Effects of Concentration and Temperature of Nutrient Solution on Growth and Camptothecin Accumulation of Ophiorrhiza pumila


Abstract
1. Introduction
2. Results
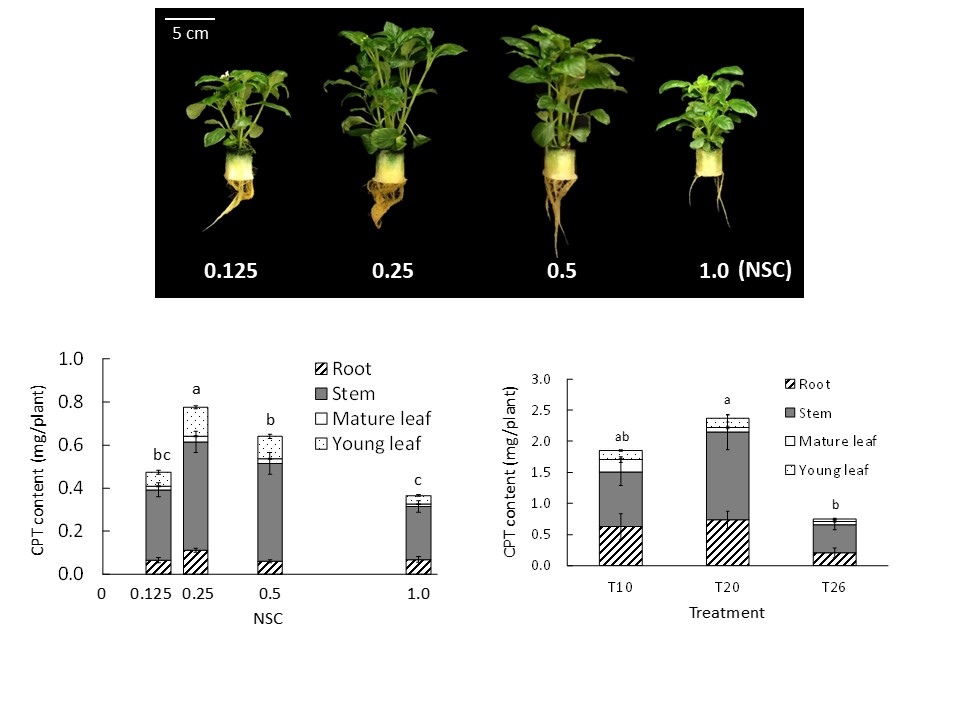
2.1. Experiment 1—Nutrient Solution Concentration (NSC)
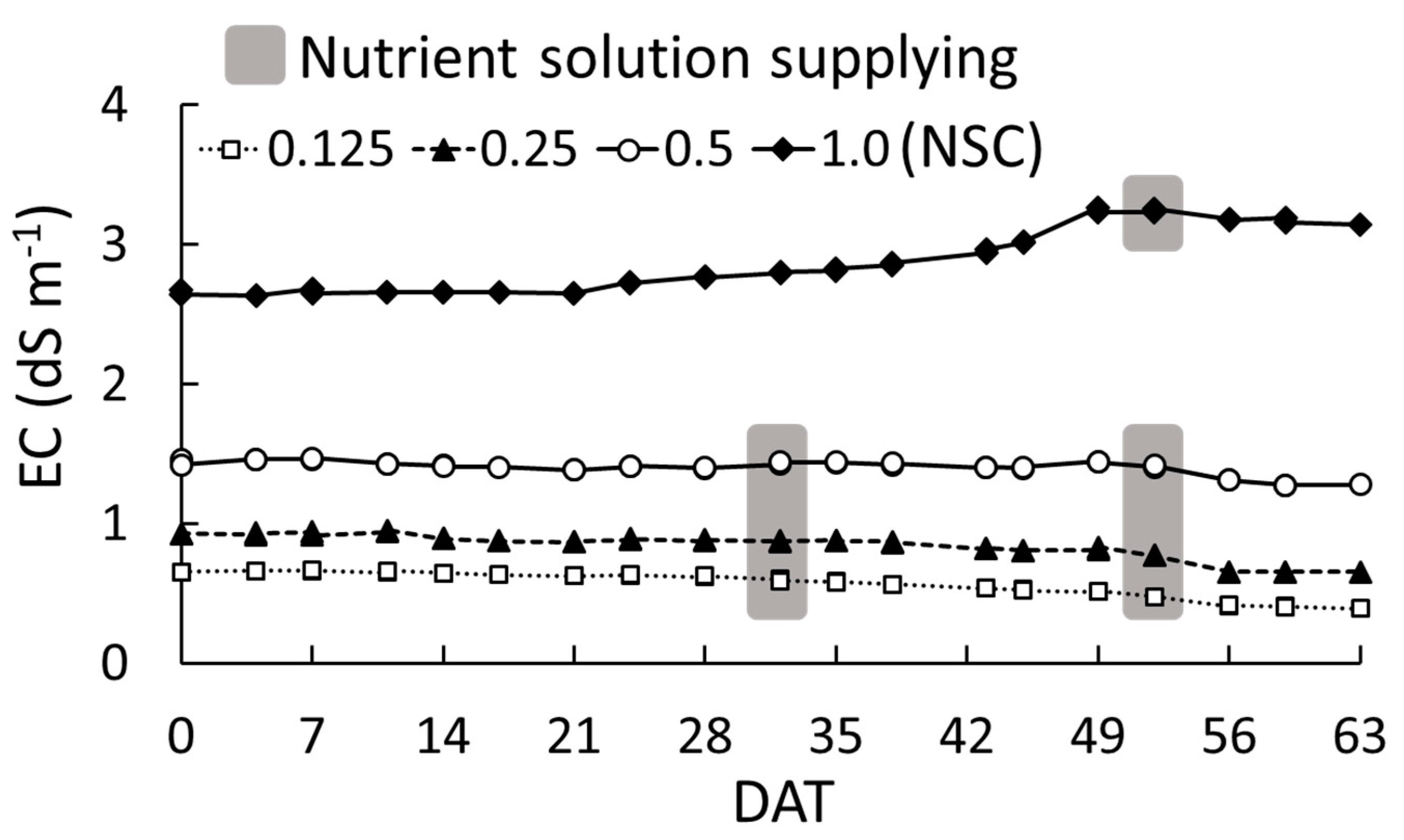
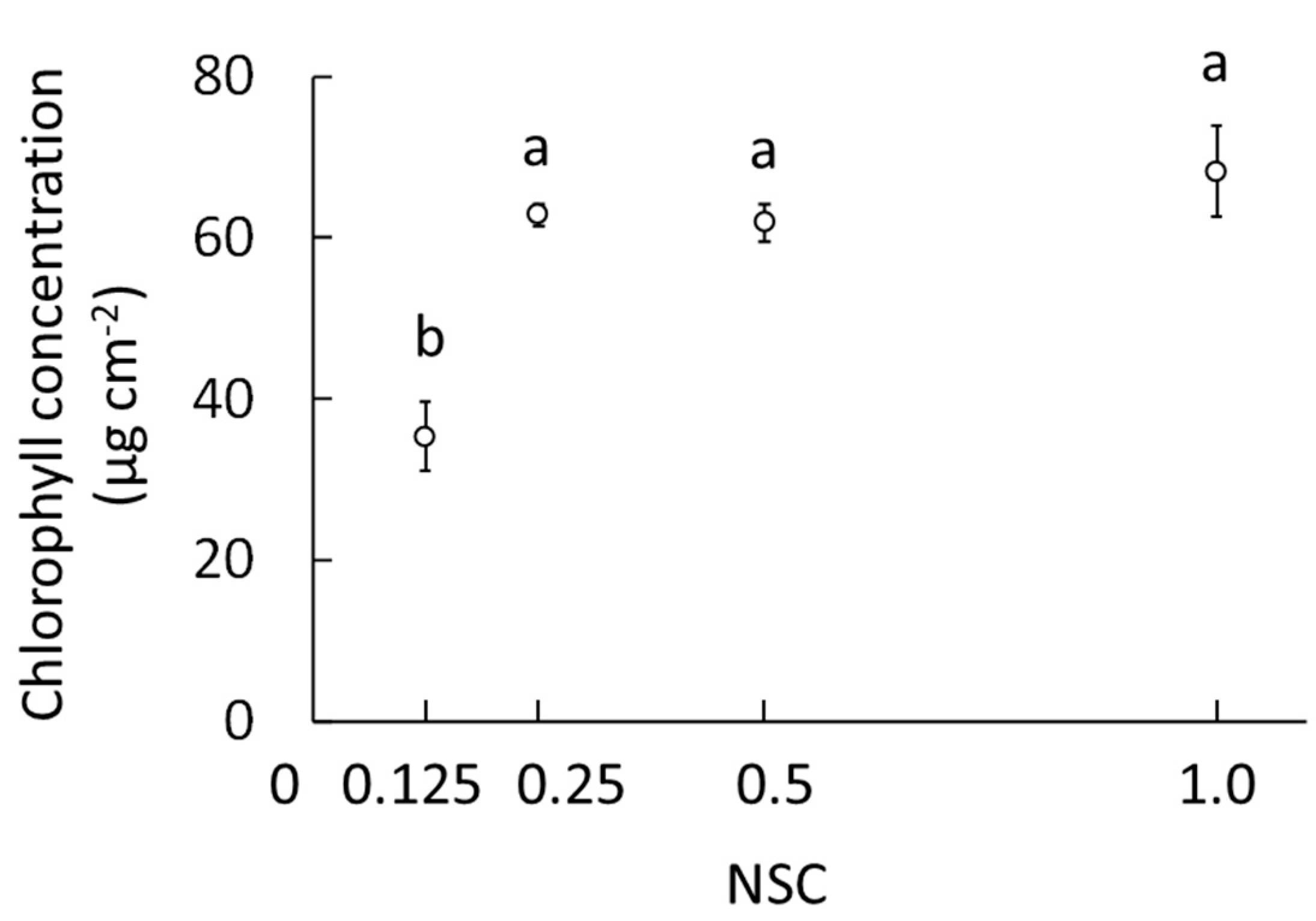
2.1.1. Electronic Conductivity, Plant Growth Characteristics, and Chlorophyll Concentration
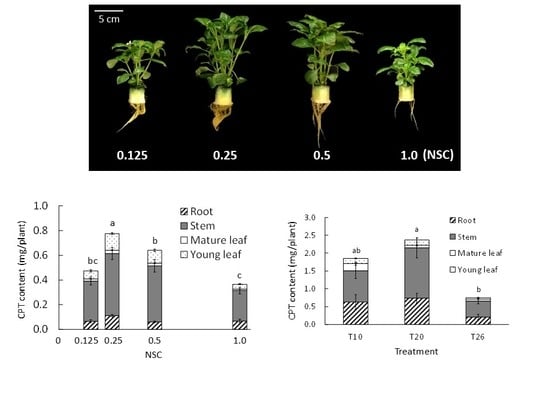
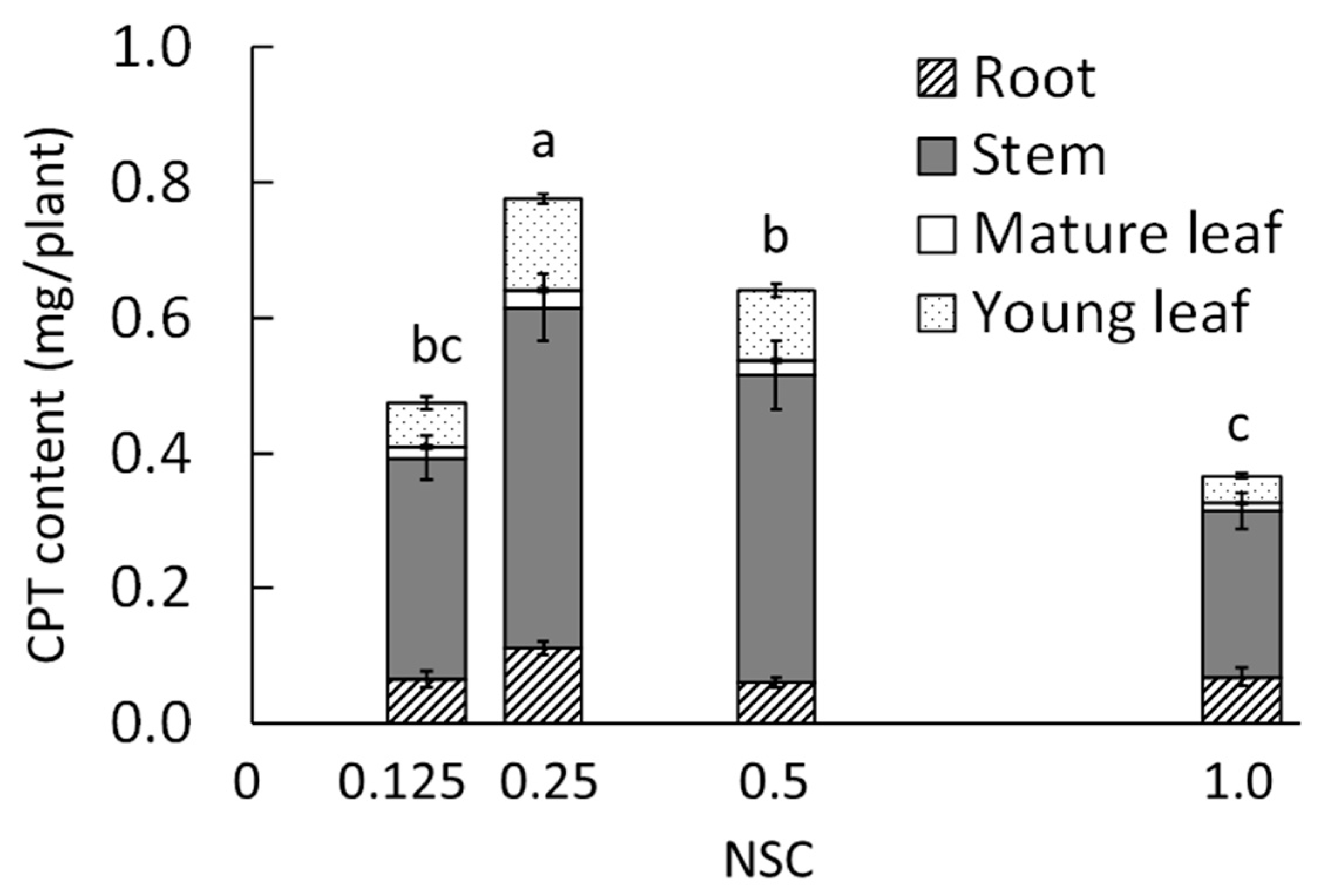
2.1.2. Camptothecin
2.2. Experiment 2—Nutrient Solution Temperature (NST)
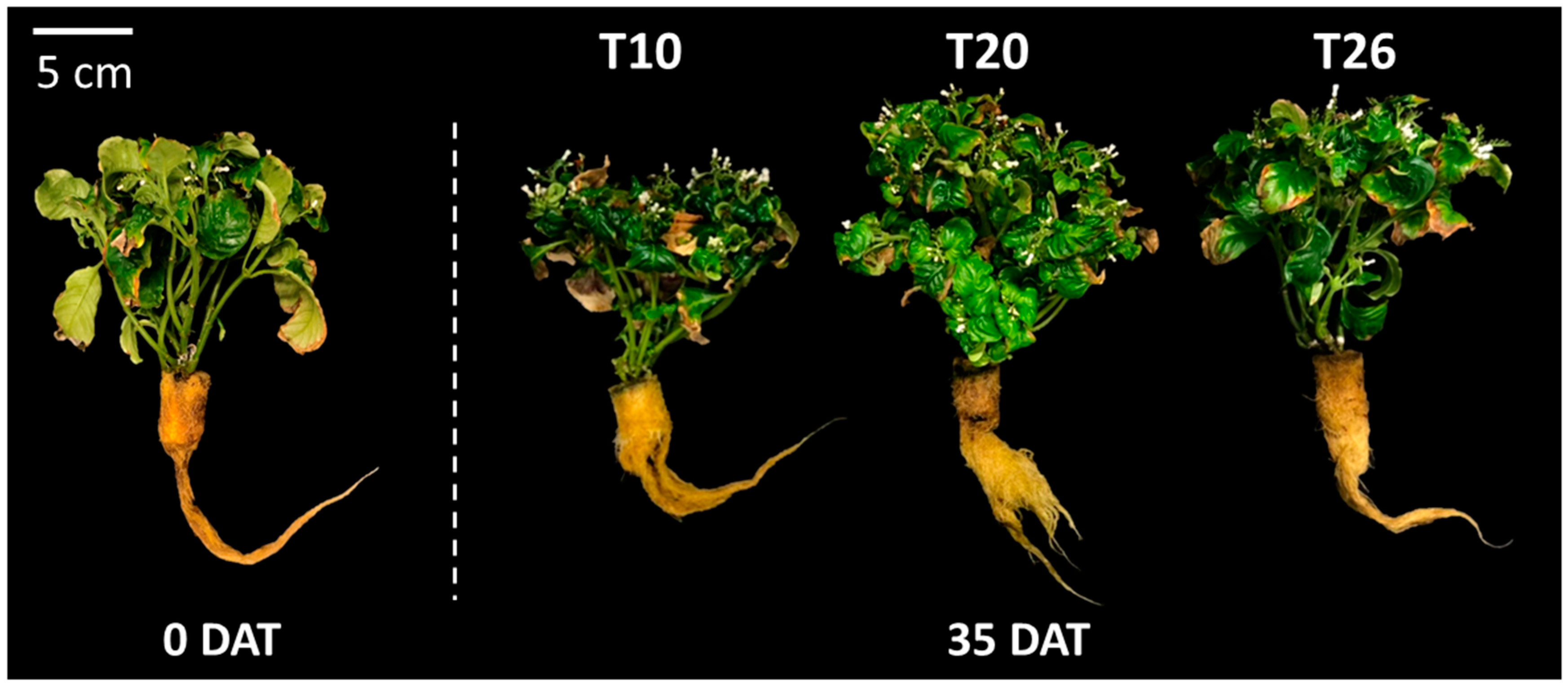
2.2.1. Plant Growth Characteristics
2.2.2. Camptothecin
3. Discussion
3.1. Experiment 1—Nutrient Solution Concentration (NSC)
3.2. Experiment 2—Nutrient Solution Temperature (NST)
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Treatment Plots
4.2.1. Experiment 1—Nutrient Solution Concentration (NSC)
4.2.2. Experiment 2—Nutrient Solution Temperature (NST)
4.3. Plant Growth Characteristics
4.4. Chlorophyll Concentration
4.5. Camptothecin Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hsiang, Y.H.; Hertzberg, R.; Hecht, S.; Liu, L.F. Camptothecin induces protein-linked DNA breaks via mammalian DNA topoisomerase I. J. Biol. Chem. 1985, 260, 14873–14878. [Google Scholar] [PubMed]
