Genome-Wide Identification and Expression Analysis of Auxin Response Factor (ARF) Gene Family in Longan (Dimocarpus longan L.)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Identification of ARF Genes in Dimocarpus Longan
2.3. Sequence Alignment and Phylogenetic Tree
2.4. Gene Structure and Domain Analysis of DlARFs
2.5. RNA Isolation and Quantitative Real Time (qRT-PCR)
3. Results
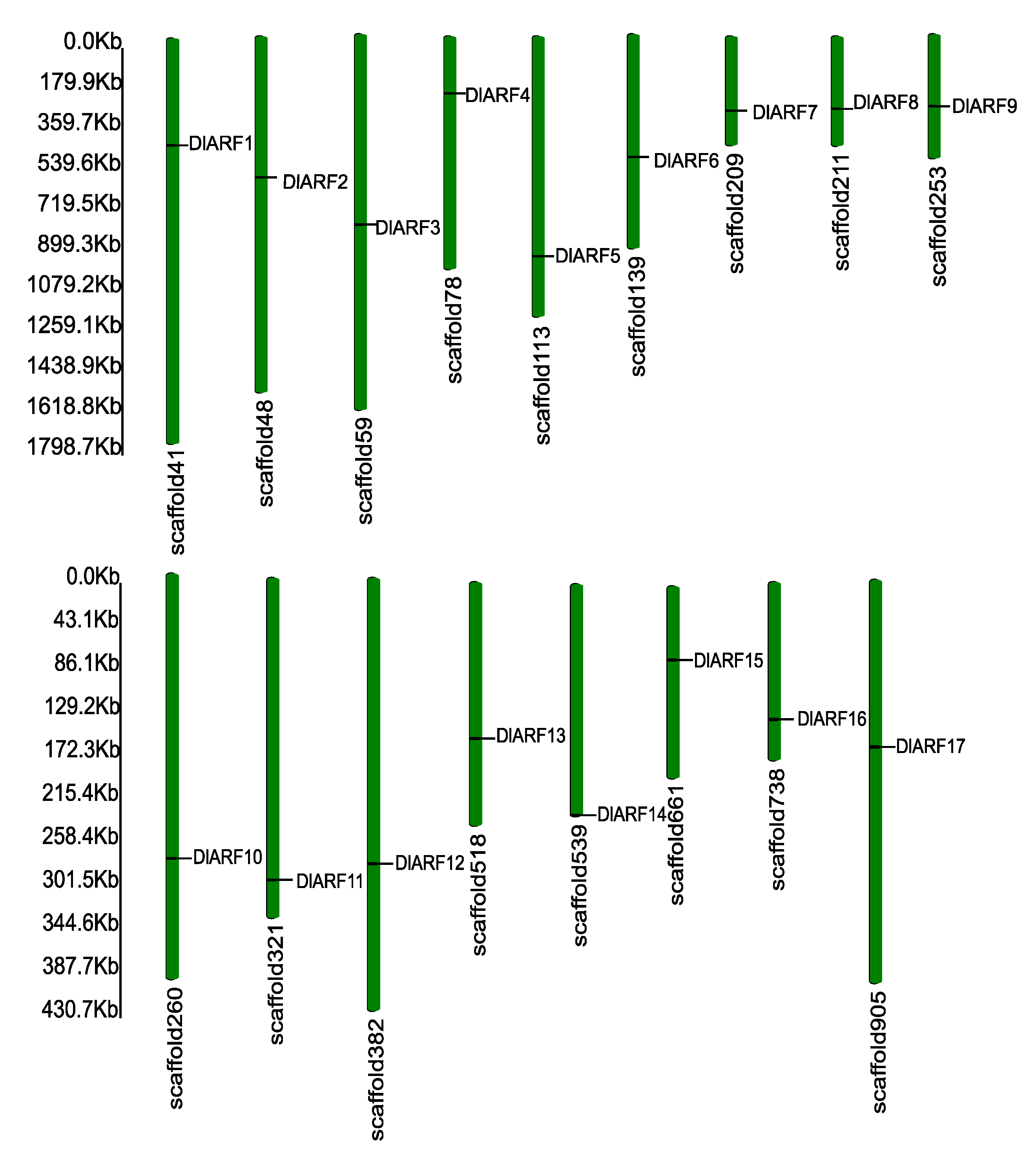
3.1. Identification of DlARF Gene Family in Longan
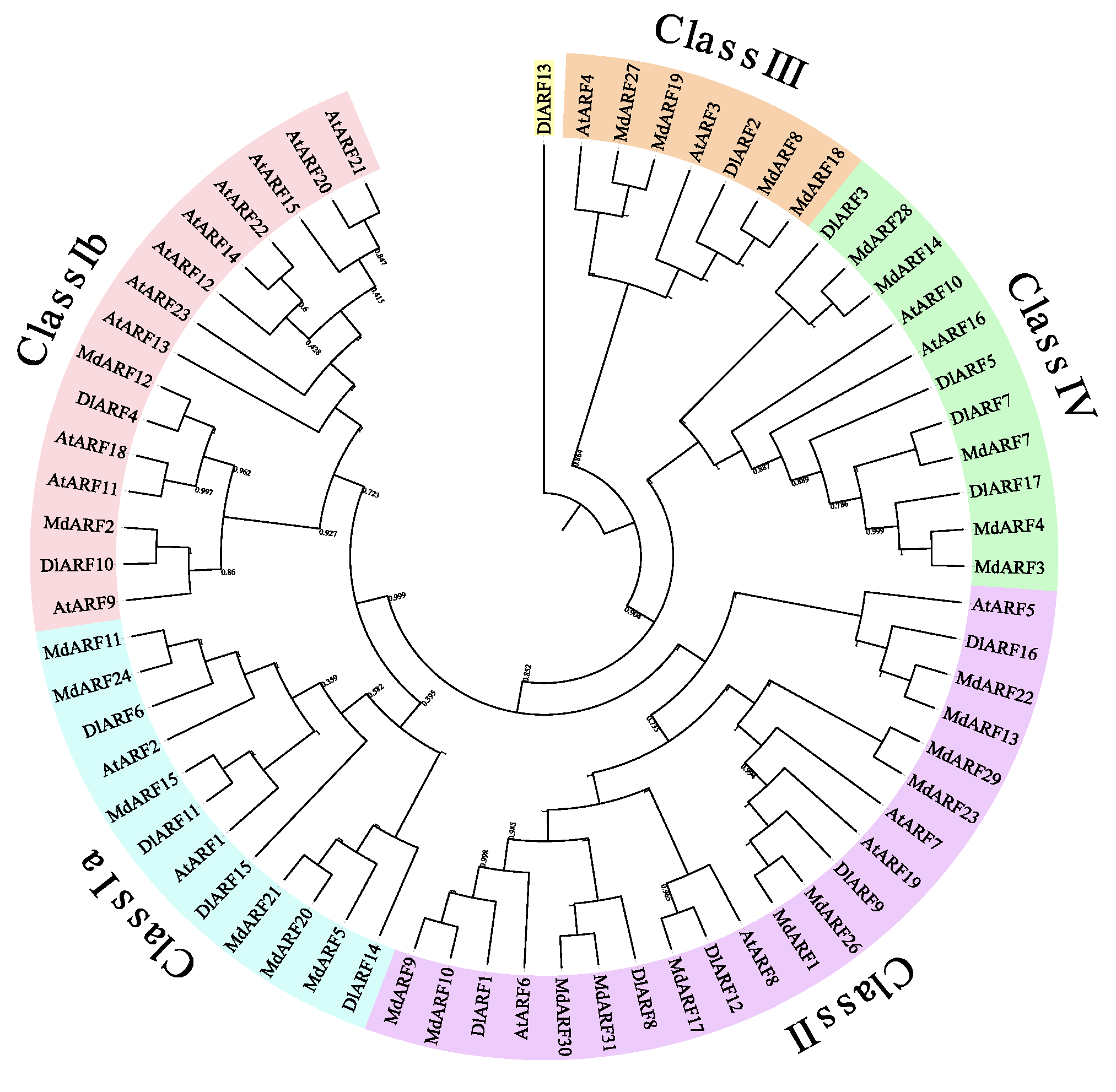
3.2. Phylogenetic Analysis and Classification of DlARF Genes
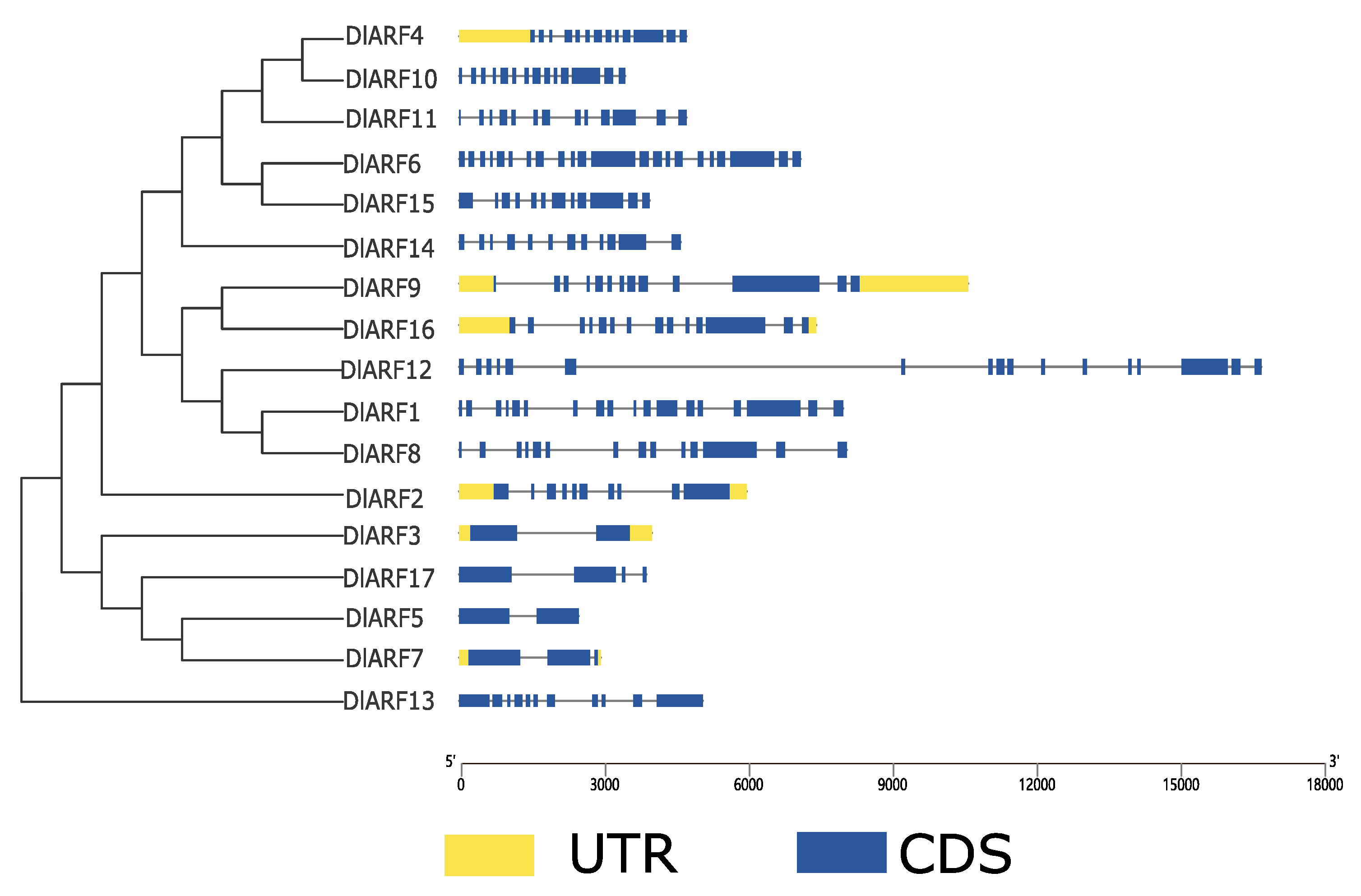
3.3. Structure Analysis and Motif Composition of DlARF Genes
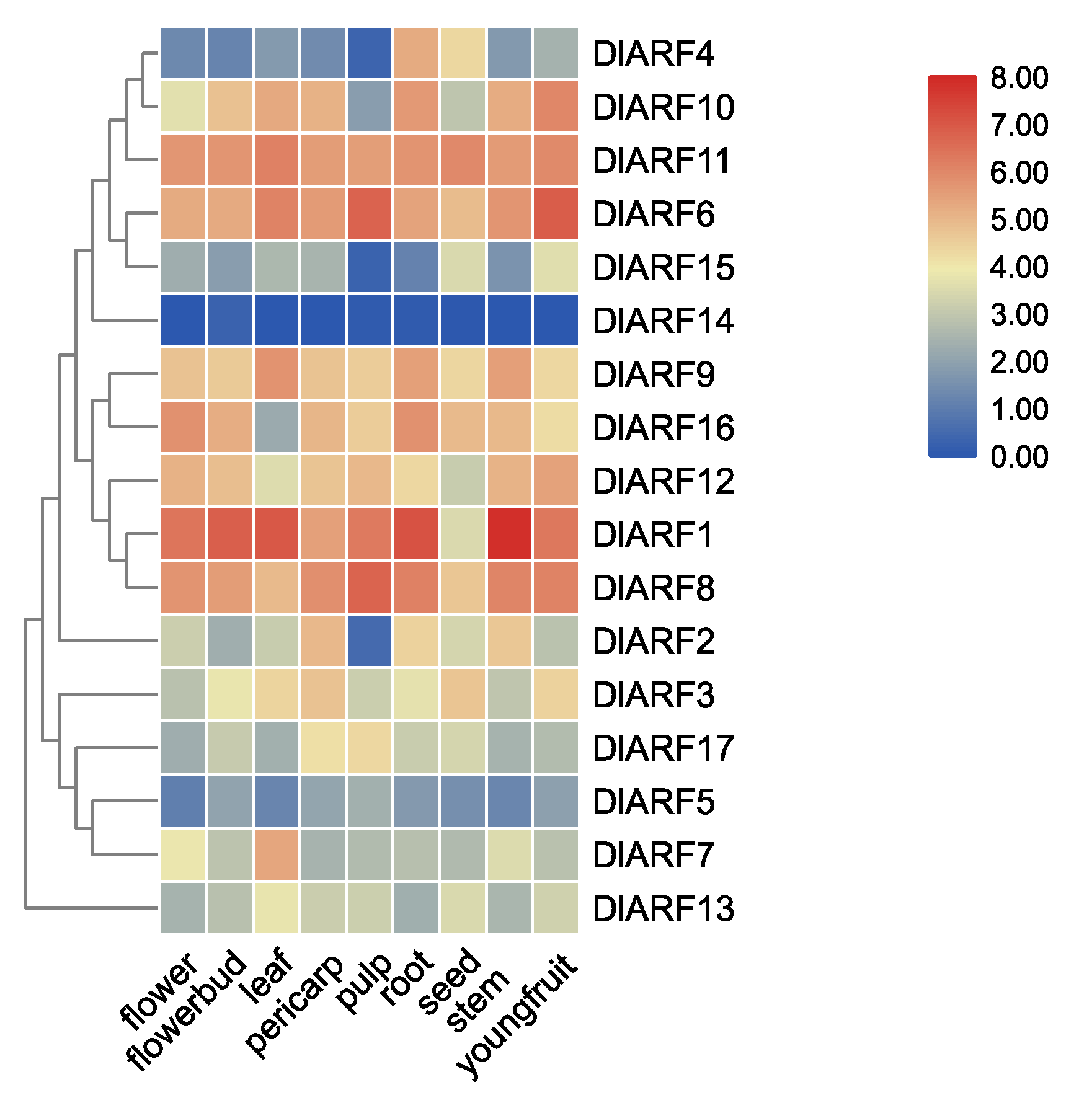
3.4. Expression Profiles of DlARF Genes in Different Plant Tissues
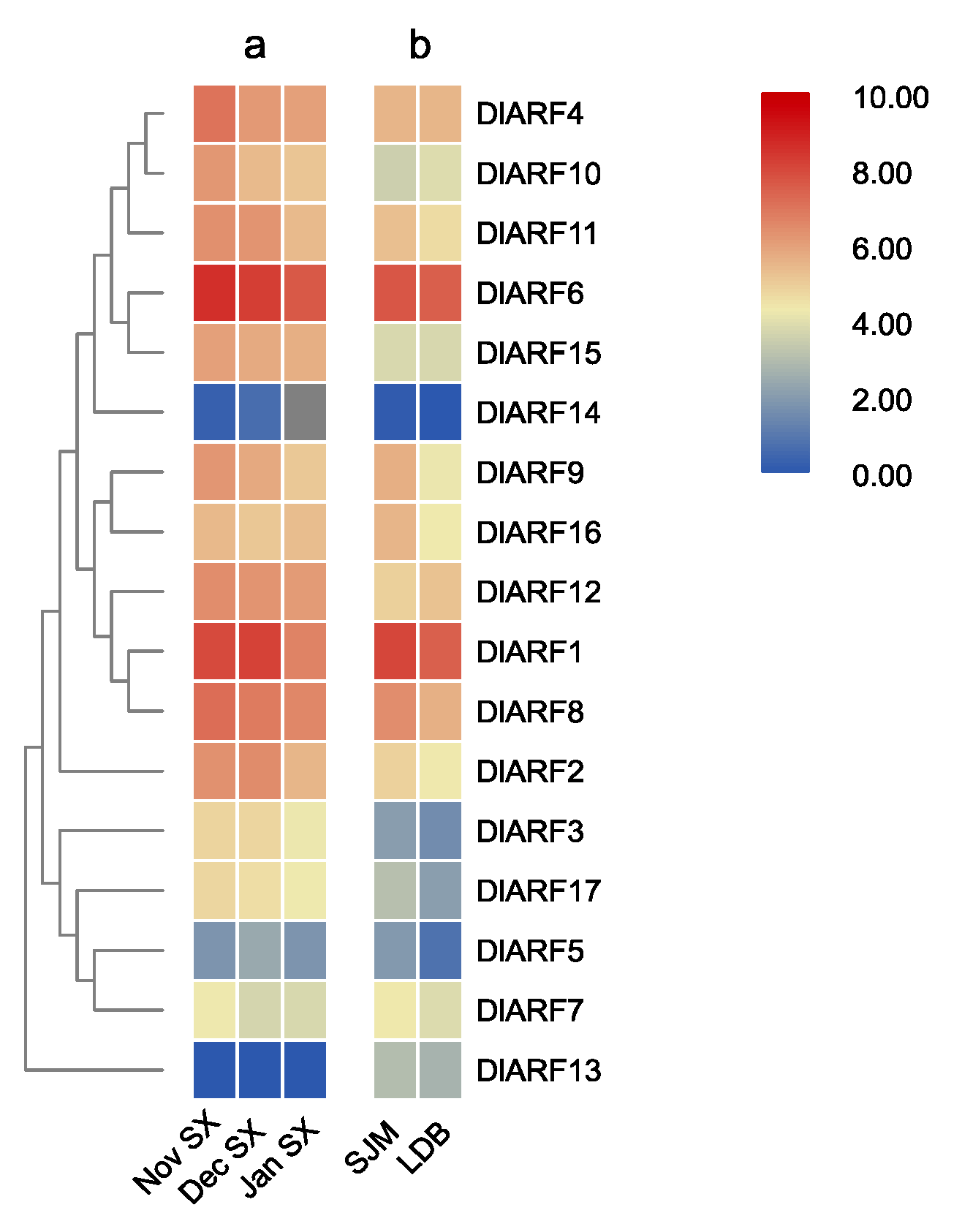
3.5. Expression Profiles of DlARF Genes in Different Longan Varieties
3.6. Expression Patterns of Seven DlARF Genes during Floral Bud Physiological Differentiation of Longan
3.7. Cis-Element Prediction in Promoters of Seven DlARF Genes
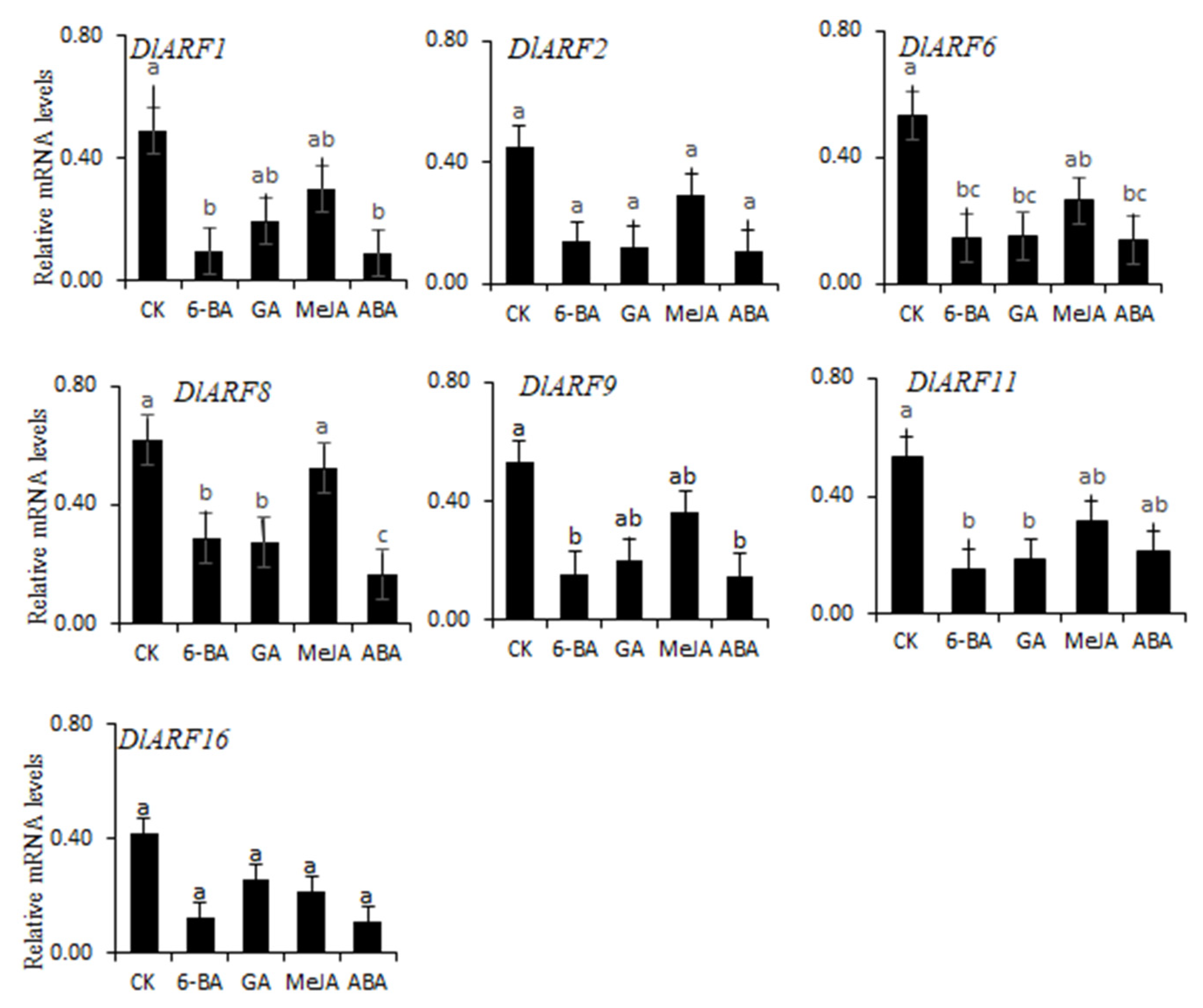
3.8. Expression Patterns of Seven DlARFs with Hormone Treatments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mattsson, J.; Ckurshumova, W.; Berleth, T. Auxin signaling in Arabidopsis leaf vascular development. Plant Physiol. 2003, 131, 1327–1339. [Google Scholar] [CrossRef] [Green Version]
- Phillips, I.D. Apical Dominance. Annu. Rev. Plant Physiol. 2003, 26, 341–367. [Google Scholar] [CrossRef]
- Robert, H.S.; Park, C.; Gutièrrez, C.L.; Wójcikowska, B.; Pěnčík, A.; Novák, O.; Chen, J.; Grunewald, W.; Dresselhaus, T.; Friml, J.; et al. Maternal auxin supply contributes to early embryo patterning in Arabidopsis. Nat. Plants 2018, 4, 548–553. [Google Scholar]
- Friml, J.; Wiśniewska, J.; Benková, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.; Peng, Y.; Zou, C.; Wang, D.; Xu, Z.; Bai, S. A simple treatment to significantly increase signal specificity in immunohistochemistry. Plant Mol. Biol. Rep. 2006, 24, 93–101. [Google Scholar] [CrossRef]
