Blepharostoma trichophyllum S.L. (Marchantiophyta): The Complex of Sibling Species and Hybrids



Abstract
:1. Introduction
2. Material and Methods
2.1. Specimen Collection
- (1)
- oil bodies size, shape, and number per cell;
- (2)
- the actual size and relative proportions of the cell length/width ratio in the middle part of the leaf segment. This parameter was measured as the size variation between the three middle cells in the cilia (thus, neither basal nor apical cells were taken into account);
- (3)
- middle cells in the leaf segments’ cross wall protrusions (dilation). These cells were called dilated if inter-cellular interval (cross wall junction in Wagner [11]) was wider than the cell width and made the cilia’s surface somewhat crenulate (not straight);
- (4)
- the stem’s cross-section features, including the cross-section’s actual size, the relative height of the section (measured as the cell number), the trigones, and the wall thickness distribution.
2.2. DNA Isolation, Amplification and Sequencing
2.3. Phylogenetic Analysis
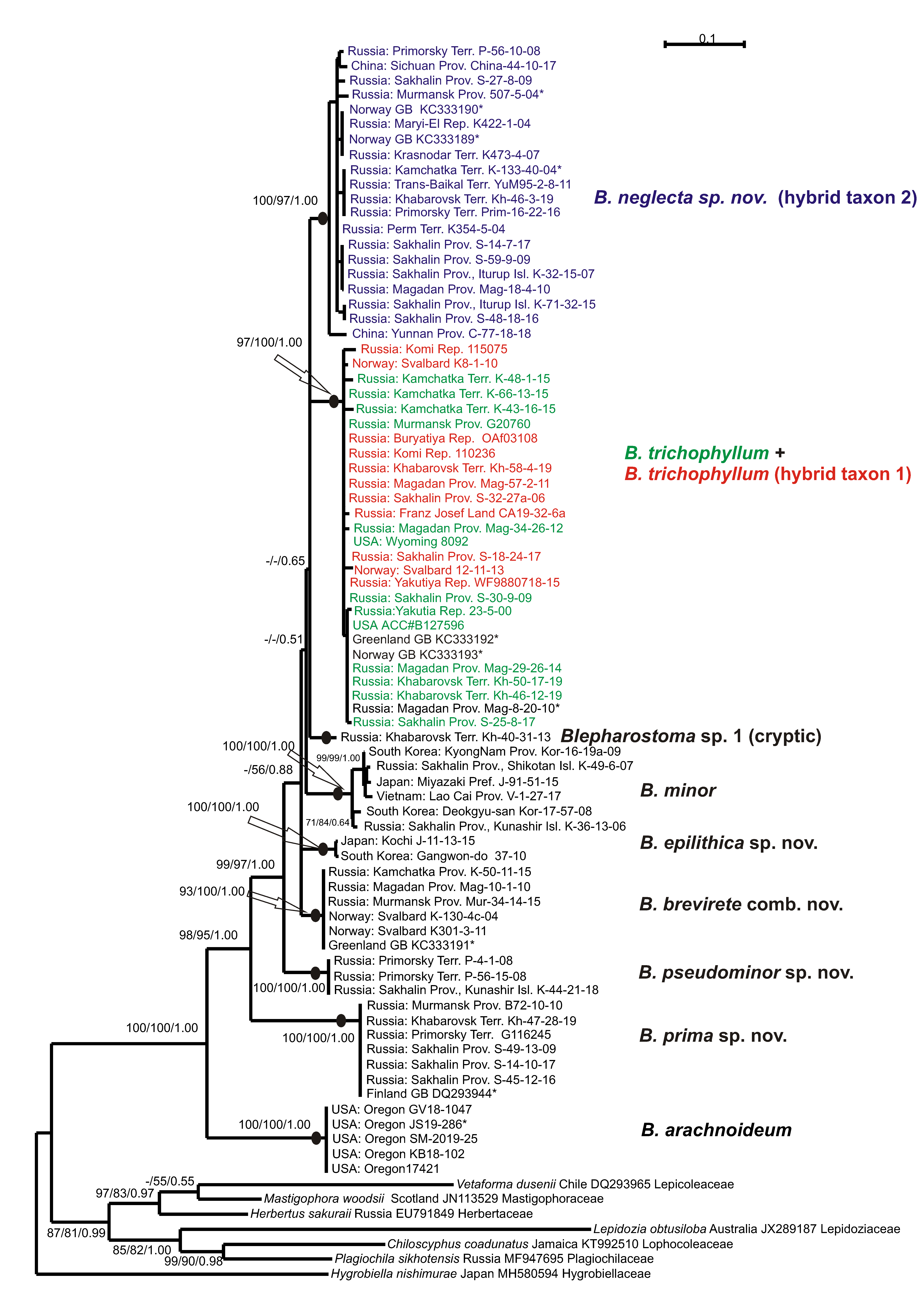
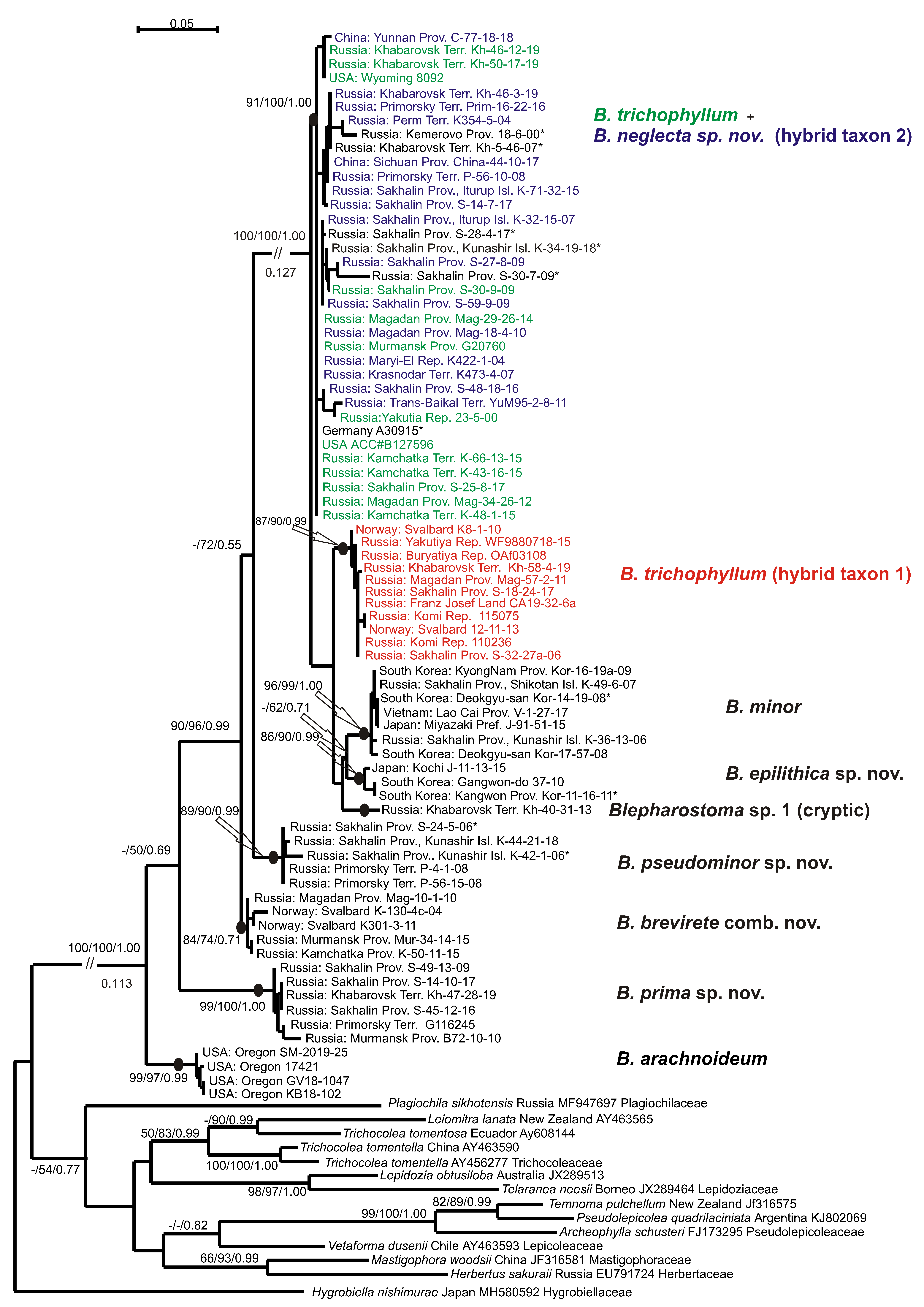
3. Results
4. Discussion
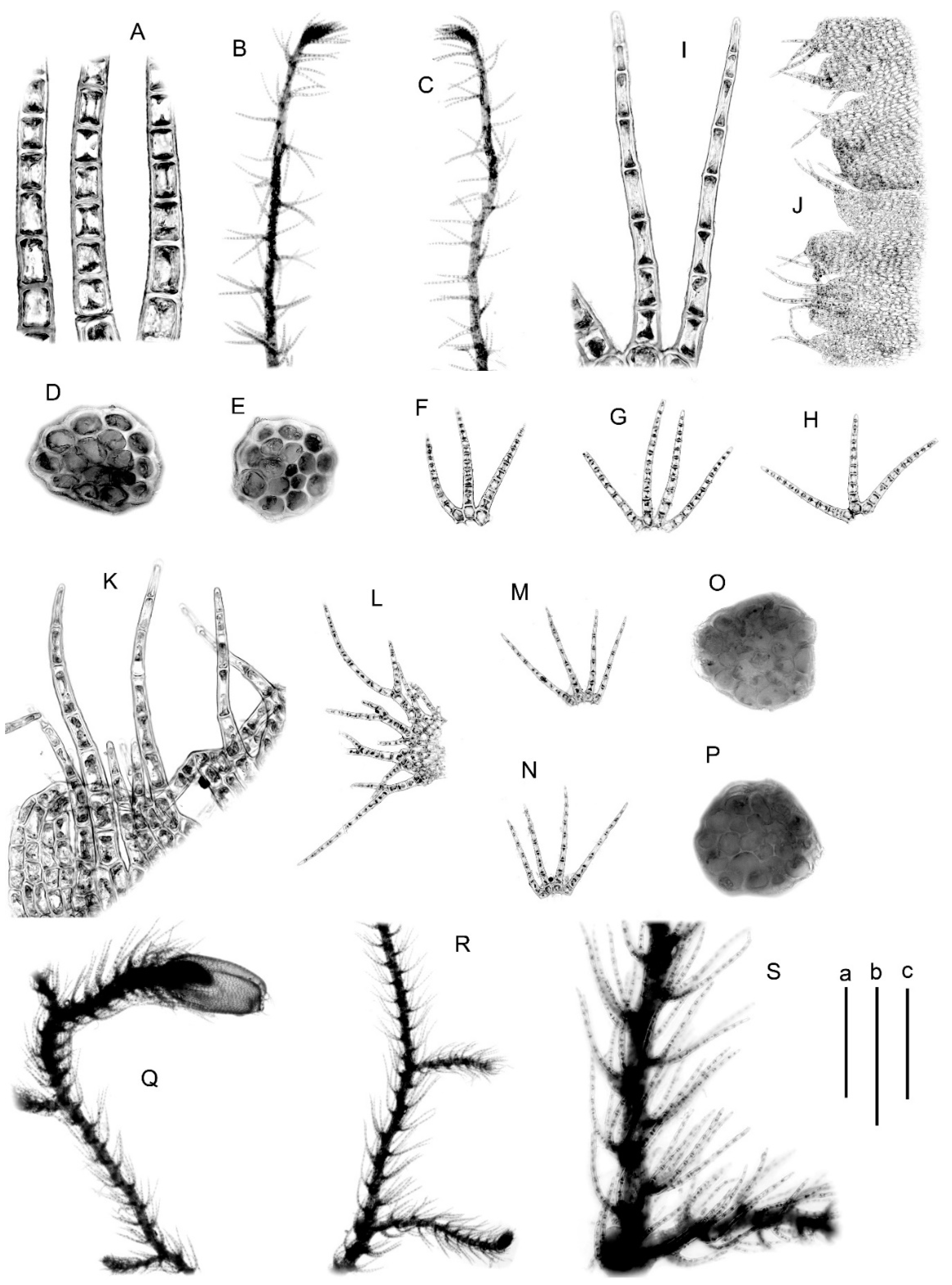
4.1. Morphology
4.2. Phytogeographic Considerations
5. Taxonomic Treatment
5.1. The Key
5.2. Descriptions
5.2.1. Blepharostoma prima Vilnet et Bakalin sp. nov.
5.2.2. Blepharostoma pseudominor Vilnet et Bakalin sp. nov.
5.2.3. Blepharostoma brevirete (Bryhn et Kaal.) Vilnet et Bakalin comb. nov. Basionym: Blepharostoma trichophyllum var. brevirete Bryhn et Kaal., Rep. Second Norweg. Arctic Exped. 11: 46, 1906. Syntypes: Canada. King Oskar Land, 76” 30′ N; Ellesmere Island, Framshavn (O, not seen).
5.2.4. Blepharostoma epilithica Vilnet et Bakalin sp. nov.
