High-Resolution Mapping of the Novel Early Leaf Senescence Gene Els2 in Common Wheat

Abstract
:1. Introduction
2. Results
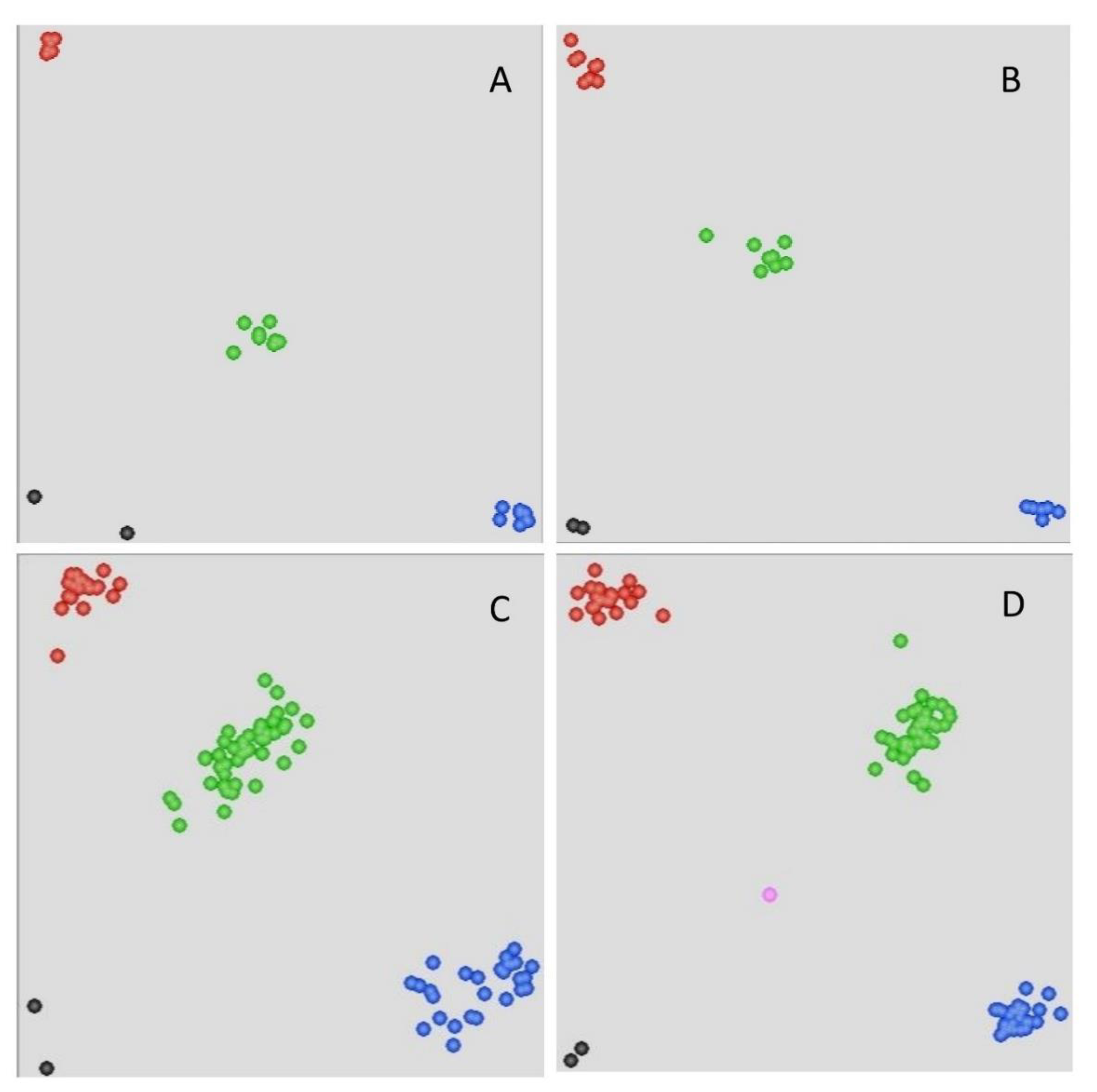
2.1. Development of Kompetitive Allele-Specific Polymerase Chain Reaction (KASP) Markers Based on the 90K Single-Nucleotide Polymorphisms (SNP) Array
2.2. Development of KASP Markers Based on the 660K SNP Array
2.3. Construction of a High-Resolution Genetic Linkage Map
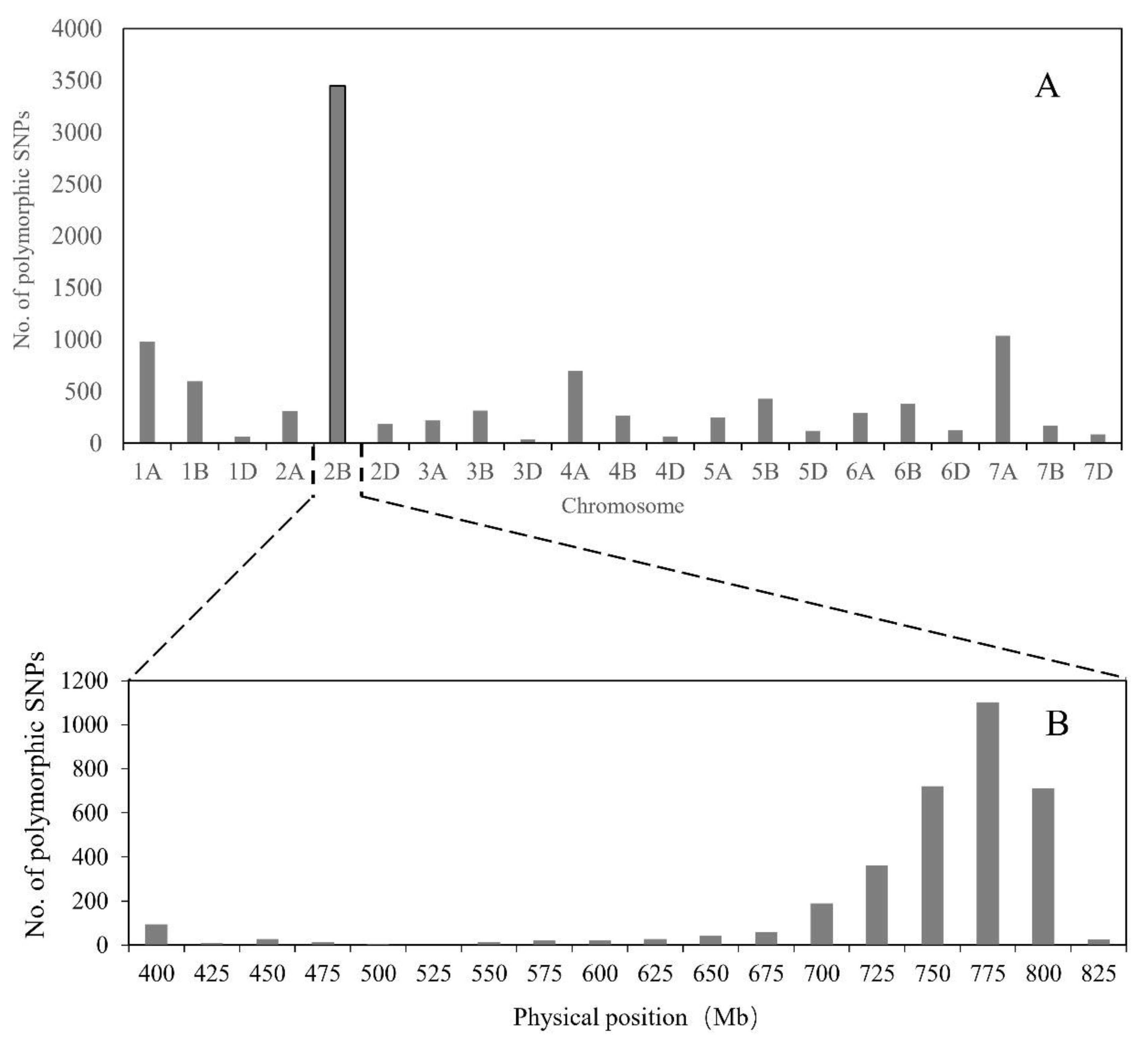
2.4. Physical Mapping and Gene Annotation of the Els2 Goal Interval
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Population Construction
4.2. Combined Bulked Segregant Analysis Using the 660K SNP Array
4.3. Conversion of SNP Markers to KASP Markers
4.4. Construction of High-Density Genetic Linkage Map
4.5. Physical Mapping and Gene Annotation of the Els2 Region
Author Contributions
Funding
Conflicts of Interest
References
- Fu, D.L.; Uauy, C.; Distelfeld, A.; Blechl, A.; Epstein, L.; Chen, X.M.; Sela, H.A.; Fahima, T.; Dubcovsky, J. A Kinase-START Gene Confers Temperature-Dependent Resistance to Wheat Stripe Rust. Science 2009, 323, 1357–1360. [Google Scholar] [CrossRef] [Green Version]
- Sylvester-Bradley, R.; Scott, R.; Wright, C. Physiology in the production and improvement of cereals. In Physiology in the Production and Improvement of Cereals; HGCA: London, UK, 1990. [Google Scholar]
- Li, M.M.; Li, B.B.; Guo, G.H.; Chen, Y.X.; Xie, J.Z.; Lu, P.; Wu, Q.H.; Zhang, D.Y.; Zhang, H.Z.; Yang, J.; et al. Mapping a leaf senescence gene els1 by BSR-Seq in common wheat. Crop J. 2018, 6, 236–243. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, H.J.; Lim, P.O.; Nam, H.G. Leaf Senescence: Systems and Dynamics Aspects. Annu. Rev. Plant Biol. 2019, 70, 347–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Woo, H.R.; Nam, H.G. Toward Systems Understanding of Leaf Senescence: An Integrated Multi-Omics Perspective on Leaf Senescence Research. Mol. Plant 2016, 9, 813–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattanayak, G.K.; Tripathy, B.C. Modulation of biosynthesis of photosynthetic pigments and light-harvesting complex in wild-type and gun5 mutant of Arabidopsis thaliana during impaired chloroplast development. Protoplasma 2016, 253, 747–752. [Google Scholar] [CrossRef]
- Morita, R.; Sato, Y.; Masuda, Y.; Nishimura, M.; Kusaba, M. Defect in non-yellow coloring 3, an alpha/beta hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J. 2009, 59, 940–952. [Google Scholar] [CrossRef]
- Rong, H.; Tang, Y.; Zhang, H.; Wu, P.; Chen, Y.; Li, M.; Wu, G.; Jiang, H. The Stay-Green Rice like (SGRL) gene regulates chlorophyll degradation in rice. J. Plant Physiol. 2013, 170, 1367–1373. [Google Scholar] [CrossRef]