- Ke, M.; Gao, Z.; Chen, J.; Qiu, Y.; Zhang, L.; Chen, X. Auxin controls circadian flower opening and closure in the waterlily. BMC Plant Biol. 2018, 18, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shizufumi, T.; Atsushi, M.; Hiroshi, H. Regulation by Abscisic Acid of In Vitro Flower Formation in Torenia Stem Segments. Plant Cell Physiol. 1985, 26, 675–682. [Google Scholar]
- Ferreiraa, W.M.; Kerbauyb, G.B.; Krausb, J.E.; Pescadorc, R.; Suzuki, R.M. Thidiazuron influences the endogenous levels of cytokinins and IAA during the flowering of isolated shoots of Dendrobium. J. Plant Physiol. 2006, 163, 1126–1134. [Google Scholar] [CrossRef]
- Gretchen, H.; Tom, G. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar]
- Woodward Andrew, W.; Bonnie, B. Auxin: Regulation, Action, and Interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [Green Version]
- Liscum, E.; Reed, J.W. Genetics of Aux/IAA and ARF action in plant growth and development. Plant Mol. Biol. 2002, 49, 387. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, S.; Rammyani, B.; Mark, E. SCFTIR1/AFB-based auxin perception: Mechanism and role in plant growth and development. Plant Cell 2015, 27, 9–19. [Google Scholar]
- Szemenyei, H.; Hannon, M.; Long, J.A. TOPLESS Mediates Auxin-Dependent Transcriptional Repression During Arabidopsis Embryogenesis. Science 2008, 319, 1384–1386. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Estelle, M. Diversity and specificity: Auxin perception and signaling through the TIR1/AFB pathway. Curr. Opin. Plant Biol. 2014, 21, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Guilfoyle, T.J.; Hagen, G. Auxin response factors. Curr. Opin. Plant Biol. 2007, 10, 453–460. [Google Scholar] [CrossRef]
- Song, S.; Hao, L.; Zhao, P.; Zhong, Y.; Xu, N.; Zhang, H.; Liu, N. Genome-wide Identification, Expression Profiling and Evolutionary Analysis of Auxin Response Factor Gene Family in Potato (Solanum tuberosum Group Phureja). Sci. Rep. 2019, 9, 1755. [Google Scholar] [CrossRef]