- Redinbo, M.R.; Stewart, L.; Kuhn, P.; Champoux, J.J.; Hol, W.G.J. Crystal structures of human topoisomerase I in covalent and noncovalent complexes with DNA. Science 1998, 279, 1504–1513. [Google Scholar] [CrossRef]
- Wall, M.E.; Wani, M.C.; Cook, C.E.; Palmer, K.H.; McPhail, A.T.; Sim, G.A. Plant antitumor agents. I. The isolation and structure of camptothecin, a novel alkaloidal leukemia and tumor inhibitor from Camptotheca acuminata. J. Am. Chem. Soc. 1966, 88, 3888–3890. [Google Scholar] [CrossRef]
- Houghton, P.J.; Cheshire, P.J.; Myers, L.; Stewart, C.F.; Synold, T.W.; Houghton, J.A. Evaluation of 9-dimethylaminomethyl-10-hydroxycamptothecin against xenografts derived from adult and childhood solid tumors. Cancer Chemother. Pharmacol. 1992, 31, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-Y.; Zu, Y.-G.; Shi, R.-Z.; Yao, L.-P. Review camptothecin: Current perspectives. Curr. Med. Chem. 2006, 13, 2021–2039. [Google Scholar] [CrossRef]
- Saito, K.; Sudo, H.; Yamazaki, M.; Koseki-Nakamura, M.; Kitajima, M.; Takayama, H.; Aimi, N. Feasible production of camptothecin by hairy root culture of Ophiorrhiza pumila. Plant Cell Rep. 2001, 20, 267–271. [Google Scholar] [CrossRef]