5.2.5. Blepharostoma minor Horik., Hikobia 1 (2): 104, 1951 [1952] Type: Japan, Settsu [Hyōgo] Pref. Toyono-gun, Mino Mt., Y. Horikawa, 5138, 23 November 1948, Holotype HIRO s.n.!
5.2.6. Cryptic Taxon (Tentatively Named ‘B. sp.’ in the Phylogenetic Tree)
5.2.7. Blepharostoma trichophyllum (L.) Dumort. (Green Colored Clade in the Figure 1 and Figure 2) Lectotype: United Kingdom,”, in Sylva Gleibergensi, Loco Fundusniger Dicto” (OXF, Not Seen)
5.2.8. Blepharostoma trichophyllum Hybrid Taxon 1 (Red Colored Clade in Figure 1 and Figure 2)
5.2.9. Blepharostoma neglecta Vilnet et Bakalin sp. nov. (Hybrid Taxon 2, Blue Colored in the Figure 1 and Figure 2)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schuster, R.M. The Hepaticae and Anthocerotae of North America East of the Hundredth Meridian. Vol. 1; Columbia University Press: New York, NY, USA, 1966; 802p. [Google Scholar]
- Damsholt, K. Illustrated Flora of Nordic Liverworts and Hornworts; Nordic Bryological Society: Lund, Sweden, 2002; 802p. [Google Scholar]
- Paton, J.A. The Liverwort Flora of the British Isles; Harley Books: Colchester, UK, 1999; 626p. [Google Scholar]
- Schljakov, R.N. Pechyenochnyye mkhi Severa SSSR [Liverworts of the North of the USSR]. Vol. 2; Nauka: Leningrad, Russia, 1979; 192p. [Google Scholar]
- Arnell, S.W. Hepaticae collected by O. Hedberg et al. on the East African Mountains. Ark. Bot. Ser. 2 1956, 3, 517–562. [Google Scholar]
- Söderström, L.; Hagborg, A.; Von Konrat, M.; Bartholomew-Began, S.; Bell, D.; Briscoe, L.; Brown, E.; Cargill, D.C.; Costa, D.P.; Crandall-Stotler, B.J.; et al. World checklist of hornworts and liverworts. PhytoKeys 2016, 59, 1–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laenen, B.; Shaw, B.; Schneider, H.; Goffinet, B.; Paradis, E.; Desamore, A.; Heinrichs, J.; Villarreal, J.C.; Gradstein, S.R.; McDaniel, S.F.; et al. Extant diversity of bryophytes emerged from successive post-Mesozoic diversification bursts. Nat. Commun. 