- Yamatani, H.; Sato, Y.; Masuda, Y.; Kato, Y.; Morita, R.; Fukunaga, K.; Nagamura, Y.; Nishimura, M.; Sakamoto, W.; Tanaka, A.; et al. NYC4, the rice ortholog of Arabidopsis THF1, is involved in the degradation of chlorophyll-protein complexes during leaf senescence. Plant J. 2013, 74, 652–662. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, T.; Peng, H.; Luo, S.; Tan, J.; Jiang, K.; Heng, Y.; Zhang, X.; Guo, X.; Zheng, J.; et al. Rice premature leaf senescence 2, encoding a glycosyltransferase (GT), is involved in leaf senescence. Front. Plant Sci. 2018, 9, 560. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Hong, X.; Hu, D.; Liu, C.; Yang, J.; Li, Y.; Huang, Y.; Feng, Y.; Gong, H.; et al. Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) induces early leaf senescence and defence responses in rice. J. Exp. Bot. 2015, 66, 973–987. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.J.; Wuriyanghan, H.; Zhang, Y.Q.; Duan, K.X.; Chen, H.W.; Li, Q.T.; Lu, X.; He, S.J.; Ma, B.; Zhang, W.K.; et al. An S-domain receptor-like kinase, OsSIK2, confers abiotic stress tolerance and delays dark-induced leaf senescence in rice. Plant Physiol. 2013, 163, 1752–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, Z.; Zhang, Y.; Wu, W.; Zhan, X.; Yu, N.; Xu, T.; Liu, Q.; Li, Z.; Shen, X.; Chen, D.; et al. ES7, encoding a ferredoxin-dependent glutamate synthase, functions in nitrogen metabolism and impacts leaf senescence in rice. Plant Sci. 2017, 259, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Ueda, N.; Aoki, K.; Kuromori, T.; Hirayama, T.; Shinozaki, K.; Yamaya, T.; Sakakibara, H. AtIPT3 is a key determinant of nitrate-dependent cytokinin biosynthesis in Arabidopsis. Plant Cell Physiol. 2004, 45, 1053–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Turano, F.J. The putative glutamate receptor 1.1 (AtGLR1.1) functions as a regulator of carbon and nitrogen metabolism in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6872–6877. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y.; et al. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takasaki, H.; Maruyama, K.; Takahashi, F.; Fujita, M.; Yoshida, T.; Nakashima, K.; Myouga, F.; Toyooka, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. SNAC-As, stress-responsive NAC transcription factors, mediate ABA-inducible leaf senescence. Plant J. 2015, 84, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Wang, Y.; Zhu, Y.; Tang, J.; Hu, B.; Liu, L.; Ou, S.; Wu, H.; Sun, X.; Chu, J.; et al. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 10013–10018. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Zhang, Y.X.; Di, C.; Zhang, Q.L.; Zhang, K.; Wang, C.C.; You, Q.; Yan, H.; Dai, S.Y.; Yuan, J.S.; et al. JAZ7 negatively regulates dark-induced leaf senescence in Arabidopsis. J. Exp. Bot. 2016, 67, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.C.; Hong, S.W.; Whang, S.S.; Lim, P.O.; Nam, H.G.; Koo, J.C. Age-Dependent Action of an ABA-Inducible Receptor Kinase, RPK1, as a Positive Regulator of Senescence in Arabidopsis Leaves. Plant Cell Physiol. 2011, 52, 651–662. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Xia, C.; Zhang, L.; Dong, C.; Liu, X.; Kong, X. Transcriptome Analysis of a Premature Leaf Senescence Mutant of Common Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2018, 19, 782. [Google Scholar] [CrossRef] [Green Version]
- Borrill, P.; Harrington, S.A.; Simmonds, J.; Uauy, C. Identification of Transcription Factors Regulating Senescence in Wheat through Gene Regulatory Network Modelling. Plant Physiol. 2019, 180, 1740–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregersen, P.L.; Holm, P.B. Transcriptome analysis of senescence in the flag leaf of wheat (Triticum aestivum L.). Plant Biotechnol. J. 2007, 5, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, P.L.; Holm, P.B.; Krupinska, K. Leaf senescence and nutrient remobilisation in barley and wheat. Plant Biol. 2008, 10, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Jaiswal, V.; Kumar, S.; Balyan, H.S.; Gupta, P.K. Chromosome mapping of four novel mutants in bread wheat (Triticum aestivum L.). Acta Physiol. Plant 2015, 37, 66. [Google Scholar] [CrossRef]
- Harrington, S.A.; Cobo, N.; Karafiatova, M.; Dolezel, J.; Borrill, P.; Uauy, C. Identification of a Dominant Chlorosis Phenotype Through a Forward Screen of the Triticum turgidum cv. Kronos TILLING Population. Front. Plant Sci. 2019, 10, 963. [Google Scholar] [CrossRef] [Green Version]
- Ansari, M.J.; Al-Ghamdi, A.; Usmani, S.; Kumar, R.; Nuru, A.; Singh, K.; Dhaliwal, H.S. Characterization and gene mapping of a brittle culm mutant of diploid wheat (Triticum monococcum L.) with irregular xylem vessels development. Acta Physiol. Plant 2013, 35, 2407–2419. [Google Scholar] [CrossRef]
- Li, N.; Jia, J.; Xia, C.; Liu, X.; Kong, X. Characterization and mapping of novel chlorophyll deficient mutant genes in durum wheat. Breed Sci. 2013, 63, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Bian, N.; Wen, M.; Xiao, J.; Yuan, C.; Cao, A.; Zhang, S.; Wang, X.; Wang, H. Characterization of a common wheat (Triticum aestivum L.) high-tillering dwarf mutant. Theor. Appl. Genet. 2017, 130, 483–494. [Google Scholar] [CrossRef]