- Cui, L.; Ni, X.; Ji, Q.; Teng, X.; Yang, Y.; Wu, C.; Zekria, D.; Zhang, D.; Kai, G. Co-overexpression of geraniol-10-hydroxylase and strictosidine synthase improves anti-cancer drug camptothecin accumulation in Ophiorrhiza pumila. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef]
- Lorence, A.; Nessler, C.L. Camptothecin, over four decades of surprising findings. Phytochemistry 2004, 65, 2735–2749. [Google Scholar] [CrossRef]
- Asano, T.; Kobayashi, K.; Kashihara, E.; Sudo, H.; Sasaki, R.; Iijima, Y.; Aoki, K.; Shibata, D.; Saito, K.; Yamazaki, M. Suppression of camptothecin biosynthetic genes results in metabolic modification of secondary products in hairy roots of Ophiorrhiza pumila. Phytochemistry 2013, 91, 128–139. [Google Scholar] [CrossRef]
- Nakamura, K.; Debda, T.; Yokota, M. Homostyly and autogamy in Ophiorrhiza pumila (Rubiaceae) from the Ryukyu archipelago. J. Jpn. Bot. 2006, 81, 113–120. [Google Scholar]
- Kozai, T.; Niu, G. Role of the plant factory with artificial lighting (PFAL) in urban areas. In Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Kozai, T., Niu, G., Takagaki, M., Eds.; Academic Press: London, UK, 2015; pp. 7–33. [Google Scholar]
