Distinct Responses to Light in Plants


BioISI—Biosystems & Integrative Sciences Institute, Faculty of Sciences, University of Lisbon, 1749-016 Lisbon, Portugal
Plants 2020, 9(7), 894; https://doi.org/10.3390/plants9070894
Submission received: 22 June 2020
/
Revised: 13 July 2020
/
Accepted: 13 July 2020
/
Published: 15 July 2020
(This article belongs to the Special Issue Light Signaling in Plants)
{kind=link}
{kind=link}
Abstract
:The development of almost every living organism is, to some extent, regulated by light. When discussing light regulation on biological systems, one is referring to the sun that has long been positioned in the center of the solar system. Through light regulation, all life forms have evolved around the presence of the sun. As soon our planet started to develop an atmospheric shield against most of the detrimental solar UV rays, life invaded land, and in the presence of water, it thrived. Especially for plants, light (solar radiation) is the source of energy that controls a high number of developmental aspects of growth, a process called photomorphogenesis. Once hypocotyls reach soil′s surface, its elongation deaccelerates, and the photosynthetic apparatus is established for an autotrophic growth due to the presence of light. Plants can sense light intensities, light quality, light direction, and light duration through photoreceptors that accurately detect alterations in the spectral composition (UV-B to far-red) and are located throughout the plant. The most well-known mechanism promoted by light occurring on plants is photosynthesis, which converts light energy into carbohydrates. Plants also use light to signal the beginning/end of key developmental processes such as the transition to flowering and dormancy. These two processes are particularly important for plant´s yield, since transition to flowering reduces the duration of the vegetative stage, and for plants growing under temperate or boreal climates, dormancy leads to a complete growth arrest. Understanding how light affects these processes enables plant breeders to produce crops which are able to retard the transition to flowering and avoid dormancy, increasing the yield of the plant.
1. Overview of Light Transduction Scheme
All life is shaped by the Earth’s rotation and orbit around its own axis and the sun, providing a diurnal rhythm of day and night, and depending on the latitude, an annual cycle of day length which reaches its maximum and minimum on the solstices of summer and winter, respectively. Plants use dawn and dusk as signals to organize all aspects of their growth, such as photosynthesis. Light is an environmental factor permanently present, and yet displays a dynamic control on components of plant’s functionality such as germination, phototropism, and reproduction. The decreasing number of hours of light, which starts right after the beginning of autumn, also works as an indicator to enter dormancy, preventing the detrimental effects that winter conditions might have on plant cells. Plants also react to variations of low and high light intensities (e.g., acclimation) [1], which involves a set of environmental adjustments such as leaf morphology variations, chloroplast structure, and/or modification of the composition of the photosynthetic electron transport chain, consequently affecting photosynthesis [2,3]. Plants growing under limiting light conditions show leaf and chloroplast movements toward the source of light, as well as general bending to maximize light capture, increasing photosynthetic efficiency. On the other hand, when plants are submitted to environmental conditions of high light intensity and UV-B, the chloroplasts and nucleus move away from the light, and protective photoreceptors pigments are synthesized to prevent photodamage [4,5]. These two types of acclimation responses can be reversible (dynamic acclimation) or can become permanent, resulting in irreversible or developmental acclimation mechanisms adopted by the plant [6,7].
The primary determinant of crop yield is the cumulative rate of photosynthesis over the growing season, which is regulated by the amount of light captured and its ability to efficiently use energy to convert CO2 into biomass and harvestable yield stored as carbohydrates [8]. The use of light during photosynthesis is achieved through two photosystems present in the chloroplast’s photosynthetic pigments, which show a narrow pick absorption range of the solar spectrum despite pigments absorbed throughout the photosynthetically active radiation (PAR) spectrum. Unlike the light regulation mediated by photoreceptor genes present in the nucleus and responsible for physiological responses such as germination, flowering, circadian clock input, and dormancy, photosynthesis requires a coordinated regulation of the nuclear and plastidic genomes [9]. In the chloroplast, photons of light are converted into glucose molecules in a two-step reaction pathway. First, energy is stored in the bonds of adenosine triphosphate (ATP) and nicotinamide adenine dinucleotide phosphate (NADPH), which are then used in the Calvin cycle (second step) to produce organic molecules after combining carbon atoms from carbon dioxide (CO2) with glucose as the end result. This six-carbon molecule is then utilized by the mitochondria to produce ATP. Diurnal cycle induces two distinct photosynthetic reactions: The light reaction, where water molecules are split into oxygen, hydrogen, protons, and electrons; and the dark reaction, where protons and electrons are taken to reduce CO2 to carbohydrates [10]. This way, photosynthesis is the primary source of the food chain because it promotes the conversion of solar energy into chemical energy.