- Wang, D.; Pei, K.; Fu, Y.; Sun, Z.; Li, S.; Liu, H.; Tang, K.; Han, B.; Tao, Y. Genome-wide analysis of the auxin response factors (ARF) gene family in rice (Oryza sativa). Gene 2007, 394, 13–24. [Google Scholar] [CrossRef]
- Liu, K.; Yuan, C.; Li, H.; Lin, W.; Yang, Y.; Shen, C.; Zheng, X. Genome-wide identification and characterization of auxin response factor (ARF) family genes related to flower and fruit development in papaya (Carica papaya L.). BMC Genom. 2015, 16, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Die, J.V.; Gil, J.; Millan, T. Genome-wide identification of the auxin response factor gene family in Cicer arietinum. BMC Genom. 2018, 19, 301. [Google Scholar] [CrossRef] [Green Version]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.W.; Wu, M.F.; Reeves, P.H.; Hodgens, C.; Yadav, V.; Hayes, S.; Pierik, R. Three Auxin Response Factors Promote Hypocotyl Elongation. Plant Physiol. 2018, 178, 864–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagpal, P.; Ellis, C.M.; Weber, H.; Ploense, S.E.; Barkawi, L.S.; Guilfoyle, T.J.; Hagen, G.; Alonso, J.M.; Cohen, J.D.; Farmer, E.E.; et al. Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development 2005, 132, 132–4107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Li, Z.; Zhao, D. Deregulation of the OsmiR160 Target Gene OsARF18 Causes Growth and Developmental Defects with an Alteration of Auxin Signaling in Rice. Sci. Rep. 2016, 6, 29938. [Google Scholar] [CrossRef] [PubMed]
- Attia, K.A.; Abdelkhalik, A.F.; Ammar, M.H.; Wei, C.; Yang, J.; Lightfoot, D.A.; El-Sayed, W.M.; El-Shemy, H.A. Antisense phenotypes reveal a functional expression of OsARF1, an auxin response factor, in transgenic rice. Curr. Issues Mol. Biol. 2009, 11 (Suppl. 1), i29. [Google Scholar]
