Plant Aspartic Proteases for Industrial Applications: Thistle Get Better


Abstract
:1. Introduction
2. The Thistle Flower and the Impact of Its Variability on Milk-Clotting Activity
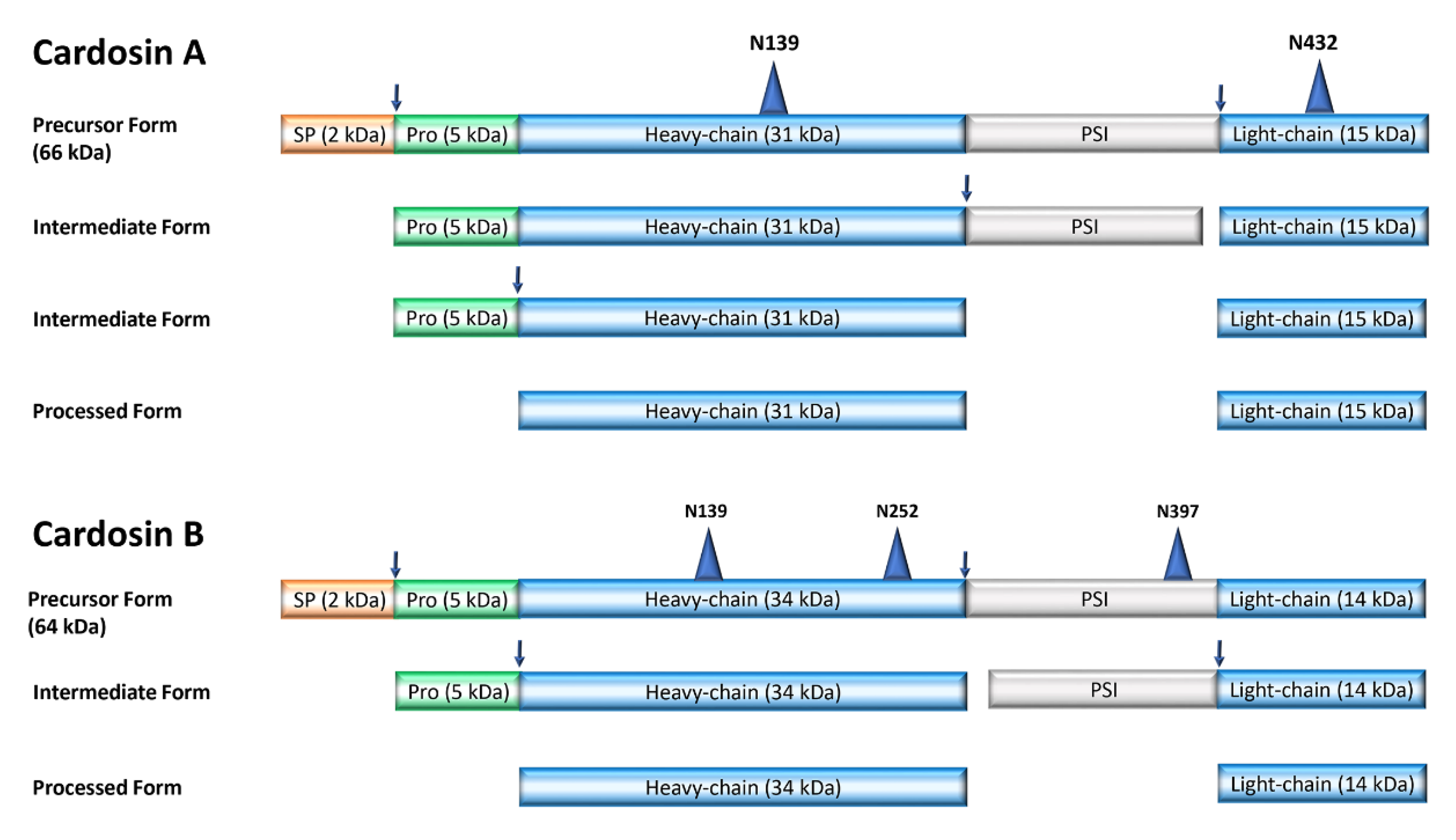
3. Cellular and Intracellular Localization of Cardosins
4. Production of Thistle APs Using Alternative Platforms
5. Molecular Farming of Aspartic Proteases
6. Challenges and Opportunities
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feijoo-Siota, L.; Villa, T.G. Native and Biotechnologically Engineered Plant Proteases with Industrial Applications. Food Bioprocess Technol. 2011, 4, 1066–1088. [Google Scholar] [CrossRef]
- Coelho, D.A.; Carrola, T.E.P.; Couvinhas, A.F. Improvement of certified artisan cheese production through systemic analysis-Serra da Estrela PDO. Sustainability 2017, 9, 468. [Google Scholar] [CrossRef] [Green Version]
- Rolet-Répécaud, O.; Arnould, C.; Dupont, D.; Gavoye, S.; Beuvier, E.; Achilleos, C. Quantification of pepsin in rennet using a monoclonal antibody-based inhibition ELISA. LWT Food Sci. Technol. 2017, 76, 190–196. [Google Scholar] [CrossRef]
- Longenbach, J.I.; Heinrichs, A.J. A review of the importance and physiological role of curd formation in the abomasum of young calves. Anim. Feed Sci. Technol. 1998, 73, 85–97. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Moreno-Hernández, J.M.; Ramírez-Suarez, J.C. Milk-Clotting Plant Proteases for Cheesemaking. In Biotechnological Applications of Plant Proteolytic Enzymes; Guevara, M.G., Daleo, G.R., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 21–41. ISBN 978-3-319-97132-2. [Google Scholar]
- Mazorra-Manzano, M.A.; Perea-Gutiérrez, T.C.; Lugo-Sánchez, M.E.; Ramirez-Suarez, J.C.; Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Cordoba, B. Comparison of the milk-clotting properties of three plant extracts. Food Chem. 2013, 141, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- Roseiro, L.B.; Barbosa, M.; Ames, J.M.; Wilbey, R.A. Cheesemaking with vegetable coagulants—The use of Cynara L. for the production of ovine milk cheeses. Int. J. Dairy Technol. 2003, 56, 76–85. [Google Scholar] [CrossRef]
- Soares, A.; Ribeiro Carlton, S.M.; Simões, I. Atypical and nucellin-like aspartic proteases: Emerging players in plant developmental processes and stress responses. J. Exp. Bot. 2019, 70, 2059–2076. [Google Scholar] [CrossRef]
- Faro, C.; Gal, S. Aspartic Proteinase Content of the Arabidopsis Genome. Curr. Protein Pept. Sci. 2005, 6, 493–500. [Google Scholar] [CrossRef]