- Ansari, M.J.; Al-Ghamdi, A.; Kumar, R.; Usmani, S.; Al-Attal, Y.; Nuru, A.; Mohamed, A.A.; Singh, K.; Dhaliwal, H.S. Characterization and gene mapping of a chlorophyll-deficient mutant clm1 of Triticum monococcum L. Biol. Plant. 2013, 57, 442–448. [Google Scholar] [CrossRef]
- Zhang, L.L.; Liu, C.; An, X.Y.; Wu, H.Y.; Feng, Y.; Wang, H.; Sun, D.J. Identification and genetic mapping of a novel incompletely dominant yellow leaf color gene, Y1718, on chromosome 2BS in wheat. Euphytica 2017, 213, 141. [Google Scholar] [CrossRef]
- Brenchley, R.; Spannagl, M.; Pfeifer, M.; Barker, G.L.; D’Amore, R.; Allen, A.M.; McKenzie, N.; Kramer, M.; Kerhornou, A.; Bolser, D.; et al. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature 2012, 491, 705–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.C.; Gu, Y.Q.; Puiu, D.; Wang, H.; Twardziok, S.O.; Deal, K.R.; Huo, N.; Zhu, T.; Wang, L.; Wang, Y.; et al. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 2017, 551, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Zhang, N.; Fan, X.L.; Zhang, W.; Zhao, C.H.; Yang, L.J.; Pan, R.Q.; Chen, M.; Han, J.; Zhao, X.Q.; et al. Utilization of a Wheat660K SNP array-derived high-density genetic map for high-resolution mapping of a major QTL for kernel number. Sci. Rep. 2017, 7, 3788. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Zhao, S.; Kong, X.; Li, Y.; Zhao, G.; He, W.; Appels, R.; Pfeifer, M.; Tao, Y.; Zhang, X.; et al. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature 2013, 496, 91–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, K.; Duran, C.; Berkman, P.J.; Lorenc, M.T.; Stiller, J.; Manoli, S.; Hayden, M.J.; Forrest, K.L.; Fleury, D.; Baumann, U.; et al. Single nucleotide polymorphism discovery from wheat next-generation sequence data. Plant Biotechnol. J. 2012, 10, 743–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Winfield, M.O.; Allen, A.M.; Burridge, A.J.; Barker, G.L.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G.; et al. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef]
- Klindworth, D.L.; Saini, J.; Long, Y.; Rouse, M.N.; Faris, J.D.; Jin, Y.; Xu, S.S. Physical mapping of DNA markers linked to stem rust resistance gene Sr47 in durum wheat. Theor. Appl. Genet. 2017, 130, 1135–1154. [Google Scholar] [CrossRef]
- Ren, T.; Hu, Y.; Tang, Y.; Li, C.; Yan, B.; Ren, Z.; Tan, F.; Tang, Z.; Fu, S.; Li, Z. Utilization of a Wheat55K SNP Array for Mapping of Major QTL for Temporal Expression of the Tiller Number. Front. Plant Sci. 2018, 9, 333. [Google Scholar] [CrossRef]