- De Jong, M.; Wolters-Arts, M.; Feron, R.; Mariani, C.; Vriezen, W.H. The Solanum lycopersicum auxin response factor 7 (SlARF7) regulates auxin signaling during tomato fruit set and development. Plant J. 2010, 57, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Brian, J.; Pierre, F.; Enrique, O.; Hicham, Z.; Li, Z.; Alain, L.; Claude, P.J.; Mondher, B. Down-regulation of DR12, an auxin response factor homolog, in the tomato results in a pleiotropic phenotype including dark green and blotchy ripening fruit. Plant J. 2002, 32, 603–613. [Google Scholar]
- Li, S.; OuYang, W.; Hou, X.; Xie, L.; Hu, C.; Zhang, J. Genome-wide identification, isolation and expression analysis of auxin response factor (ARF) gene family in sweet orange (Citrus sinensis). Front. Plant Sci. 2015, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Zuo, J.; Hou, X.; Yan, Y.; Wei, Y.; Liu, J.; Li, M.; Xu, B.; Jin, Z. The auxin response factor gene family in banana: Genome-wide identification and expression analyses during development, ripening, and abiotic stress. Front. Plant Sci. 2015, 6, 742. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto Tracie, K. Genes uniquely expressed in vegetative and potassium chlorate induced floral buds of Dimocarpus longan. Plant Sci. 2006, 170, 500–510. [Google Scholar] [CrossRef]
- Park, S.J.; Dong, H.P.; Dong, H.K.; Lee, S.; Yoon, B.H.; Jung, W.Y.; Lee, K.T.; Cheong, J.H.; Ryu, J.H. The memory-enhancing effects of Euphoria longan fruit extract in mice. J. Ethnopharmacol. 2010, 128, 160–165. [Google Scholar] [CrossRef]
- Lin, Y.; Min, J.; Lai, R.; Wu, Z.; Chen, Y.; Yu, L.; Cheng, C.; Jin, Y.; Tian, Q.; Liu, Q.; et al. Genome-wide sequencing of longan (Dimocarpus longan Lour.) provides insights into molecular basis of its polyphenol-rich characteristics. GigaScience 2017, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, T.; Wei, D.; Meng, S.; Allan, A.C.; Zeng, L. Identification of Regulatory Genes Implicated in Continuous Flowering of Longan (Dimocarpus longan L.). PLoS ONE 2014, 9, e114568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jue, D.; Sang, X.; Liu, L.; Shu, B.; Wang, Y.; Liu, C.; Wang, Y.; Xie, J.; Shi, S. Comprehensive analysis of the longan transcriptome reveals distinct regulatory programs during the floral transition. BMC Genom. 