Plants are able to utilize their internal circadian clock to synchronize physiology and development according to daily and yearly environmental changes and season cycles [11]. The circadian clock comprises a substantial number of gene activations and inhibitions in multiple feedback regulations, which are focused on two transcription factors containing DNA-binding motifs MYB: CIRCADIAN CLOCK-ASSOCIATED 1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY) [12]. These proteins activate the transcription of PSEUDO-RESPONSE REGULATOR 9 (PRR9) and PRR7 in the morning [13]. The diurnal cascade of gene activation in Arabidopsis initiates at dawn with a pick of CCA1 transcripts. Throughout the day, PRR9, PRR7, and PRR5 spike up early in the morning, mid-day, and afternoon, respectively. The feedback activation of these genes starts with the binding of CCA1 to the PRR9 and PRR7 promoter region, with further activation of these genes. In turn, PRR9 and PRR7 proteins bind to the CCA1 promoter, inhibiting its transcription during the day [14]. Alternatively, CCA1 and LHY inhibit the expression of other clock genes that accumulate in the afternoon and evening, such as PRR5, TIMING OF CAB EXPRESSION 1 (TOC1), CCA1 HIKING EXPEDITION (CHE), GIGANTEA (GI), LUX ARRHYTHMO (LUX), and EARLY FLOWERING 4 (ELF4) [15,16]. As night approaches, CCA1 and LHY protein levels decrease, and its repressing action no longer takes place on its target genes, leading to the accumulation of the evening clock gene transcripts [17]. Besides the regulation of PRR7 and PRR5, still during the day, CCA1 and LHY also regulate the expression timing of GI and FLAVIN-BINDING KELCH REPEAT F-Box 1 (FKF1), all of which regulate CONSTANT (CO) expression [18,19]. CO is an important integration protein in the control of flowering in the correct season [20]. The PRRs also regulate the expression of the CO repressor, CYCLING DOF FACTOR 1 (CDF1) [19]. In turn, its degradation is mediated by the GI-CDF complex with the subsequent activation of CO [21]. Another circadian clock feedback activation mechanism takes place during the morning, when CDF1 with other CDFs and FLOWERING BHLHs (FBHs) repress CO expression. The FKF1-GI complex degrades CDFs, facilitating the expression of CO [22]. The GI and ZEITLUPE (ZTL) complex are also responsible for TOC1 degradation by the 26S proteasome upon blue light and temperature sensing [23]. All these complex regulatory mechanisms permit the precise expression of the florigen FLOWERING LOCUS T (FT), which is responsible for flowering. For example, in Arabidopsis, a long-day (LD) responsive plant, flowering is promoted by CO, which, in turn, activates FT under LD [24].
Fully expanded leaves, which play an important role in capturing light, are also important for flowering, as FT is expressed in specific phloem companion cells in this organ [25]. The florigen FT is mobile, travelling from leaves to the SAM, where it promotes flowering in Arabidopsis [26] through the initiation of the expression of floral identity genes by binding to the bZIP transcription factor FD [27]. In the tree Populus, however, it is possible to identify two FT homologues acting in different pathways: FT1 is involved in the reproductive onset with its transcript picking under short days (winter time), whereas FT2 is involved in vegetative growth and cessation or bud set (dormancy) in response to seasonal day length changes [28]. Trees growing under temperate and boreal climates must adapt their growth in order to avoid the detrimental effects that winter cold might inflict on cells. This way, as day lengths become shorter, FT2 expression is reduced, and both apical and radial growth are arrested [29]. FT2 protein is an important integrator of light regulation which controls vegetative growth under LD. The transcription of FT2 takes place upon CO accumulation, which is mediated by the activation of phytochromes [30]. Notably, the required FT2 transcription threshold, which is achieved in long days, coincides with the higher photosynthetic rate, resulting in the necessary carbon supply for active growth.