- Sun, C.; Dong, Z.; Zhao, L.; Ren, Y.; Zhang, N.; Chen, F. The Wheat 660K SNP array demonstrates great potential for marker-assisted selection in polyploid wheat. Plant Biotechnol. J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yang, W.; Li, Y.; Shan, Q.; Ye, X.; Wang, D.; Yu, K.; Lu, W.; Xin, P.; Pei, Z.; et al. A wheat dominant dwarfing line with Rht12, which reduces stem cell length and affects gibberellic acid synthesis, is a 5AL terminal deletion line. Plant J. 2019, 97, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wen, W.; Xie, L.; Fu, L.; Xu, D.; Fu, C.; Wang, D.; Chen, X.; Xia, X.; Chen, Q.; et al. Molecular Mapping of Reduced Plant Height Gene Rht24 in Bread Wheat. Front. Plant Sci. 2017, 8, 1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zeng, Q.; Wang, Q.; Liu, S.; Yu, S.; Mu, J.; Huang, S.; Sela, H.; Distelfeld, A.; Huang, L.; et al. SNP-based pool genotyping and haplotype analysis accelerate fine-mapping of the wheat genomic region containing stripe rust resistance gene Yr26. Theor. Appl. Genet. 2018, 131, 1481–1496. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Q.; Xu, L.; Chen, X.; Li, B.; Mu, J.; Zeng, Q.; Huang, L.; Han, D.; Kang, Z. Combining Single Nucleotide Polymorphism Genotyping Array with Bulked Segregant Analysis to Map a Gene Controlling Adult Plant Resistance to Stripe Rust in Wheat Line 03031-1-5 H62. Phytopathology 2017, 108, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, Q.; Liu, S.; Huang, S.; Mu, J.; Zeng, Q.; Huang, L.; Han, D.; Kang, Z. Saturation Mapping of a Major Effect QTL for Stripe Rust Resistance on Wheat Chromosome 2B in Cultivar Napo 63 Using SNP Genotyping Arrays. Front. Plant Sci. 2017, 8, 653. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.B.; Wang, N.; Jian, J.T.; Wang, C.S.; Xie, Y.Z. Rapid mapping of a chlorina mutant gene cn-A1 in hexaploid wheat by bulked segregant analysis and single nucleotide polymorphism genotyping arrays. Crop Pasture Sci. 2019, 70, 827–836. [Google Scholar] [CrossRef]
- Wang, N.; Xie, Y.Z.; Li, Y.Z.; Wu, S.N.; Wei, H.H.; Wang, C.S. Molecular mapping a novel early leaf-senescence gene Els2 in common wheat by SNP genotyping arrays. Crop Pasture Sci. 2020, 71, 356–367. [Google Scholar] [CrossRef]
- Wang, M.; Wang, S.; Xia, G. From genome to gene: A new epoch for wheat research? Trends Plant Sci. 2015, 20, 380–387. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Martinez, S.A.; Shorinola, O.; Conselman, S.; See, D.; Skinner, D.Z.; Uauy, C.; Steber, C.M. Exome sequencing of bulked segregants identified a novel TaMKK3-A allele linked to the wheat ERA8 ABA-hypersensitive germination phenotype. Theor. Appl. Genet. 