2019, 20, 126. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface. bioRxiv 2018. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, H.; Liu, L.; Li, W.; Wei, Y.; Shi, S. Validation of Reference Genes for RT-qPCR Studies of Gene Expression in Preharvest and Postharvest Longan Fruits under Different Experimental Conditions. Front. Plant Sci. 2016, 7, 780. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Su, M.; Liu, Z.; Zhuang, Y. Effects of endogenous hormones in bearing basal shoots of ‘Shuizhang’ longan on floral bud differentiation. J. Trop. Crops 1997, 18, 66–71. [Google Scholar]
- Hou, Z.; Huang, W. Immunohistochemical localization of IAA and ABP1 in strawberry shoot apexes during floral induction. Planta 2005, 222, 678–687. [Google Scholar] [CrossRef]
- Jenik Pablo, D.; Barton, M.K. Surge and destroy: The role of auxin in plant embryogenesis. Development 2005, 132, 3577–3585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyser, O. The fall and rise of apical dominance. Curr. Opin. Genet. Dev. 2005, 15, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Tim, U.; Jane, M.; Gretchen, H.; Guilfoyle Tom, J. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 1997, 9, 1963–1971. [Google Scholar]
- Guilfoyle, T.; Hagen, G.; Ulmasov, T.; Murfett, J. How does auxin turn on genes? Plant Physiol. 1998, 118, 341–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piya, S.; Shrestha, S.K.; Binder, B.; Stewart, N.; Hewezi1, T. Protein-protein interaction and gene co-expression maps of ARFs and Aux/IAAs in Arabidopsis. Front. Plant Sci. 2014, 5, 744. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Xie, Z.; Hu, C.; Zhang, J. A Review of Auxin Response Factors (ARFs) in Plants. Front. Plant Sci. 2016, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Tim, U.; Gretchen, H.; Tom, G.J. Dimerization and DNA binding of auxin response factors. Plant J. 1999, 19, 309–319. [Google Scholar]