2014, 5, 5134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrichs, J.; Hentschel, J.; Wilson, R.; Feldberg, K.; Schneider, H. Evolution of leafy liverworts (Jungermanniidae, Marchantiophyta): Estimating divergence times from chloroplast DNA sequences using penalized likelihood with integrated fossil evidence. Taxon 2007, 56, 31–44. [Google Scholar]
- He-Nygren, X.; Juslen, A.; Ahonen, I.; Glenny, D.; Piippo, S. Illuminating the evolutionary history of liverworts (Marchantiophyta)—Towards a natural classification. Cladistics 2006, 22, 1–31. [Google Scholar] [CrossRef]
- Hassel, K.; Segreto, R.; Ekrem, T. Restricted variation in plant barcoding markers limits identification in closely related bryophyte species. Mol. Ecol. Resour. 2013, 13, 1047–1057. [Google Scholar] [CrossRef]
- Wagner, D.H. Observations on Blepharostoma arachnoideum (Pseudolepicoleaceae) of western North America. Bryologist 2011, 114, 696–701. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Snisky, J.J., White, T.J., Eds.; Academic Press: San Diego, Brazil, 1990; pp. 315–322. [Google Scholar]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Goloboff, P.A.; Catalano, S.T.N.T. version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Hülsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar]
- Keane, T.M.; Naughton, T.J.; McInerney, J.O. ModelGenerator: Amino acid and nucleotide substitution model selection. 2004. Available online: http://bioinf.may.ie/software/modelgenerator/ (accessed on 20 March 2020).