- Goto, E. Plant production in a closed plant factory with artificial lighting. Acta Hortic. 2012, 956, 37–49. [Google Scholar] [CrossRef]
- Abou-Hadid, A.F.; Abd-Elmoniem, E.M.; El-Shinawy, M.Z.; Abou-Elsoud, M. Electrical conductivity effect on growth and mineral composition of lettuce plants in hydroponic system. Acta Hortic. 1996, 434, 59–66. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Lucini, L.; Rea, E.; Colla, G. Nutrient solution concentration affects growth, mineral composition, phenolic acids, and flavonoids in leaves of Artichoke and Cardoon. HortScience 2012, 47, 1424–1429. [Google Scholar] [CrossRef]
- Park, S.Y.; Oh, S.B.; Kim, S.M.; Cho, Y.Y.; Oh, M.M. Evaluating the effects of a newly developed nutrient solution on growth, antioxidants, and chicoric acid contents in Crepidiastrum denticulatum. Hortic. Environ. Biotechnol. 2016, 57, 478–486. [Google Scholar] [CrossRef]
- Chadirin, Y.; Hidaka, K.; Takahashi, T.; Sago, Y.; Wajima, T.; Kitano, M. Application of temperature stress to roots of Spinach I. Effect of the low temperature stress on quality. Environ. Control Biol. 2011, 49, 133–139. [Google Scholar] [CrossRef][Green Version]