- Simões, I.; Faro, C. Structure and function of plant aspartic proteinases. Eur. J. Biochem. 2004, 271, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Castanheira, P.; Samyn, B.; Sergeant, K.; Clemente, J.C.; Dunn, B.M.; Pires, E.; Van Beeumen, J.; Faro, C. Activation, proteolytic processing, and peptide specificity of recombinant cardosin A. J. Biol. Chem. 2005, 280, 13047–13054. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Pereira, S.; Satiat-Jeunemaitre, B.; Pissarra, J. Cardosin A contains two vacuolar sorting signals using different vacuolar routes in tobacco epidermal cells. Plant J. 2013, 76, 87–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, M.E.; D’Ippolito, S.; Pepe, A.; Daleo, G.R.; Guevara, M.G. Transgenic expression of plant-specific insert of potato aspartic proteases (StAP-PSI) confers enhanced resistance to Botrytis cinerea in Arabidopsis thaliana. Phytochemistry 2018, 149, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Conceição, C.; Martins, P.; Alvarenga, N.; Dias, J.; Lamy, E.; Garrido, L.; Gomes, S.; Freitas, S.; Belo, A.; Brás, T.; et al. Cynara cardunculus: Use in Cheesemaking and Pharmaceutical Applications. Technol. Approaches Nov. Appl. Dairy Process. 2018, 73. [Google Scholar]
- Barracosa, P.; Oliveira, J.; Barros, M.; Pires, E. Morphological evaluation of cardoon (Cynara cardunculus L.): Assessing biodiversity for applications based on tradition, innovation and sustainability. Genet. Resour. Crop Evol. 2018, 65, 17–28. [Google Scholar] [CrossRef]
- Barracosa, P.; Rosa, N.; Barros, M.; Pires, E. Selected Cardoon (Cynara cardunculus L.) Genotypes Suitable for PDO Cheeses in Mediterranean Regions. Chem. Biodivers. 2018, 15. [Google Scholar] [CrossRef]
- Correia, P.; Vítor, A.; Tenreiro, M.; Correia, A.C.; Madanelo, J.; Guiné, R. Effect of different thistle flower ecotypes as milk-clotting in Serra da Estrela cheese. Nutr. Food Sci. 2016, 46, 458–475. [Google Scholar] [CrossRef]
- Gomes, S.; Belo, A.T.; Alvarenga, N.; Dias, J.; Lage, P.; Pinheiro, C.; Pinto-Cruz, C.; Brás, T.; Duarte, M.F.; Martins, A.P.L. Characterization of Cynara cardunculus L. flower from Alentejo as a coagulant agent for cheesemaking. Int. Dairy J. 2019, 91, 178–184. [Google Scholar] [CrossRef]
- Louro Martins, A.P.; Pestana De Vasconcelos, M.M.; De Sousa, R.B. Thistle (Cynara cardunculus L.) flower as a coagulant agent for cheesemaking. Short characterization. Lait 1996, 76, 473–477. [Google Scholar] [CrossRef]
- Cordeiro, M.C.; Pais, M.S.; Brodelius, P.E. Tissue-specific expression of multiple forms of cyprosin (aspartic proteinase) in flowers of Cynara cardunculus. Physiol. Plant. 1994, 92, 645–653. [Google Scholar] [CrossRef]
- Veríssimo, P.; Esteves, C.; Faro, C.; Pires, E. The vegetable rennet of Cynara cardunculus L. contains two proteinases with chymosin and pepsin-like specificities. Biotechnol. Lett. 1995, 17, 621–626. [Google Scholar] [CrossRef]
