Indications for a Central Role of Hexokinase Activity in Natural Variation of Heat Acclimation in Arabidopsis thaliana



{kind=link}
{kind=link}
{kind=link}
{kind=link}
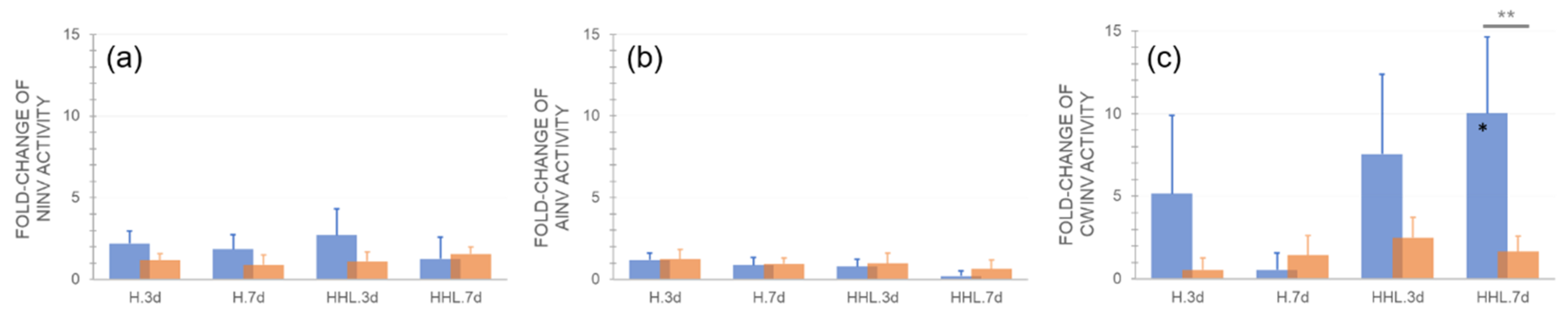{kind=link}
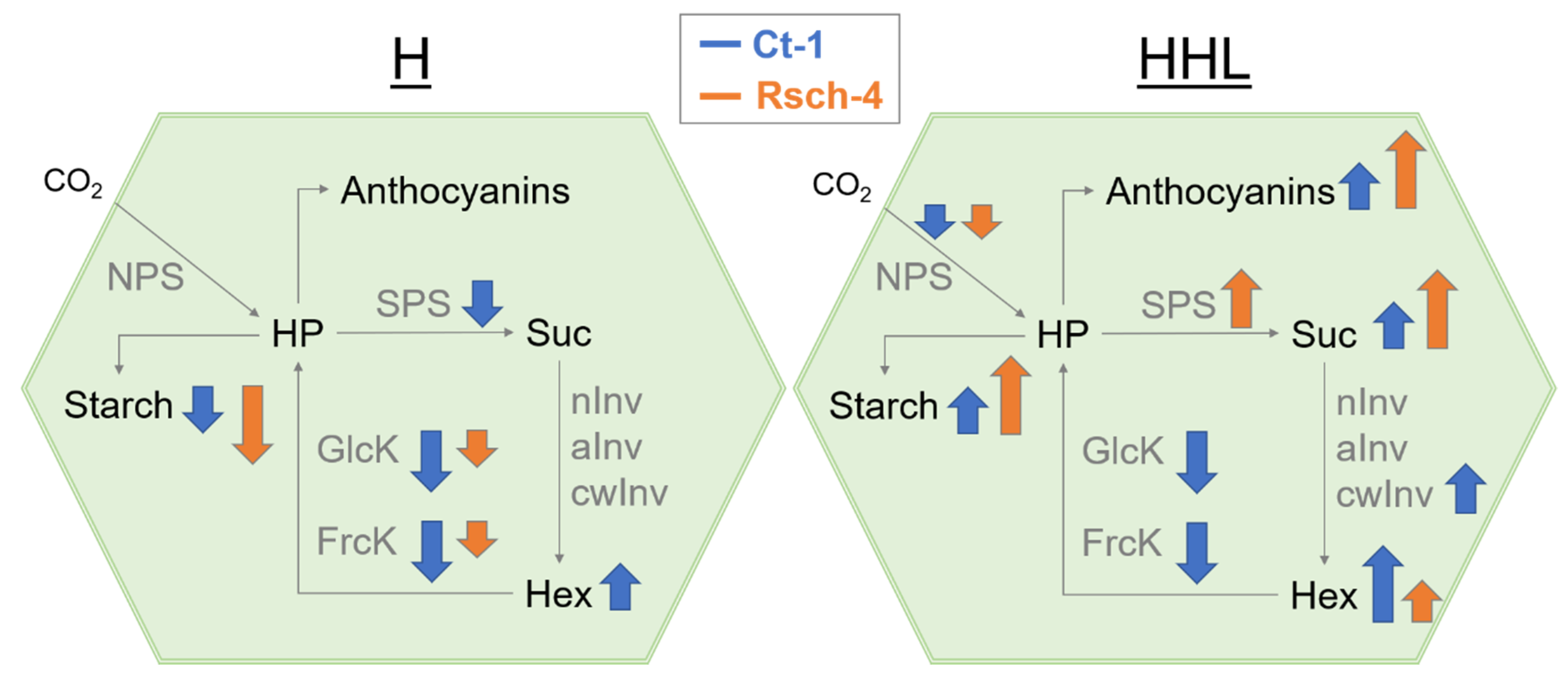{kind=link}
Abstract
Share and Cite
Atanasov, V.; Fürtauer, L.; Nägele, T. Indications for a Central Role of Hexokinase Activity in Natural Variation of Heat Acclimation in Arabidopsis thaliana. Plants 2020, 9, 819. https://doi.org/10.3390/plants9070819
Atanasov V, Fürtauer L, Nägele T. Indications for a Central Role of Hexokinase Activity in Natural Variation of Heat Acclimation in Arabidopsis thaliana. Plants. 2020; 9(7):819. https://doi.org/10.3390/plants9070819
Chicago/Turabian StyleAtanasov, Vasil, Lisa Fürtauer, and Thomas Nägele. 2020. "Indications for a Central Role of Hexokinase Activity in Natural Variation of Heat Acclimation in Arabidopsis thaliana" Plants 9, no. 7: 819. https://doi.org/10.3390/plants9070819
APA StyleAtanasov, V., Fürtauer, L., & Nägele, T. (2020). Indications for a Central Role of Hexokinase Activity in Natural Variation of Heat Acclimation in Arabidopsis thaliana. Plants, 9(7), 819. https://doi.org/10.3390/plants9070819