- Mark, R.; Sébastien, P.; Dolf, W. Auxin Response Factors: Output control in auxin biology. J. Exp. Bot. 2018, 69, 179–188. [Google Scholar]
- Barbara, M.; Colette, A.H.T.; Xiang, D.; Nerys, W.; González, L.L.; Saiko, Y.; Margo, S.; Raju, D.; Dolf, W. Auxin response cell-autonomously controls ground tissue initiation in the early Arabidopsis embryo. Proc. Natl. Acad. Sci. USA 2017, 114, E2533–E2539. [Google Scholar]
- Ding, S.F.; Chen, W.S.; Su, C.; Du, B.S.; Twitchin, B.; Bhaskar, V.K. Changes in free and conjugated indole-3-acetic acid during early stage of flower bud differentiation in Polianthes tuberosa. Plant Physiol. Biochem. 1999, 37, 161–165. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, J.; Wang, X.; Shi, X.; Wang, B.; Zheng, X.C.; Liu, F. The Influence of Changes of Endogenous Hormones in Shoot on the Grapes Flower Bud Differentiation in Greenhouse. Scientia Agricultura Sinica 2014, 134, 1065–1071. [Google Scholar]
- Yang, Z.; Huang, X.; Chen, Y. Changes of Endogenous Hormone Level during Flower Development in On-season and Off-season Longan. J. Anhui Agric. Sci. 2019, 47, 47–50. [Google Scholar]
- Liu, N.; Wu, S.; van Houten, J.; Wang, Y.; Ding, B.; Fei, Z.; Clarke, T.H.; Reed, J.W.; van der Knaap, E. Down-regulation of AUXIN RESPONSE FACTORS 6 and 8 by microRNA 167 leads to floral development defects and female sterility in tomato. J. Exp. Bot. 2014, 65, 2507–2520. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Dinh, T.T.; Li, D.; Shi, B.; Li, Y.; Cao, X.; Guo, L.; Pan, Y.; Jiao, Y.; Chen, X. AUXIN RESPONSE FACTOR 3 integrates the functions of AGAMOUS and APETALA2 in floral meristem determinacy. Plant J. 2014, 80, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Wang, R.; Zi, H.; Li, Y.; Cao, X.; Li, D.; Guo, L.; Tong, J.; Pan, Y.; Jiao, Y.; et al. AUXIN RESPONSE FACTOR3 Regulates Floral Meristem Determinacy By Repressing Cytokinin Biosynthesis and Signaling. Plant Cell 2018, 30, 705–2017. [Google Scholar] [CrossRef] [Green Version]