- Pattengale, N.D.; Alipour, M.; Bininda-Emonds, O.R.P.; Moret, B.M.E.; Stamatakis, A. How many bootstrap replicates are necessary? J. Comput. Biol. 2010, 17, 337–354. [Google Scholar] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum Likelihood-based Phylogenetic Analyses with Thousands of Taxa and Mixed Models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar]
- Rambaut, A.; Drummond, A.J. Tracer v1.4. 2007. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 20 March 2020).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA 5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Method. Molec. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [PubMed] [Green Version]
- Vilnet, A.; Konstantinova, N.; Troitsky, A. Molecular phylogenetic data on reticulate evolution in the genus Barbilophozia Löske (Anastrophyllaceae, Marchantiophyta) and evidence of non-concerted evolution of rDNA in Barbilophozia rubescens allopolyploid. Phytotaxa 2012, 49, 6–22. [Google Scholar]
- Bakalin, V.; Vilnet, A.; Klimova, K.; Ma, W.Z.; Nguyen, V.S. Diplophyllum purpurascens (Scapaniaceae, Marchantiophyta)—A new species from Sino-Himalaya (China). Phytotaxa 2020, 447, 116–126. [Google Scholar]
- Mamontov, Y.S.; Vilnet, A.A.; Potemkin, A.D. Scapania marsupelloides sp. nov. (Scapaniaceae, Marchantiophyta), a remarkable new species near the base of Scapania phylogeny. Phytotaxa 2018, 385, 55–66. [Google Scholar]
- Bakalin, V.A.; Vilnet, A.A. Note on taxonomic position and distribution of Hygrobiella nishimurae N. Kitag. (Hygrobiellaceae, Marchantiophyta). Bot. Pacif. 2018, 7, 23–30. [Google Scholar]
- Asthana, G.; Saxena, M.; Maurya, M. A new species of Blepharostoma, B. indica sp. nov. from the western Himalaya, India, with observations on the closely allied B. trichophyllum (L.) Dumort. J. Bryol. 2013, 35, 266–269. [Google Scholar]
- Struck, T.H.; Feder, J.L.; Bendiksby, M.; Birkeland, S.; Cerca, J.; Gusarov, V.I.; Kistenich, S.; Larsson, K.H.; Liow, L.H.; Nowak, M.D.; et al. Finding Evolutionary Processes Hidden in Cryptic Species. Trends Ecol. Evol. 2018, 33, 153–163. [Google Scholar] [PubMed] [Green Version]
- Minelli, A. Taxonomy needs pluralism, but a controlled and manageable one. Megataxa 2020, 1, 9–18. [Google Scholar]
- Korshunova, T.; Martynov, A.; Bakken, T.; Picton, B. External diversity is restrained by internal conservatism: New nudibranch mollusc contributes to the cryptic species problem. Zool. Scr. 2017, 46, 683–692. [Google Scholar]
- Sun, H. Tethys retreat and Himalayas-Hengduanshan Mountains uplift and their significance on the origin and development of the Sino-Himalayan elements and alpine flora. Acta Bot. Yunnan. 2002, 24, 273–288. [Google Scholar]
- Sun, H.; Zhang, J.; Deng, T. Origins and evolution of plant diversity in the Hengduan Mountains, China. Plant Divers. 2017, 39, 161–166. [Google Scholar]
- Tada, R.; Zheng, H.; Clift, P.D. Evolution and variability of the Asian monsoon and its potential linkage with uplift of the Himalaya and Tibetan Plateau. Prog. Earth Planet. Sci. 2016, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Bakalin, V.A.; Klimova, K.G.; Nguyen, V.S. A review of Calypogeia (Marchantiophyta) in the eastern Sino-Himalaya and Meta-Himalaya based mostly on types. PhytoKeys 2020, 153, 111–154. [Google Scholar] [CrossRef]
- Bakalin, V.A.; Fedosov, V.E.; Maltseva, Y.D.; Milyutina, I.A.; Klimova, K.G.; Nguyen, H.M.; Troitsky, A.V. Overview of Schistochilopsis (Hepaticae) in Pacific Asia with the Description Protochilopsis gen. nov. Plants 2020, 9, 850. [Google Scholar]
- Bakalin, V.A.; Vilnet, A.A. Two new species of the liverwort genus Hygrobiella Spruce (Marchantiophyta) described from the North Pacific based on integrative taxonomy. Plant Syst. Evol. 2014, 300, 2277–2291. [Google Scholar]
- Frey, W.; Stech, M. New suprageneric taxa of liverworts (Marchantiophyta) and mosses (Bryophyta). Nova Hedwig. 2008, 87, 261–267. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Preliminary Identification | Country | Specimen Voucher | Collector | GenBank | |