Plants make use of a vast number of types of photoreceptors so they can accurately detect the changes of the spectral composition (UV-B to far-red) and light direction and duration (photoperiod). Regardless of the fact photoreceptors can be found throughout the plant, in most cases, the site of light perception is the same as that where responses to the light stimulus take place. Nevertheless, there are examples where the site of light response is distant from the location of light perception, such as those with FTs responsible for floral transition and LD responses [31]. Photoreceptors are divided according to its wavelength sensing. They are named phytochromes in response to red/far-red (R/FR) light; cryptochromes, phototropins, and zeitlupes when sensing blue/UV-A light; and UV-B photoreceptors (UVR8) when responding to UV-B (Figure 1) [32]. Once photoactivated, photoreceptors interact with other players involved in light responses, such as ELONGATED HYPOCOTYL 5 (HY5), a photomorphogenesis promoting transcription factor, and the CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) - SUPPRESSOR OF PHYA-105 (SPA) complex, among others. COP1, a RING E3 ubiquitin ligase, forms a complex with SPA protein family (SPA1-SPA4), targeting key regulators for degradation [33]. Especially for SPA2, its action is rapidly repressed due to its degradation promoted by light-activated phytochrome photoreceptors but not by cryptochromes. The degradation of SPA2 is dependent on COP1′s ubiquitin role, meaning that the light regulation of COP1-SPA complex is achieved through the degradation of SPA2 protein but not of COP1 [34]. During the night, the COP1-SPA complex is no longer degraded by light-activated photoreceptors, enabling the target of HY5 and CO for ubiquitination and degradation with consequent suppression of photomorphogenesis and flowering, respectively [35]. DE-ETIOLATED 1 (DET1) is a protein that interacts with COP10 and DAMAGED DNA BINDING PROTEIN 1 (DDB1), establishing a complex that assists COP1 mediating the degradation of transcription factors responsible for light responses such as HY5, HY5 HOMOLOGUE (HYH), LONG AFTER FAR-RED LIGHT1 (LAF1), and LONG HYPOCOTYL IN FAR-RED1 (HFR1) [36]. In sum, COP1 and DET1 belong to the regulatory ubiquitin-mediated proteolic degradation machinery responsible for the inhibition of photomorphogenesis during the dark period.
The diverse photomorphogenic responses of plants, such as seed germination, de-etiolation, and circadian clock regulation, are, to a great extent, associated with light perception by photoreceptor system [37]. Photoreceptors act through the presence of specific photoreceptive chromophores [38,39], and are the gatekeepers for different types of light sensing which are responsible for regulating a high number of physiologically and developmental downstream signaling pathways [40].
2. Introduction to Photoreceptors
2.1. Phytochromes
It is possible to identify five photochromes in Arabidopsis: phyA, phyB, phyC, phyD, and phyE [41,42]. They are involved in several life cycle functions, including germination, de-etiolation, stomata development, flowering transition, senescence, and shade avoidance [43,44]. Phys are synthesized as a biologically inactive (Pr) form that is converted to an active (Pfr) form after red light absorption. This latter form is rapidly converted back to the inactive Pr state upon far-red light irradiation or slowly by dark reversion [45]. Pfr leads to the inhibition of several PHYTOCHROME INTERACTING FACTORs (PIFs) either by triggering their proteasome-mediated degradation or through their phosphorylation [44] (Figure 1a). The ubiquitin E3 ligase complex (COP1/SPA/FUS) is also inhibited by Pfr, enabling the stabilization of the transcription factors required for light-growth development such as HY5 [46]. In the dark, upon Pfr form spontaneously change back to the Pr form after phosphorylation, it acquires the capacity to enter the nucleus and establish subnuclear foci called photobodies [47]. These are heterogeneous, meaning that they can change composition throughout the day [48,49]. Photobodies act upstream of PIFs by promoting their phosphorylation and targeting them for degradation via ubiquitination [50,51].
2.2. Cryptochromes
Arabidopsis has two cryptochromes, Cry1 and Cry2 [52], both present in the nucleus. When activated by blue light, Cry1 migrates to cytosol, while Cry2 remains in the nucleus [53]. Cry1 plays a predominant function during de-etiolation, and Cry2 displays its main role in the photoperiodic control of flowering [54]. The light-activated Cry1 binds to SPA1, leading to the inhibition of the COP1-SPA1 E3 ubiquitin ligase, which, consequently, gives rise to an accumulation of HY5 and CO transcription factors, promoting de-etiolation and flowering, respectively [55,56] (Figure 1b). Regarding Cry2, once activated by blue light, it interacts with CRY-INTERACTING bHLH (CIB1, 2, 4, 5), directly activating the transcription factor FLOWERING LOCUS T (FT) in order to promote flowering [57]. The other CRY-interacting protein is the SUPRESSOR OF PHYTOCHROME A-105 1 (SPA1), which is responsible for positive regulation of the E3 ubiquitin ligase COP1 [55,58], leading to an inhibition of COP1 dependent degradation of transcriptional regulators such as bZIP transcription factor HYH and bHLH transcription factor HFR1 [56].