- Ogawa, E.; Hikosaka, S.; Goto, E. Effects of nutrient solution temperature on the concentration of major bioactive compounds in red perilla. J. Agric. Meteorol. 2018, 74, 71–78. [Google Scholar] [CrossRef]
- Son, K.-H.; Ide, M.; Goto, E. Growth characteristics and phytochemicals of canola (Brassica napus) grown under UV radiation and low root zone temperature in a controlled environment. Hortic. Environ. Biotechnol. 2020, 61, 267–277. [Google Scholar] [CrossRef]
- Huerta-Heredia, A.A.; Marín-López, R.; Ponce-Noyola, T.; Cerda-García-Rojas, C.M.; Trejo-Tapia, G.; Ramos-Valdivia, A.C. Oxidative stress induces alkaloid production in Uncaria tomentosa root and cell cultures in bioreactors. Eng. Life Sci. 2009, 9, 211–218. [Google Scholar] [CrossRef]
- Vera-Reyes, I.; Huerta-Heredia, A.A.; Ponce-Noyola, T.; Flores-Sanchez, I.J.; Esparza-García, F.; Cerda-García-Rojas, C.M.; Trejo-Tapia, G.; Ramos-Valdivia, A.C. Strictosidine-related enzymes involved in the alkaloid biosynthesis of Uncaria tomentosa root cultures grown under oxidative stress. Biotechnol. Prog. 2013, 29, 621–630. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Urano, A.; Sudo, H.; Kitajima, M.; Takayama, H.; Yamazaki, M.; Aimi, N.; Saito, K. Metabolite profiling of alkaloids and strictosidine synthase activity in camptothecin producing plants. Phytochemistry 2003, 62, 461–470. [Google Scholar] [CrossRef]