|---|---|---|---|---|---|---|
| ITS1-2 nrDNA | trnL-F cpDNA | |||||
| B. arachnoideum | arachnoideum | USA: Oregon | SM-2019-25 (VBGI) | D. Wagner | MT586201 | MT585796 |
| B. arachnoideum | arachnoideum | USA: Oregon | KB18-102 (VBGI) | D. Wagner | MT586202 | MT585797 |
| B. arachnoideum | arachnoideum | USA: Oregon | 17421 (VBGI) | D. Wagner | MT586203 | MT585798 |
| B. arachnoideum | arachnoideum | USA: Oregon | JS19-286 (VBGI) | S. Jessup | MT586204 | no data |
| B. arachnoideum | arachnoideum | USA: Oregon | GV18-1047 (VBGI) | D. Wagner | MT586205 | MT585799 |
| B. brevirete comb. nov. | trichophyllum subsp. brevirete | Greenland | K. Hassel, T. Presto | KC333191 | no data | |
| B. brevirete comb. nov. | trichophyllum subsp. brevirete | Norway: Svalbard | K-130-4c-04, 110,026 (KPABG) | N. Konstantinova | MT586187 | MT585780 |
| B. brevirete comb. nov. | trichophyllum | Norway: Svalbard | K301-3-11, 115,395 (KPABG) | N. Konstantinova | MT586188 | MT585781 |
| B. brevirete comb. nov. | trichophyllum subsp. brevirete | Russia: Kamchatka Prov. | K-50-11-15, 300,157 (VBGI) | V. Bakalin | MT586189 | MT585782 |
| B. brevirete comb. nov. | trichophyllum subsp. brevirete | Russia: Magadan Prov. | Mag-10-1-10, 313,792 (VBGI), 115,163 (KPABG) | V. Bakalin | MT586190 | MT585783 |
| B. brevirete comb. nov. | trichophyllum | Russia: Murmansk Prov. | Mur-34-14-15, 308,832 (VBGI) | V. Bakalin | MT586191 | MT585784 |
| B. epilithica sp. nov. | minor | Japan: Kochi | J-11-13-15 (VBGI) | V. Bakalin | MT586185 | MT585777 |
| B. epilithica sp. nov. | trichophyllum | South Korea: Gangwon Prov. | 37-10, 115,503 (KPABG) | S.-S. Choi | MT586186 | MT585778 |
| B. epilithica sp. nov. | minor | South Korea: Gangwon Prov. | Kor-11-16-11, 317,266 (VBGI), 115,597 (KPABG) | V. Bakalin | no data | MT585779 |
| B. minor | minor | Japan: Miyazaki Pref. | J-91-51-15, 303,782 (VBGI) | V. Bakalin | MT586179 | MT585770 |
| B. minor | minor | Russia: Sakhalin Prov., Kuril I., Kunashir Isl. | K-36-13-06, 311,880 (VBGI), 115,146 (KPABG) | V. Bakalin | MT586180 | MT585771 |
| B. minor | minor | Russia: Sakhalin Prov., Kuril I., Shikotan Isl. | K-49-6-07, 313,369 (VBGI), 115,152 (KPABG) | V. Bakalin | MT586181/MT586210 | MT585772 |
| B. minor | minor | South Korea: Jeonbuk Prov. | Kor-14-19-08, 311,767 (VBGI), 115,150 (KPABG) | V. Bakalin | no data | MT585773 |
| B. minor | minor | South Korea: Jeonbuk Prov. | Kor-17-57-08, 311,857 (VBGI), 115,151 (KPABG) | V. Bakalin | MT586182/MT586211 | MT585774 |
| B. minor | minor | South Korea: Gyeongnam Prov. | Kor-16-19a-09, 317,099 (VBGI), 115,153 (KPABG) | V. Bakalin | MT586183 | MT585775 |
| B. minor | minor | Vietnam: Lao Cai Prov. | V-1-27-17, 35,095 (VBGI), 122,638 (KPABG) | V. Bakalin & K.Klimova | MT586184 | MT585776 |
| B. prima sp. nov. | trichophyllum | Finland | He-Nygren &Piippo | DQ293944 | no data | |
| B. prima sp. nov. | trichophyllum | Russia: Khabarovsk Terr. | kh-47-28-19 (VBGI) | V. Bakalin | MT586195 | MT585790 |
| B. prima sp. nov. | trichophyllum | Russia: Murmansk Prov. | B72-10-10, 20,464 (KPABG) | O. Belkina | MT586196 | MT585791 |
| B. prima sp. nov. | minor | Russia: Primorsky Terr. | G116245 (KPABG) | E. Borovichev | MT586197 | MT585792 |
| B. prima sp. nov. | aff. trichophyllum | Russia: Sakhalin Prov. | S-14-10-17, 122,635 (KPABG) | V. Bakalin | MT586198 | MT585793 |
| B. prima sp. nov. | trichophyllum | Russia: Sakhalin Prov. | S-45-12-16 (VBGI) | V. Bakalin | MT586199 | MT585794 |
| B. prima sp. nov. | trichophyllum | Russia: Sakhalin Prov. | S-49-13-09, 309,815 (VBGI), 115,158 (KPABG) | V. Bakalin | MT586200 | MT585795 |
| B. pseudominor sp. nov. | trichophyllum | Russia: Primorsky Terr. | P-4-1-08, 310,194 (VBGI), 115,159 (KPABG) | V. Bakalin | MT586192 | MT585785 |
| B. pseudominor sp. nov. | Russia: Primorsky Terr. | P-56-15-08 (VBGI) | V. Bakalin | MT586193 | MT585786 | |
