 0.3778 | 0.65 ± 0.04 | 0.56 ± 0.02 | 0.0510 |
| Total carbon (C) | 47.43 ± 0.10 | 47.23 ± 0.26 | 0.4819 | 48.27 ± 0.16 | 48.12 ± 0.15 | 0.5100 | 50.05 ± 0.15 | 49.84 ± 0.09 | 0.2465 |
| Calcium (Ca) | 10.05 ± 0.25 | 18.59 ± 1.62 | 0.0001 | 2.41 ± 0.43 | 1.84 ± 0.16 | 0.2390 | 3.39 ± 0.17 | 4.55 ± 0.36 | 0.0116 |
| Potassium (K) | 13.21 ± 0.82 | 10.80 ± 0.76 | 0.0487 | 5.39 ± 0.73 | 7.77 ± 1.03 | 0.0804 | 4.99 ± 0.25 | 4.29 ± 0.15 | 0.0306 |
| Magnesium (Mg) | 4.15 ± 0.24 | 4.25 ± 0.31 | 0.7938 | 1.13 ± 0.12 | 0.92 ± 0.04 | 0.1130 | 1.40 ± 0.08 | 1.11 ± 0.06 | 0.0100 |
| Phosphorus (P) | 2.07 ± 0.05 | 2.05 ± 0.08 | 0.7630 | 0.17 ± 0.01 | 0.14 ± 0.01 | 0.0646 | 0.86 ± 0.06 | 0.70 ± 0.03 | 0.0360 |
| Aluminum (Al) | 205.12 ± 31.06 | 314.15 ± 31.07 | 0.0264 | 245.40 ± 67.75 | 325.15 ± 36.67 | 0.3181 | 94.69 ± 21.66 | 347.98 ± 29.86 | <0.0001 |
| Cadmium (Cd) | 0.12 ± 0.08 | 0.20 ± 0.13 | 0.6453 | 0.16 ± 0.13 | 0.02 ± 0.02 | 0.3006 | 0.12 ± 0.09 | 0.37 ± 0.09 | 0.0666 |
| Chloride (Cl) | 341.61 ± 25.09 | 262.58 ± 15.89 | 0.0186 | Not tested | Not tested | - | Not tested | Not tested | - |
| Cobalt (Co) | 6.37 ± 2.21 | 5.35 ± 1.21 | 0.6909 | 13.99 ± 2.18 | 12.21 ± 0.91 | 0.4624 | 0.55 ± 0.55 | 3.57 ± 1.29 | 0.0493 |
| Chromium (Cr) | 5.55 ± 0.83 | 3.20 ± 0.96 | 0.0860 | 1.03 ± 0.74 | 0.00 ± 0.00 | 0.1873 | 1.24 ± 0.63 | 5.12 ± 1.69 | 0.0505 |
| Copper (Cu) | 5.40 ± 0.81 | 8.37 ± 1.51 | 0.1044 | 2.81 ± 0.39 | 8.08 ± 0.97 | 0.0002 | 4.86 ± 0.53 | 2.03 ± 0.42 | 0.0009 |
| Iron (Fe) | 504.40 ± 39.27 | 232.02 ± 20.15 | <0.0001 | 5.58 ± 1.45 | 1.79 ± 0.94 | 0.0454 | 89.12 ± 14.89 | 43.45 ± 6.65 | 0.0141 |
| Manganese (Mn) | 376.22 ± 27.99 | 231.50 ± 21.98 | 0.0012 | 65.11 ± 9.05 | 15.89 ± 4.06 | 0.0002 | 65.08 ± 8.61 | 33.56 ± 6.15 | 0.0099 |
| Sodium (Na) | 9.15 ± 0.66 | 6.49 ± 0.58 | 0.0089 | 14.62 ± 2.47 | 5.74 ± 1.70 | 0.0103 | 6.96 ± 0.62 | 4.14 ± 0.23 | 0.0008 |
| Nickel (Ni) | 4.32 ± 1.26 | 9.33 ± 2.33 | 0.0796 | 2.99 ± 0.86 | 4.34 ± 0.62 | 0.2277 | 4.63 ± 1.58 | 1.02 ± 0.89 | 0.0669 |
| Lead (Pb) | 7.00 ± 1.29 | 10.13 ± 1.64 | 0.1570 | 6.03 ± 2.02 | 9.69 ± 1.74 | 0.1921 | 5.26 ± 1.86 | 5.79 ± 1.83 | 0.8412 |
| Zinc (Zn) | 139.86 ± 12.08 | 169.37 ± 17.40 | 0.1854 | 25.63 ± 2.28 | 19.53 ± 1.28 | 0.0348 | 41.06 ± 2.09 | 43.45 ± 2.81 | 0.5055 |