- Kang, J.-G.; van Iersel, M.W. Nutrient solution concentration affects shoot: Root ratio, leaf area ratio, and growth of subirrigated salvia (Salvia splendens). HortScience 2004, 39, 49–54. [Google Scholar] [CrossRef]
- Ding, X.; Jiang, Y.; Zhao, H.; Guo, D.; He, L.; Liu, F.; Zhou, Q.; Nandwani, D.; Hui, D.; Yu, J. Electrical conductivity of nutrient solution influenced photosynthesis, quality, and antioxidant enzyme activity of pakchoi (Brassica campestris L. Ssp. Chinensis) in a hydroponic system. PLoS ONE 2018, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fallovo, C.; Rouphael, Y.; Rea, E.; Battistelli, A.; Colla, G. Nutrient solution concentration and growing season affect yield and quality of Lactuca sativa L. var. acephala in floating raft culture. J. Sci. Food Agric. 2009, 89, 1682–1689. [Google Scholar] [CrossRef]
- Xu, Q.; Huang, B.; Wang, Z. Photosynthetic responses of creeping bentgrass to reduced root-zone temperatures at supraoptimal air temperature. J. Am. Soc. Hortic. Sci. 2002, 127, 754–758. [Google Scholar] [CrossRef][Green Version]
- Klock, K.A.; Taber, H.G.; Graves, W.R. Root respiration and phosphorus nutrition of tomato plants grown at a 36 °C root-zone temperature. J. Am. Soc. Hortic. Sci. 1997, 122, 175–178. [Google Scholar] [CrossRef]