| B. pseudominor sp. nov. | cf. minor | Russia: Sakhalin Prov. | S-24-5-06, 312,731 (VBGI), 115,149 (KPABG) | V. Bakalin | no data | MT585787 |
| B. pseudominor sp. nov. | minor | Russia: Sakhalin Prov., Kuril I., Kunashir Isl. | K-42-1-06, 312,097 (VBGI), 110,259 (KPABG) | V. Bakalin | no data | MT585788 |
| B. pseudominor sp. nov. | minor | Russia: Sakhalin Prov., Kuril I., Kunashir Isl. | K-44-21-18, 57,923 (VBGI), 122,499 (KPABG) | V. Bakalin, K.Klimova | MT586194 | MT585789 |
| Possible cryptic taxon tentatively named as ‘B. sp.’ | brevirete | Russia: Khabarovsk Terr. | Kh-40-31-13, 302,859 (VBGI) | V. Bakalin | MT586178 | MT585769 |
| B. trichophyllum (“green” parent) | brevirete | Russia: Magadan Prov. | Mag-29-26-14, 301,897 (VBGI) | V. Bakalin | MT586135 | MT585723 |
| B. trichophyllum (“green” parent) | trichophyllum | Russia: Magadan Prov. | Mag-34-26-12, 306,377 (VBGI) | V. Bakalin | MT586136 | MT585724 |
| B. trichophyllum (“green” parent) | trichophyllum | Russia: Murmansk Prov. | G20760 (KPABG) | E. Borovichev | MT586137 | MT585725 |
| B. trichophyllum (“green” parent) | minus | Russia: Kamchatka Terr. | K-48-1-15, 300,081 (VBGI) | V. Bakalin | MT586138 | MT585726 |
| B. trichophyllum (“green” parent) | brevirete | Russia: Kamchatka Terr. | K-66-13-15, 300,362 (VBGI) | V. Bakalin | MT586139 | MT585727 |
| B. trichophyllum (“green” parent) | trichophyllum | Russia: Kamchatka Terr. | K-43-16-15, 300,019 (VBGI) | V. Bakalin | MT586140 | MT585728 |
| B. trichophyllum (“green” parent) | trichophyllum | Russia: Khabarovsk Terr. | Kh-46-12-19 (VBGI) | V. Bakalin | MT586141 | MT585729 |
| B. trichophyllum (“green” parent) | trichophyllum | Russia: Khabarovsk Terr. | Kh-50-17-19 (VBGI) | V. Bakalin | MT586142 | MT585730 |
| B. trichophyllum (“green” parent) | trichophyllum | |||||
| B. trichophyllum (“green” parent) | trichophyllum var. brevirete | Russia: Sakhalin Prov. | S-25-8-17, 122,637 (KPABG) | V. Bakalin | MT586143 | MT585731 |
| B. trichophyllum (“green” parent) | trichophyllum | Russia: Sakhalin Prov. | S-30-9-09, 309,613 (VBGI), 115,162 (KPABG) | V. Bakalin | MT586144 | MT585732 |
| B. trichophyllum (“green” parent) | trichophyllum | Russia: Yakutia Rep. | 23-5-00, 101,623 (KPABG) | V. Bakalin | MT586145 | MT585733 |
| B. trichophyllum (“green” parent) | trichophyllum | USA: Wyoming | 8092 (VBGI) | Kosovich | MT586146 | MT585734 |
| B. trichophyllum (“green” parent) | trichophyllum | USA | ACC#B127596 (KPABG) | Schofield W., R. Belland, T. Hedderson | MT586147 | MT585735 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Norway: Svalbard | K-8-1-10, 113,992 (KPABG) | N. Konstantinova, A. Savchenko | MT586168 | MT585758 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Norway: Svalbard | 12-11-13, 116,486 (KPABG) | N. Koroleva | MT586169/MT586208 | MT585759 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Russia: Arkhangelsk Prov., Franz Josef Land, Jekson Isl. | CA19-32-6a | A. Savchenko | MT586148 | MT585760 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Russia: Buryatiya Rep. | OAf03108, 113,959 (KPABG) | O. Afonina | MT586170 | MT585761 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Russia: Khabarovsk Terr. | Kh-58-4-19 (VBGI) | V. Bakalin | MT586171 | MT585762 |
| B. trichophyllum hybrid taxon 1 | trichophyllum | Russia: Komi Rep. | 110236 (KPABG) | M. Dulin | MT586172 | MT585763 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Russia: Komi Rep. | 115075 (KPABG) | M. Dulin | MT586173/MT586209 | MT585764 |
| B. trichophyllum hybrid taxon 1 | trichophyllum | Russia: Magadan Prov. | Mag-57-2-11, 316,644 (VBGI), | V. Bakalin | MT586174 | MT585765 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Russia: Sakhalin Prov. | S-18-24-17, 122,634 (KPABG) | V. Bakalin | MT586175 | MT585766 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Russia: Sakhalin Prov. | S-32-27a-06, 115,166 (KPABG), 313,003 (VBGI) | V. Bakalin | MT586176 | MT585767 |
| B. trichophyllum hybrid taxon 1 | trichophyllum var. brevirete | Russia: Yakutiya Rep. | WF9880718-15 (KPABG) | Filin | MT586177 | MT585768 |