| PHYTOMAI | ||||
|---|---|---|---|---|
| DN34 | NM6 | Difference | NM6 Advantage (%) | |
| ------------ kg ha−1 yr−1 ------------ | ||||
| Total nitrogen (N) | 6.5 | 19.3 | 12.8 | 196 |
| Total carbon (C) | 688.2 | 2302.1 | 1613.8 | 234 |
| Calcium (Ca) | 4.1 | 15.0 | 11.0 | 270 |
| Potassium (K) | 7.3 | 28.3 | 21.1 | 290 |
| Magnesium (Mg) | 1.8 | 4.8 | 3.0 | 169 |
| Phosphorus (P) | 0.7 | 2.0 | 1.3 | 174 |
| ------------- g ha−1 yr−1 ------------ | ||||
| Aluminum (Al) | 238.1 | 1581.9 | 1343.8 | 564 |
| Cadmium (Cd) | 0.2 | 0.9 | 0.7 | 368 |
| Cobalt (Co) | 10.2 | 37.1 | 26.9 | 264 |
| Chromium (Cr) | 1.6 | 12.0 | 10.4 | 657 |
| Copper (Cu) | 5.4 | 23.8 | 18.4 | 343 |
| Iron (Fe) | 66.3 | 106.3 | 40.0 | 60 |
| Manganese (Mn) | 91.1 | 116.2 | 25.1 | 28 |
| Sodium (Na) | 15.1 | 23.2 | 8.1 | 54 |
| Nickel (Ni) | 5.3 | 12.6 | 7.3 | 136 |
| Lead (Pb) | 7.9 | 36.4 | 28.5 | 360 |
| Zinc (Zn) | 46.7 | 148.0 | 101.3 | 217 |
| Chloride (Cl) (Leaves) | 34.2 | 78.8 | 44.6 | 131 |
| Site | Rhinelander, WI | Escanaba, MI |
|---|---|---|
| Application | Phytoremediation | Biomass |
| Latitude, Longitude | 45.6266° N, 89.3899° W | 45.7708° N, 87.1978° W |
| County | Oneida | Delta |
| Year Planted | 1999 | 2001 |
| Stocking (trees ha−1) | 834 | 1075 |
| Tree Age at Harvest (yr) | 17 | 9 |
| Height at Harvest (m) (mean ± one standard error) | 15.8 ± 0.5 | 12.0 ± 0.2 |
| Diameter at Harvest (cm) (mean ± one Standard error) | 15.4 ± 0.9 | 15.3 ± 0.7 |
| Annual Precipitation (P) (mm) | 571 ± 33 | 511 ± 11 |
| Average Temperature (Tavg) (°C) | 13.2 ± 0.2 | 13.5 ± 0.1 |
| Maximum Temperature (Tmax) (°C) | 19.6 ± 0.3 | 20.0 ± 0.1 |
| Minimum Temperature (Tmin) (°C) | 6.7 ± 0.2 | 6.9 ± 0.1 |
| Maximum—minimum Temperature (Tdiff) (°C) | 12.9 ± 0.2 | 13.1 ± 0.0 |
| Drought Index (abnormally dry) (D0) (%) | 45.0 ± 8.5 | 53.5 ± 4.8 |
| Drought Index (moderate drought) (D1) (%) | 25.1 ± 8.3 | 22.1 ± 3.2 |
| Drought Index (severe drought) (D2) (%) | 8.5 ± 3.9 | 9.3 ± 2.0 |
| Drought Index (extreme drought) (D3) (%) | 0.8 ± 0.8 | 0.2 ± 0.1 |
| Soil Depth (cm) | ||
|---|---|---|
| 0 to 10 | 10 to 30 | |
| Texture | Sandy loam | Sandy loam |
| Sand (%) | 61.9 | 69.5 |
| Silt (%) | 35.2 | 26.1 |