- Cordeiro, M.C.; Xue, Z.T.; Pietrzak, M.; Salomé Pais, M.; Brodelius, P.E. Isolation and characterization of a cDNA from flowers of Cynara cardunculus encoding cyprosin (an aspartic proteinase) and its use to study the organ-specific expression of cyprosin. Plant Mol. Biol. 1994, 24, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Veríssimo, P.; Faro, C.; Moir, A.J.G.; Lin, Y.; Tang, J.; Pires, E. Purification, characterization and partial amino acid sequencing of two new aspartic proteinases from fresh flowers of Cynara cardunculus L. Eur. J. Biochem. 1996, 235, 762–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmento, A.C.; Lopes, H.; Oliveira, C.S.; Vitorino, R.; Samyn, B.; Sergeant, K.; Debyser, G.; Van Beeumen, J.; Domingues, P.; Amado, F.; et al. Multiplicity of aspartic proteinases from Cynara cardunculus L. Planta 2009, 230, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, C.; Van Der Straeten, D.; Pires, E.; Faro, C.; Rodrigues-Pousada, C. Characterization and expression analysis of the aspartic protease gene family of Cynara cardunculus L. FEBS J. 2007, 274, 2523–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waugh, D.F.; von Hippel, P.H. κ-Casein and the stabilization of casein micelles. J. Am. Chem. Soc. 1956, 78, 4576–4582. [Google Scholar] [CrossRef]
- Walstra, P. The Syneresis of Curd. In Cheese: Chemistry, Physics and Microbiology; Fox, P.F., Ed.; Springer US: Boston, MA, USA, 1993; pp. 141–191. ISBN 978-1-4615-2650-6. [Google Scholar]
- Delfour, A.; Jollès, J.; Alais, C.; Jollès, P. Caseino-glycopeptides: Characterization of a methionine residue and of the N-terminal sequence. Biochem. Biophys. Res. Commun. 1965, 19, 452–455. [Google Scholar] [CrossRef]
- Macedo, I.Q.; Faro, C.J.; Pires, E.M. Specificity and kinetics of the milk-clotting enzyme from cardoon (Cynara cardunculus L.) toward bovine kappa-casein. J. Agric. Food Chem. 1993, 41, 1537–1540. [Google Scholar] [CrossRef]
- Silva, V.S.; Xavier Malcata, F. Proteolysis of ovine caseins by cardosin A, an aspartic acid proteinase from Cynara cardunculus L. Lait 1998, 78, 513–519. [Google Scholar] [CrossRef]
- Silva, S.V.; Malcata, F.X. On the activity and specificity of cardosin B, a plant proteinase, on ovine caseins. Food Chem. 1999, 67, 373–378. [Google Scholar] [CrossRef]
- Silva, S.V.; Allmere, T.; Malcata, F.X.; Andrén, A. Comparative studies on the gelling properties of cardosins extracted from Cynara cardunculus and chymosin on cow’s skim milk. Int. Dairy J. 2003, 13, 559–564. [Google Scholar] [CrossRef]
- Agboola, S.; Chen, S.; Zhao, J. Formation of bitter peptides during ripening of ovine milk cheese made with different coagulants. Lait 2004, 84, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.; Pissarra, J.; Veríssimo, P.; Castanheira, P.; Costa, Y.; Pires, E.; Faro, C. Molecular cloning and characterization of cDNA encoding cardosin B, an aspartic proteinase accumulating extracellularly in the transmitting tissue of Cynara cardunculus L. Plant Mol. Biol. 2001, 45, 529–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho-Santos, M.; Pissarra, J.; Veríssimo, P.; Pereira, S.; Salema, R.; Pires, E.; Faro, C.J. Cardosin A, an abundant aspartic proteinase, accumulates in protein storage vacuoles in the stigmatic papillae of Cynara cardunculus L. Planta 1997, 203, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Da Costa, D.S.; Pereira, S.; De Moura Nogueira, F.; Albuquerque, P.M.; Teixeira, J.; Faro, C.; Pissarra, J. Cardosins in postembryonic development of cardoon: Towards an elucidation of the biological function of plant aspartic proteinases. Protoplasma 2008, 232, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.; Pereira, C.; da Costa, D.S.; Teixeira, J.; Fidalgo, F.; Pereira, S.; Pissarra, J. Characterization of aspartic proteinases in C. cardunculus L. callus tissue for its prospective transformation. Plant Sci. 2010, 178, 140–146. [Google Scholar] [CrossRef]
- Duarte, P.; Pissarra, J.; Moore, I. Processing and trafficking of a single isoform of the aspartic proteinase cardosin a on the vacuolar pathway. Planta 2008, 227, 1255–1268. [Google Scholar] [CrossRef]
- Da Costa, D.S.; Pereira, S.; Moore, I.; Pissarra, J. Dissecting cardosin B trafficking pathways in heterologous systems. Planta 2010, 232, 1517–1530. [Google Scholar] [CrossRef]
- Grosshans, B.L.; Ortiz, D.; Novick, P. Rabs and their effectors: Achieving specificity in membrane traffic. Proc. Natl. Acad. Sci. USA 2006, 103, 11821–11827. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.; Ashford, D.A.; Nimtz, M.; Bento, I.; Frazäo, C.; Esteves, C.L.; Faro, C.J.; Kervinen, J.; Pires, E.; Veríssimo, P.; et al. The glycosylation of the aspartic proteinases from barley (Hordeum vulgare L.) and cardoon (Cynara cardunculus L.). Eur. J. Biochem. 1997, 243, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Törmäkangas, K.; Hadlington, J.L.; Pimpl, P.; Hillmer, S.; Brandizzi, F.; Teeri, T.H.; Denecke, J. A vacuolar sorting domain may also influence the way in which proteins leave the endoplasmic reticulum. Plant Cell 2001, 13, 2021–2032. [Google Scholar] [CrossRef] [Green Version]
- Terauchi, K.; Asakura, T.; Ueda, H.; Tamura, T.; Tamura, K.; Matsumoto, I.; Misaka, T.; Hara-Nishimura, I.; Abe, K. Plant-specific insertions in the soybean aspartic proteinases, soyAP1 and soyAP2, perform different functions of vacuolar targeting. J. Plant Physiol. 2006, 163, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.M.; Gomes, D.; Faro, C.; Simões, I. Engineering a cardosin B-derived rennet for sheep and goat cheese manufacture. Appl. Microbiol. Biotechnol. 2014, 99, 269–281. [Google Scholar] [CrossRef]
- Almeida, C.M.; Manso, J.A.; Figueiredo, A.C.; Antunes, L.; Cruz, R.; Manadas, B.; Bur, D.; Pereira, P.J.B.; Faro, C.; Simões, I. Functional and structural characterization of synthetic cardosin B-derived rennet. Appl. Microbiol. Biotechnol. 2017, 101, 6951–6968. [Google Scholar] [CrossRef] [PubMed]