| B. neglecta sp. nov. (hybrid taxon 2) | aff. trichophyllum | China: Sichuan Prov. | China-44-10-17, 122,632 (KPABG), 37,380 (VBGI) | V. Bakalin, K. Klimova | MT586150 | MT585736 |
| B. neglecta sp. nov. (hybrid taxon 2) | sp. indet. | China: Yunnan Prov. | C-77-18-18 (VBGI) | V. Bakalin | MT586151 | MT585737 |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Norway | H. Blom | KC333189 | no data | |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Norway | J. Jordal | KC333190 | no data | |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Kamchatka Terr. | K-133-40-04, 115,167 (KPABG), 309,158 (VBGI) | V. Bakalin | MT586152 | no data |
| B. neglecta sp. nov. (hybrid taxon 2) | Russia: Khabarovsk Terr. | Kh-46-3-19 (VBGI) | V. Bakalin | MT586153 | MT585738 | |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Krasnodar Terr. | K473-4-07, 111,729 (KPABG) | N. Konstantinova | MT586154 | MT585739 |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Magadan Prov. | Mag-18-4-10, 115,164 (KPABG), 313,978 (VBGI) | V. Bakalin | MT586155 | MT585740 |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Maryi-El Rep. | K422-1-04, 107,997 (KPABG) | N. Konstantinova | MT586156/MT586206 | MT585741 |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Murmansk Prov. | 507-5-04, 12,268 (KPABG) | N. Konstantinova | MT586157 | no data |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Perm Terr. | K354-5-04, 108,337 (KPABG) | N. Konstantinova | MT586158/MT586207 | MT585742 |
| B. neglecta sp. nov. (hybrid taxon 2) | minor | Russia: Primorsky Terr. | P-56-10-08, 115,148 (KPABG) | V. Bakalin | MT586159 | MT585743 |
| B. neglecta sp. nov. (hybrid taxon 2) | sp. | Russia: Primorsky Terr. | Prim-16-22-16 (VBGI) | V. Bakalin | MT586160 | MT585744 |
| B. neglecta sp. nov. (hybrid taxon 2) | aff. trichophyllum | Russia: Sakhalin Prov. | S-14-7-17, 122,636 (RPABG) | V. Bakalin | MT586161 | MT585745 |
| B. neglecta sp. nov. (hybrid taxon 2) | minor | Russia: Sakhalin Prov. | S-27-8-09, 115,154 (KPABG), 309,566 (VBGI) | V. Bakalin | MT586162 | MT585746 |
| B. neglecta sp. nov. (hybrid taxon 2) | cf. minus | Russia: Sakhalin Prov. | S-48-18-16 (VBGI) | V. Bakalin | MT586163 | MT585747 |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Sakhalin Prov. | S-59-9-09, 309,959 (VBGI), 115,156 (KPABG) | V. Bakalin | MT586164 | MT585748 |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Sakhalin Prov., Kuril I., Iturup Isl. | K-32-15-07, 311,588 (VBGI), 115,160 (KPABG) | V. Bakalin | MT586165 | MT585749 |
| B. neglecta sp. nov. (hybrid taxon 2) | minor | Russia: Sakhalin Prov., Kuril I., Iturup Isl. | K-71-32-15, 308,208 (VBGI) | V. Bakalin | MT586166 | MT585751 |
| B. neglecta sp. nov. (hybrid taxon 2) | trichophyllum | Russia: Trans-Baikal Terr. | YuM95-2-8-11, 115,321 (KPABG) | Yu. Mamontov | MT586167 | MT585752 |
| B. trichophyllum unclear taxonomic position * | trichophyllum subsp. brevirete | Norway | K. Hassel, T. Presto | KC333193 | no data | |
| B. trichophyllum unclear taxonomic position * | trichophyllum subsp. brevirete | Greenland | K. Hassel, T. Presto | KC333192 | no data | |
| B. trichophyllum unclear taxonomic position * | trichophyllum | Russia: Magadan Prov. | Mag-8-20-10, 313,747 (VBGI), 115,169 (KPABG) | V. Bakalin | MT586149 | no data |
| B. trichophyllum unclear taxonomic position * | cf. minor | Russia: Sakhalin Prov., Kuril I., Kunashir Isl. | K-34-19-18, 57,099 (VBGI), 122,498 (KPABG) | V. Bakalin, K.Klimova | no data | MT585750 |
| B. trichophyllum unclear taxonomic position * | trichophyllum | Germany | A30915 (KPABG) | no data | no data | MT585753 |
| B. trichophyllum unclear taxonomic position * | trichophyllum | Russia: Kemerovo Prov. | 18-6-00, 101,790 (KPABG) | N. Konstantinova | no data | MT585754 |
| B. trichophyllum unclear taxonomic position * | Trichophyllum | Russia: Khabarovsk Terr. | Kh-5-46-07, 115,161 (KPABG) | V. Bakalin | no data | MT585755 |