| Clay (%) | 2.9 | 4.4 |
| pH | 4.79 ± 0.06 | 4.51 ± 0.03 |
| Total nitrogen (%) | 0.18 ± 0.01 | 0.12 ± 0.01 |
| Total carbon (%) | 2.41 ± 0.12 | 1.31 ± 0.08 |
| --------------------- mg kg−1 --------------------- | ||
| Aluminum (Al) | 821.55 ± 28.08 | 1137.89 ± 25.43 |
| Calcium (Ca) | 667.23 ± 42.17 | 336.00 ± 24.39 |
| Cadmium (Cd) | 0.03 ± 0.01 | 0.05 ± 0.01 |
| Chloride (Cl) | 0.74 ± 0.09 | 0.90 ± 0.07 |
| Cobalt (Co) | 0.72 ± 0.06 | 0.87 ± 0.06 |
| Chromium (Cr) | 0.51 ± 0.06 | 0.62 ± 0.06 |
| Copper (Cu) | 2.92 ± 0.15 | 5.03 ± 0.94 |
| Iron (Fe) | 463.33 ± 7.23 | 489.73 ± 7.67 |
| Potassium (K) | 106.55 ± 4.80 | 59.74 ± 2.17 |
| Magnesium (Mg) | 150.20 ± 9.73 | 96.10 ± 7.03 |
| Manganese (Mn) | 129.66 ± 4.65 | 96.85 ± 6.11 |
| Sodium (Na) | 7.53 ± 1.61 | 15.03 ± 1.74 |
| Nickel (Ni) | 0.41 ± 0.05 | 0.78 ± 0.06 |
| Phosphorus (P) | 31.20 ± 1.59 | 25.36 ± 1.16 |
| Lead (Pb) | 2.07 ± 0.17 | 1.12 ± 0.11 |
| Zinc (Zn) | 3.31 ± 0.19 | 3.36 ± 0.38 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jr., R.S.Z.; Zhu, J.Y.; Headlee, W.L.; Gleisner, R.; Pilipović, A.; Acker, J.V.; Bauer, E.O.; Birr, B.A.; Wiese, A.H. Ecosystem Services, Physiology, and Biofuels Recalcitrance of Poplars Grown for Landfill Phytoremediation. Plants 2020, 9, 1357. https://doi.org/10.3390/plants9101357
Jr. RSZ, Zhu JY, Headlee WL, Gleisner R, Pilipović A, Acker JV, Bauer EO, Birr BA, Wiese AH. Ecosystem Services, Physiology, and Biofuels Recalcitrance of Poplars Grown for Landfill Phytoremediation. Plants. 2020; 9(10):1357. https://doi.org/10.3390/plants9101357
Chicago/Turabian StyleJr., Ronald S. Zalesny, J. Y. Zhu, William L. Headlee, Roland Gleisner, Andrej Pilipović, Joris Van Acker, Edmund O. Bauer, Bruce A. Birr, and Adam H. Wiese. 2020. "Ecosystem Services, Physiology, and Biofuels Recalcitrance of Poplars Grown for Landfill Phytoremediation" Plants 9, no. 10: 1357. https://doi.org/10.3390/plants9101357
APA StyleJr., R. S. Z., Zhu, J. Y., Headlee, W. L., Gleisner, R., Pilipović, A., Acker, J. V., Bauer, E. O., Birr, B. A., & Wiese, A. H. (2020). Ecosystem Services, Physiology, and Biofuels Recalcitrance of Poplars Grown for Landfill Phytoremediation. Plants, 9(10), 1357. https://doi.org/10.3390/plants9101357