2020, 133, 719–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.H.; Li, J.T.; Wu, P.P.; Li, Y.H.; Qiu, D.; Qu, Y.F.; Xie, J.Z.; Zhang, H.J.; Yang, L.; Fu, T.T.; et al. Development of SNP, KASP, and SSR Markers by BSR-Seq Technology for Saturation of Genetic Linkage Map and Efficient Detection of Wheat Powdery Mildew Resistance Gene Pm61. Int. J. Mol. Sci. 2019, 20, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Rao, Y.; Liu, H.; Fang, Y.; Dong, G.; Huang, L.; Leng, Y.; Guo, L.; Zhang, G.; Hu, J.; et al. Characterization and fine mapping of an early senescence mutant (es-t) in Oryza sativa L. Chin. Sci. Bull. 2011, 56, 2437–2443. [Google Scholar] [CrossRef] [Green Version]
- Christ, B.; Sussenbacher, I.; Moser, S.; Bichsel, N.; Egert, A.; Muller, T.; Krautler, B.; Hortensteiner, S. Cytochrome P450 CYP89A9 Is Involved in the Formation of Major Chlorophyll Catabolites during Leaf Senescence in Arabidopsis. Plant Cell 2013, 25, 1868–1880. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Mendoza, M.; Velasco-Arroyo, B.; Gonzalez-Melendi, P.; Martinez, M.; Diaz, I. C1A cysteine protease-cystatin interactions in leaf senescence. J. Exp. Bot. 2014, 65, 3825–3833. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.H.; Huang, L.T.; Yap, M.N.; Lee, R.H.; Huang, Y.J.; Cheng, M.C.; Chen, S.C. Molecular characterization of a senescence-associated gene encoding cysteine proteinase and its gene expression during leaf senescence in sweet potato. Plant Cell Physiol. 2002, 43, 984–991. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Cai, Z.; Guo, Y.; Gan, S. An arabidopsis mitogen-activated protein kinase cascade, MKK9-MPK6, plays a role in leaf senescence. Plant Physiol. 2009, 150, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.T.; Doyle, J.L. Isolation of Plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Ramirez-Gonzalez, R.H.; Uauy, C.; Caccamo, M. PolyMarker: A fast polyploid primer design pipeline. Bioinformatics 2015, 31, 2038–2039. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Liu, S.; Wang, Q.; Zeng, Q.; Mu, J.; Huang, S.; Yu, S.; Han, D.; Kang, Z. Rapid identification of an adult plant stripe rust resistance gene in hexaploid wheat by high-throughput SNP array genotyping of pooled extremes. Theor. Appl. Genet. 2018, 131, 43–58. [Google Scholar] [CrossRef]
- Van Ooijen, J.W. JoinMap® 4, Software for the Calculation of Genetic Linkage Maps in Experimental Populations. Available online: https://www.scienceopen.com/document?vid=baa76c8c-fb55-4c13-a6ca-24c71002ab5a (accessed on 15 December 2006).
- Liu, R.H.; Meng, J.L. MapDraw:A Microsoft Excel Macro for Drawing Genetic Linkage Maps Based on Given Genetic Linkage Data. Hereditas 2003, 25, 317–321. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sources | Marker Name (SNP ID) | Primer Sequences (5’–3’) |