| B. trichophyllum unclear taxonomic position * | Trichophyllum | Russia: Sakhalin Prov. | S-28-4-17, 122,633 (KPABG) | V. Bakalin | no data | MT585756 |
| B. trichophyllum unclear taxonomic position * | Trichophyllum | Russia: Sakhalin Prov. | S-30-7-09, 309,611 (VBGI), 115,168 (KPABG) | V. Bakalin | no data | MT585757 |
| Species | Country | Specimen Voucher | Collector | GenBank | |
|---|---|---|---|---|---|
| ITS1-2 nrDNA | trnL-F cpDNA | ||||
| Archeophylla schusteri (E.A.Hodgs. et Allison) R.M.Schust. | no data | FJ173295 | |||
| Chiloscyphus coadunatus (Sw.) J.J. Engel & R.M. Schust. | Jamaica | 35035 (JE) | Schaefer-Verwimp & Verwimp | KT992510 | no data |
| Herbertus sakuraii (Warnst.) S. Hatt. | Russia | P-74-15-05 (KPABG) | V. Bakalin | EU791849 | EU791724 |
| Hygrobiella nishimurae N. Kitag. | Japan | ExJ-11-42-15 (VBGI, KPABG) | V. Bakalin | MH580594 | MH580592 |
| Leiomitra lanata (Hook.) R.M.Schust. | New Zealand | 8521 | D. Glenny | no data | AY463565 |
| Lepidozia obtusiloba Steph. | Australia | NSW 110 | E. Cooper | JX289187 | JX289513 |
| Mastigophora woodsii (Hook.) Nees | China | 33696 (E) | D. Long | no data | JF316581 |
| Scotland | D. Long | JN113529 | no data | ||
| Plagiochila sikhotensis Bakalin & Vilnet | Russia | V. Bakalin | MF947695 | MF947697 | |
| Pseudolepicolea quadrilaciniata (Sull.) Fulford et J.Taylor. | Argentina | 31658 (E) | D. Long | no data | KJ802069 |
| Telaranea neesii (Lindenb.) Fulford | Borneo | NSW | E. Brown | no data | JX289464 |
| Temnoma pulchellum (Hook.) Mitt. | New Zealand | 92/87 (F) | J.Braggins | no data | JF316575 |
| Trichocolea tomentosa (Sw.) Gottsche | Ecuador | 368 (DUKE) | K. Davis | no data | AY608144 |
| Trichocolea tomentella (Ehrh.) Dumort. | China | 1137 | He-Nygren | no data | AY463590 |
| Trichocolea tomentella (Ehrh.) Dumort. | no data | AY456277 | |||
| Vetaforma dusenii (Steph.) Fulford et J.Taylor | Chile | 11423 (H) | J.J. Engel | DQ293965 | AY463593 |
| Number | Taxon | Infraspecific P-Distances, ITS1-2/trnL-F, % | Infrageneric P-Distances, ITS1-2/trnL-F, % | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| 1 | B. trichophyllum | 0.9/0.4 | ||||||||||
| 2 | B. neglecta sp. nov. hybrid 2 | 1.4/1.1 | 8.1/0.9 | |||||||||
| 3 | B. trichophyllum hybrid 1 | 1.2/0.2 | 1.4/2.6 | 8.3/3.2 | ||||||||
| 4 | B. ‘sp’ cryptic taxon | n/c/n/c | 6.9/3.3 | 6.7/3.9 | 7.1/2.7 | |||||||
| 5 | B. minor | 2.0/0.4 | 10.5/3.8 | 10.4/4.4 | 10.6/3.5 | 9.4/3.6 | ||||||
| 6 | B. epilytica sp. nov. | 0.6/0.6 | 7.5/3.2 | 7.3/3.7 | 7.8/3.2 | 6.8/3.3 | 9.1/3.5 | |||||
| 7 | B. brevirete comb. nov. | 0.0/0.5 | 6.9/6.8 | 6.6/7.4 | 7.0/6.4 | 5.7/5.4 | 8.4/7.2 | 5.8/7.4 | ||||
| 8 | B. pseudominor sp. nov. | 0.2/0.5 | 10.3/8.3 | 9.4/9.0 | 10.3/7.7 | 9.4/6.8 | 11.2/7.9 | 10.2/8.8 | 7.7/3.3 | |||
| 9 | B. prima sp. nov. | 0.2/0.5 | 16.6/12.7 | 16.6/13.4 | 16.7/13.1 | 17.2/11.2 | 18.5/12.9 | 16.3/12.8 | 15.3/8.8 | 16.6/10.3 | ||
| 10 | B. arachnoideum | 0.0/0.3 | 20.0/11.5 | 19.3/12.3 | 19.6/11.9 | 20.1/9.9 | 20.7/12.2 | 20.1/13.1 | 18.5/7.7 | 18.6/9.8 | 21.9/9.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalin, V.A.; Vilnet, A.A.; Choi, S.S.; Nguyen, V.S. Blepharostoma trichophyllum S.L. (Marchantiophyta): The Complex of Sibling Species and Hybrids. Plants 2020, 9, 1423. https://doi.org/10.3390/plants9111423
Bakalin VA, Vilnet AA, Choi SS, Nguyen VS. Blepharostoma trichophyllum S.L. (Marchantiophyta): The Complex of Sibling Species and Hybrids. Plants. 2020; 9(11):1423. https://doi.org/10.3390/plants9111423
Chicago/Turabian StyleBakalin, Vadim A., Anna A. Vilnet, Seung Se Choi, and Van Sinh Nguyen. 2020. "Blepharostoma trichophyllum S.L. (Marchantiophyta): The Complex of Sibling Species and Hybrids" Plants 9, no. 11: 1423. https://doi.org/10.3390/plants9111423
APA StyleBakalin, V. A., Vilnet, A. A., Choi, S. S., & Nguyen, V. S. (2020). Blepharostoma trichophyllum S.L. (Marchantiophyta): The Complex of Sibling Species and Hybrids. Plants, 9(11), 1423. https://doi.org/10.3390/plants9111423