- Xu, Q.; Huang, B. Effects of differential air and soil temperature on carbohydrate metabolism in creeping bentgrass. Crop Sci. 2000, 40, 1368–1374. [Google Scholar] [CrossRef]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2003, 8, 343–351. [Google Scholar] [CrossRef]
- Sakamoto, M.; Mayuka, U.; Kengo, M.; Takahiro, S. Effect of root-zone temperature on the growth and fruit quality of hydroponically grown strawberry plants. J. Agric. Sci. 2016, 8, 122–131. [Google Scholar]
- Tachibana, S. Effect of root temperature on the rate of water and nutrient absorption in cucumber cultivars and figleaf gourd. J. Jpn. Soc. Hortic. Sci. 1987, 55, 461–467. [Google Scholar] [CrossRef]
- Nagasuga, K.; Murai-Hatano, M.; Kuwagata, T. Effects of low root temperature on dry matter production and root water uptake in rice plants. Plant Prod. Sci. 2011, 14, 22–29. [Google Scholar] [CrossRef]
- Lee, J.Y.; Shimano, A.; Hikosaka, S.; Ishigami, Y.; Goto, E. Establishment of optimal light environmental conditions for camptothecin accumulation in the cultivation of Ophiorrhiza pumila based on whole-plant gas exchange rates. J. Agric. Meteorol. (under review).