- Przemeck, G.K.; Mattsson, J.; Hardtke, C.S.; Sung, Z.R.; Berleth, T. Studies on the role of the Arabidopsis gene MONOPTEROS in vascular development and plant cell axialization. Planta 1996, 200, 229–237. [Google Scholar] [CrossRef]
- Fowler, S.; Lee, K.; Onouchi, H.; Samach, A.; Richardson, K.; Morris, B.; Coupland, G.; Putterill, J. GIGANTEA: A circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains. Embo J. 1999, 18, 4679–4688. [Google Scholar] [CrossRef] [Green Version]
- Galvão, V.C.; Collani, S.; Horrer, D.; Schmid, M. Gibberellic acid signaling is required for ambient temperature-mediated induction of flowering in Arabidopsis thaliana. Plant J. 2015, 84, 949–962. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, N.; Winter, C.M.; Wu, M.; Kanno, Y.; Yamaguchi, A.; Seo, M.; Wagner, D. Gibberellin acts positively then negatively to control onset of flower formation in Arabidopsis. Science 2014, 344, 638–641. [Google Scholar] [CrossRef]
- Tomoko, N.; Takamasa, S.; Yumiko, T.; Rie, I.; Tetsuya, H.; Hitoshi, S.; Sumie, I. Jasmonic acid facilitates flower opening and floral organ development through the upregulated expression of SlMYB21 transcription factor in tomato. Biosci. Biotechnol. Biochem. 2018, 82, 292–303. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Identifier | Exon Numbers | PI | MW (kDa) | Genomic Position | ORF Length (bp) | Homolog in Arabidopsis | MR Domain (Amino Acid Content) |
|---|---|---|---|---|---|---|---|---|
| DlARF1 | Dlo.020240.1 | 18 | 6.66 | 132.00 | scaffold41:473073-481372 | 3558 | AtARF6 | QSL |
| DlARF2 | Dlo.022331.1 | 18 | 6.61 | 80.87 | scaffold48:620922-626016 | 2238 | AtARF3 | GLS |
| DlARF3 | Dlo.025765.1 | 2 | 5.76 | 63.54 | scaffold59:841815-845259 | 1728 | AtARF10 | PSV |
| DlARF4 | Dlo.030332.1 | 13 | 6.83 | 76.01 | scaffold78:251174-254549 | 2043 | AtARF11.18 | LSV |
| DlARF5 | Dlo.002610.1 | 2 | 6.48 | 72.72 | scaffold113:973236-975819 | 1986 | AtARF16 | GLS |
| DlARF6 | Dl0.005415.2 | 22 | 6.15 | 166.68 | scaffold139:542989-550375 | 4497 | AtARF2 | EPS |
| DlARF7 | Dlo.011077.1 | 2 | 6.58 | 77.10 | scaffold209:327655-330450 | 2100 | AtARF16 | QSL |