- White, P.C.; Cordeiro, M.C.; Arnold, D.; Brodelius, P.E.; John, K. Processing, activity, and inhibition of recombinant cyprosin, an aspartic proteinase from cardoon (Cynara cardunculus). J. Biol. Chem. 1999, 274, 16685–16693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, P.N.; Fortes, A.M.; Cabral, J.M.S.; Pais, M.S.; Fonseca, L.P. Production and characterization of recombinant cyprosin B in Saccharomyces cerevisiae (W303-1A) strain. J. Biosci. Bioeng. 2008, 105, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, P.N.; Neto, H.; Poejo, P.; Serrazina, S.M.T.; Pais, M.S.S. Overexpression and characterization of cyprosin B in transformed suspension cells of Cynara cardunculus. Plant Cell Tissue Organ Cult. 2010, 101, 311–321. [Google Scholar] [CrossRef]
- Feijoo-Siota, L.; Rama, J.L.R.; Sánchez-Pérez, A.; Villa, T.G. Expression, activation and processing of a novel plant milk-clotting aspartic protease in Pichia pastoris. J. Biotechnol. 2018, 268, 28–39. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Fevereiro, P.; Cabral, J.M.S.; Novais, J.M.; Salomé, M.; Pais, S. Callus and suspension cultures for biomass production of Cynara cardunculus (Compositae). Biotechnol. Lett. 1987, 9, 213–218. [Google Scholar] [CrossRef]
- Lima Costa, M.E.; Van Gulik, W.M.; Ten Hoopen, J.G.; Pais, M.S.S.; Cabral, J.M.S. Protease and phenol production of Cynara cardunculus L. Cell suspension in a chemostat. Enzyme Microb. Technol. 1996, 19, 493–500. [Google Scholar] [CrossRef]
- Wei, Z.Y.; Zhang, Y.Y.; Wang, Y.P.; Fan, M.X.; Zhong, X.F.; Xu, N.; Lin, F.; Xing, S.C. Production of bioactive recombinant bovine chymosin in tobacco plants. Int. J. Mol. Sci. 2016, 17, 624. [Google Scholar] [CrossRef] [Green Version]
- Folgado, A.; Pires, A.S.; Figueiredo, A.C.; Pimentel, C.; Abranches, R. Toward alternative sources of milk coagulants for cheese manufacturing: Establishment of hairy roots culture and protease characterization from Cynara cardunculus L. Plant Cell Rep. 2020, 39, 89–100. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Thistle AP | Organism | Processing | Reference |
|---|---|---|---|
| Cardosin A | Escherichia coli | Inactive | [11] |
| Cardosin A | Nicotiana tabacum Arabidopsis thaliana | Active | [38] |
| Cardosin B | Kluyveromyces lactis | Inactive | [44,45] |
| Cardosin B | Nicotiana tabacum Arabidopsis thaliana | Active | [39] |
| Cardosin B | Tobacco BY2 cell suspension cultures | Active | Folgado et al. (in preparation) |
| Cyprosin B | Pichia pastoris | Inactive | [46] |
| Cyprosin B | Saccharomyces cerevisiae | Inactive/active | [47] |
| Cyprosin B | Cynara cardunculus L. cell suspension cultures | Active | [48] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Folgado, A.; Abranches, R. Plant Aspartic Proteases for Industrial Applications: Thistle Get Better. Plants 2020, 9, 147. https://doi.org/10.3390/plants9020147
Folgado A, Abranches R. Plant Aspartic Proteases for Industrial Applications: Thistle Get Better. Plants. 2020; 9(2):147. https://doi.org/10.3390/plants9020147
Chicago/Turabian StyleFolgado, André, and Rita Abranches. 2020. "Plant Aspartic Proteases for Industrial Applications: Thistle Get Better" Plants 9, no. 2: 147. https://doi.org/10.3390/plants9020147