- Srivastava, N.K.; Misra, A.; Srivastava, A.K.; Sharma, S. Utilization of photosynthetically fixed 14CO2 into alkaloids in relation to primary metabolites in developing leaves of Catharanthus roseus. Photosynthetica 2004, 42, 469–472. [Google Scholar] [CrossRef]
- Malik, S.; Andrade, S.A.L.; Sawaya, A.C.H.F.; Bottcher, A.; Mazzafera, P. Root-zone temperature alters alkaloid synthesis and accumulation in Catharanthus roseus and Nicotiana tabacum. Ind. Crops Prod. 2013, 49, 318–325. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NSC | Fresh Weight (g) | Total Dry Weight (mg) | ||||
|---|---|---|---|---|---|---|
| Leaf 1 | Stem | Root | Total | |||
| Young | Mature | |||||
| 0.125 | 0.9 b 2 | 1.0 | 1.7 b | 0.7 | 4.3 b | 36.6 b |
| 0.25 | 1.3 a | 1.1 | 2.9 a | 0.7 | 6.0 a | 51.3 a |
| 0.5 | 1.3 a | 1.1 | 2.9 a | 0.8 | 6.0 a | 46.3 a |
| 1.0 3 | 0.5 b | 0.8 | 1.2 b | 0.7 | 3.2 b | 31.5 b |
| NSC | Camptothecin Concentration (mg g−1 DW) | |||
|---|---|---|---|---|
| Leaf 1 | Stem | Root | ||
| Young | Mature | |||
| 0.125 | 0.8 b 2 | 0.1 | 2.2 | 1.0 b |
| 0.25 | 1.2 a | 0.2 | 2.0 | 1.7 a |
| 0.5 | 1.0 a | 0.2 | 2.0 | 0.9 b |
| 1.0 3 | 0.7 b | 0.1 | 1.9 | 1.0 b |
| DAT | NST (°C) | Fresh Weight (g) | Total Dry Weight (g) | Dry Matter Ratio (%) | Leaf Area (cm2) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf 1 | Stem | Others 3 | Root | Total | Shoot | Root | |||||
| Young | Mature | ||||||||||
| 0 | 0.5 | 2.6 | 3.8 | 2.3 | 2.5 | 14.2 | 1.4 | 10.8 | 4.3 | 35.9 | |
| 35 | T10 | 1.3 | 3.0 | 4.5 | 4.5 b 2 | 4.7 | 18.0 b | 2.9 b | 19.4 | 7.2 a | 151.7 |
| T20 | 1.4 | 5.3 | 7.4 | 9.7 a | 6.2 | 30.1 a | 4.6 a | 17.7 | 6.0 ab | 254.2 | |
| T26 4 | 1.1 | 4.1 | 6.3 | 3.3 b | 4.2 | 18.9 b | 2.6 ab | 16.3 | 5.2 b | 202.0 | |
| NST (°C) | Camptothecin Concentration (mg g−1 DW) | |||
|---|---|---|---|---|
| Leaf 1 | Stem | Root | ||
| Young | Mature | |||
| T10 | 0.5 | 0.3 a 2 | 1.3 a | 1.8 a |
| T20 | 0.5 | 0.1 b | 1.4 a | 2.0 a |
| T26 3 | 0.4 | 0.1 b | 0.8 b | 1.0 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-Y.; Hiyama, M.; Hikosaka, S.; Goto, E. Effects of Concentration and Temperature of Nutrient Solution on Growth and Camptothecin Accumulation of Ophiorrhiza pumila. Plants 2020, 9, 793. https://doi.org/10.3390/plants9060793
Lee J-Y, Hiyama M, Hikosaka S, Goto E. Effects of Concentration and Temperature of Nutrient Solution on Growth and Camptothecin Accumulation of Ophiorrhiza pumila. Plants. 2020; 9(6):793. https://doi.org/10.3390/plants9060793
Chicago/Turabian StyleLee, Ji-Yoon, Miki Hiyama, Shoko Hikosaka, and Eiji Goto. 2020. "Effects of Concentration and Temperature of Nutrient Solution on Growth and Camptothecin Accumulation of Ophiorrhiza pumila" Plants 9, no. 6: 793. https://doi.org/10.3390/plants9060793
APA StyleLee, J.-Y., Hiyama, M., Hikosaka, S., & Goto, E. (2020). Effects of Concentration and Temperature of Nutrient Solution on Growth and Camptothecin Accumulation of Ophiorrhiza pumila. Plants, 9(6), 793. https://doi.org/10.3390/plants9060793