| DlARF8 | Dlo.011327.1 | 14 | 5.89 | 100.03 | scaffold211:316240-324625 | 2718 | AtARF6 | QSL |
| DlARF9 | Dlo.013714.2 | 13 | 6.10 | 126.04 | scaffold253:313870-321781 | 3393 | AtARF19 | QSL |
| DlARF10 | Dlo.014083.1 | 14 | 6.05 | 77.38 | scaffold260:282418-286013 | 2079 | AtARF9 | LSV |
| DlARF11 | Dlo.016820.1 | 12 | 5.96 | 69.86 | scaffold321:299097-304013 | 1878 | AtARF1 | PSV |
| DlARF12 | Dlo.019200.4 | 17 | 6.22 | 106.61 | scaffold382:283090-300432 | 2850 | AtARF8 | QSL |
| DlARF13 | Dlo.023607.1 | 11 | 5.99 | 104.31 | scaffold518:150916-156184 | 2793 | LSV | |
| DlARF14 | Dlo.024143.1 | 12 | 8.03 | 70.51 | scaffold539:226240:231034 | 1902 | GLS | |
| DlARF15 | Dlo.027785.3 | 12 | 6.87 | 88.20 | scaffold661:72501:76620 | 2376 | AtARF2 | LSV |
| DlARF16 | Dlo.029636.1 | 14 | 5.27 | 104.34 | scaffold738:134978:141446 | 2829 | AtARF5 | QSL |
| DlARF17 | Dlo.032913.1 | 4 | 7.21 | 79.38 | scaffold905:165555-169610 | 2181 | AtARF16 | GLS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Y.; Fang, T.; Zhang, Y.; Zhang, M.; Zeng, L. Genome-Wide Identification and Expression Analysis of Auxin Response Factor (ARF) Gene Family in Longan (Dimocarpus longan L.). Plants 2020, 9, 221. https://doi.org/10.3390/plants9020221
Peng Y, Fang T, Zhang Y, Zhang M, Zeng L. Genome-Wide Identification and Expression Analysis of Auxin Response Factor (ARF) Gene Family in Longan (Dimocarpus longan L.). Plants. 2020; 9(2):221. https://doi.org/10.3390/plants9020221
Chicago/Turabian StylePeng, Yuan, Ting Fang, Yiyong Zhang, Mengyuan Zhang, and Lihui Zeng. 2020. "Genome-Wide Identification and Expression Analysis of Auxin Response Factor (ARF) Gene Family in Longan (Dimocarpus longan L.)" Plants 9, no. 2: 221. https://doi.org/10.3390/plants9020221
APA StylePeng, Y., Fang, T., Zhang, Y., Zhang, M., & Zeng, L. (2020). Genome-Wide Identification and Expression Analysis of Auxin Response Factor (ARF) Gene Family in Longan (Dimocarpus longan L.). Plants, 9(2), 221. https://doi.org/10.3390/plants9020221