|---|---|---|
| 90K | BS00030364_51 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTACAGCCAAAGACCCATCTTAA |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTACAGCCAAAGACCCATCTTAG | ||
| Common reverse: GTGCGAACAGTTGACAGTGA | ||
| 90K | BS00084417_51 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTGCTGGAACTCGCCTCTTCTA |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTGCTGGAACTCGCCTCTTCTC | ||
| Common reverse: ATTTCAGCCCATCATCCTCC | ||
| 90K | RFL_Contig4423_529 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTCATACTCACAGTAACAGATTGCAT |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTCATACTCACAGTAACAGATTGCAC | ||
| Common reverse: GCTTCTGCCACAAGCTCT | ||
| 660K | AX-94995576 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTCGTGCATCTAAACCCACGTATA |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTCGTGCATCTAAACCCACGTATC | ||
| Common reverse: AGAGTCTCTATACCGTGATCCG | ||
| 660K | AX-95682571 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTATGTGTGGTTCCATTCACCG |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTATGTGTGGTTCCATTCACCA | ||
| Common reverse: CAAGTCTGAGTGGAAGAAGGAC | ||
| 660K | AX-111595417 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTTTCAGAAGAAATAGATGTATGTGCC |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTTTCAGAAGAAATAGATGTATGTGCA | ||
| Common reverse: CCATCCCATAAACCCAGGG | ||
| 660K | AX-94859941 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTCCTCAGCTTTCTTTAGGGCT |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTCCTCAGCTTTCTTTAGGGCC | ||
| Common reverse: AATTCCGGTTCACTGGTCCA | ||
| 660K | AX-111600462 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTCCAAAAATAAATTCCGCTAGTCCT |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTCCAAAAATAAATTCCGCTAGTCCC | ||
| Common reverse: CCAAAAACCCAATTGCAAGGTTAA | ||
| 660K | AX-111497410 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTGAATGCAAAGGTTGGCCCT |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTGAATGCAAAGGTTGGCCCG | ||
| Common reverse: CTCGTGCGTACCTGCTCATT | ||
| 660K | AX-109407129 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTTCTTGCCACCGTTTCAATAGAT |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTTCTTGCCACCGTTTCAATAGAC | ||
| Common reverse: CTTTTCTTGGTCTATGTAGTTGGG | ||
| 660K | AX-111128667 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTGCGGGCAAAGCTTTGGATG |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTGCGGGCAAAGCTTTGGATA | ||
| Common reverse: GAGGTAGCGACGATGTCCC | ||
| 660K | AX-111643885 | Forward A_FAM: GAAGGTGACCAAGTTCATGCTGATGCAAATCCAAACCCATTCT |
| Forward B_HEX: GAAGGTCGGAGTCAACGGATTGATGCAAATCCAAACCCATTCG | ||
| Common reverse: GCAGGAGGAGAGCTTGGGA |
| Gene | Annotation |
|---|---|
| TraesCS2B02G575700.1 | DEAD-box ATP-dependent RNA helicase 52A |
| TraesCS2B02G575800.1 | CC-NBS-LRR family disease resistance protein |
| TraesCS2B02G575900.1 | 50S ribosomal protein L14 |
| TraesCS2B02G576000.2 | OTU domain-containing protein, putative |
| TraesCS2B02G576100.2 | Calreticulin-like protein |
| TraesCS2B02G576200.1 | cysteine-rich RLK (RECEPTOR-like protein kinase) 29 |
| TraesCS2B02G576300.1 | Cysteine proteinase |
| TraesCS2B02G576400.1 | Serine/threonine-protein kinase |
| TraesCS2B02G576500.1 | Myb family transcription factor family protein |
| TraesCS2B02G576600.2 | lectin-receptor kinase |
| TraesCS2B02G576700.1 | Cytochrome P450 |
| TraesCS2B02G576800.1 | RING/U-box superfamily protein |
| TraesCS2B02G576900.1 | Chromosome segregation in meiosis protein 3 |
| TraesCS2B02G577000.1 | RING/U-box superfamily protein |
| TraesCS2B02G577100.1 | Cytochrome P450 family protein, expressed |
| TraesCS2B02G577200.1 | Cytochrome P450 family protein, expressed |
| TraesCS2B02G577300.1 | Cysteine proteinase |
| TraesCS2B02G577400.1 | Cytochrome P450 family protein, expressed |
| TraesCS2B02G577500.1 | Cysteine proteinase |
| TraesCS2B02G577600.1 | Serine/threonine-protein phosphatase 7 long form-like protein |
| TraesCS2B02G577700.1 | Phytol kinase 1 |
| TraesCS2B02G577800.1 | Exostosin family protein |
| TraesCS2B02G577900.1 | B3 domain-containing protein |
| TraesCS2B02G578000.1 | Polyadenylate-binding protein 1-B-binding protein |
| TraesCS2B02G578100.1 | ATP-dependent Clp protease proteolytic subunit 3 |
| TraesCS2B02G578200.4 | Serine/threonine-protein kinase |
| TraesCS2B02G578300.1 | Cytochrome P450, putative |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Xie, Y.; Li, Y.; Wu, S.; Li, S.; Guo, Y.; Wang, C. High-Resolution Mapping of the Novel Early Leaf Senescence Gene Els2 in Common Wheat. Plants 2020, 9, 698. https://doi.org/10.3390/plants9060698
Wang N, Xie Y, Li Y, Wu S, Li S, Guo Y, Wang C. High-Resolution Mapping of the Novel Early Leaf Senescence Gene Els2 in Common Wheat. Plants. 2020; 9(6):698. https://doi.org/10.3390/plants9060698
Chicago/Turabian StyleWang, Na, Yanzhou Xie, Yingzhuang Li, Shengnan Wu, Shuxian Li, Yu Guo, and Chengshe Wang. 2020. "High-Resolution Mapping of the Novel Early Leaf Senescence Gene Els2 in Common Wheat" Plants 9, no. 6: 698. https://doi.org/10.3390/plants9060698
APA StyleWang, N., Xie, Y., Li, Y., Wu, S., Li, S., Guo, Y., & Wang, C. (2020). High-Resolution Mapping of the Novel Early Leaf Senescence Gene Els2 in Common Wheat. Plants, 9(6), 698. https://doi.org/10.3390/plants9060698