2.3. Phototropins
The photoreceptors that perceived UV-A and blue light are called phototropins (phots). Arabidopsis genome holds two: PHOT1 and PHOT2 [59]. The blue light is sensed by two flavin monoclueotide (FMN) chromophore-binding light oxygen voltage (LOV1 and LOV2) domains [60]. The conformational change promoted by light leads to autophosphorylation of the residues located both in the sensory and kinase domains [61,62]. Blue light triggers both phots but in different manners: It stimulates PHOT2 and downregulates PHOT1 [63]. These two genes are responsible for distinct mechanisms, such as phototropism and leaf positioning [64], the opening of stomas, and the accumulation of chloroplasts [65]. The activated phototropins bind to NONPHOTOTROPIC HYPOCOTYL 3 (NPH3) and ROOT PHOTOTROPISM2 (RPT2), initiating the phot signaling [66].
2.4. Zeitlupe Family
The zeitlupe family of LOV UV-A/blue light photoreceptors comprise ZEITLUPE (ZTL), FLAVIN-BINDING, KELCH REPEAT, F-BOX (FKF1), and LOV KELCH PROTEIN2 (LKP2) proteins [60]. These proteins contain only one LOV domain, followed by an F-box and six KELCH repeats [67]. Zeitlupes form SCF E3 ubiquitin ligase complexes, which directly control light-mediated protein degradation [21]. This activity has an important role in the photoperiodic control of floral transition [65], circadian oscillator regulation, and hypocotyl elongation [68].
The mechanism of action of ZTL, LKP2, and FKF1 is through the ubiquitin-mediated degradation of clock elements such as TOC1 and PRR5 [67,69]. The interaction of FKF1, ZTL, and LKP2 with GI is induced by blue light (Figure 1c). The FKF1-GI complex interacts with CDF1 (a repressor of CO and FT expression), leading to its degradation by the ubiquitin-proteasome system with subsequent expression of CO [22] and FT. The KELCH repeats are the regions responsible for substrate proteins recognition for ubiquitination [70], and both FKF1 and LKP2 are able to interact with CDF1 [71]. In sum, the light activation of the LOV domain in the zeitlupe family members promotes an activation with GI and modification of its ubiquitin E3 ligase activity. This way, the GI-FKF1 complex actively triggers the CO repressor for degradation. With this repression lifted, CO is expressed under light, modulating flowering. The ZTL-GI complex, on the other hand, when regulated by light, limits the ZTL degradation capacity of this target proteins, controlling the circadian clock response [59].
2.5. UVR8
The UV RESISTANT LOCUS 8 (UVR8) photoreceptor is on the front line of UV-B stress-induced responses and triggers a wide range of changes in gene expression, leading to morphological adaptions and the production of flavonols that act as UV-B protective shields [72]. It also mediates phototropic bending, stomatal movement, and circadian clock cascade gene activation [73,74]. The inactive form of UVR8 is a homodimer but is converted to an active monomer after absorbing UV-B [75]. In its active monomeric form, UVR8 interacts with E3 ubiquitin ligase COP1 in the nucleus [76], leading to the expression and stabilization of HY5 and HY5 HOMOLOGUE (HYH), which, in turn, bind to the promotor of several UV-B responsive genes [77] (Figure 1d), such as those involved in the flavonoid synthesis, and chalcone synthase [78,79], which is important in abiotic and biotic stress responses. When plants are subjected to low UV-light intensities for short periods of time, their actions beneficially enhance their resistance to pathogens and herbivores [80] but increase UV-light exposure, stimulates the generation of free radicals detrimental for DNA, proteins, lipids, chloroplasts, and photosynthetic pigments, hence impairing yield [81].
3. Plant Development under Light
3.1. Mechanisms Regulated by the Action of Light
Plants can sense light signals through photoreceptors, which are capable of transmitting those signals into a light-signaling response cascade [82]. The bHLH transcription factors PHYTOCHROME-INTERACTING FACTORs (PIFs), acting downstream of phytochromes, show a repressive action on photomorphogenesis during darkness or, in other words, promote skotomorphogenesis growth in the absence of light [44]. The PIFs family comprises PIF1/PIF3-LIKE5 (PIL5), PIF3, PIF4, PIF5/PIL6, and PIF6/PIL2 [42]. Under light conditions, active phytochromes interact with PIFs, resulting in phosphorylation and further PIFs degradation by proteasome, resulting in the photomorphogenesis responses. Recent findings on the regulation of PIF3, responsible for hypocotyl elongation, cotyledon expansion, and chloroplast development, have shown that new factors are involved in the degradation of PIF3 by the SKP1-CUL1-F-box protein complex (SCF), EIN3-BINDING F BOX PROTEINs (EBFs) 1 and 2, E3 ubiquitin LIGHT-RESPONSE BRIC-A-BRACK/TRAMTRACK/BROAD (LRB), and Photoregulatory Protein Kinases (PPKs) [83,84,85]. Regardless of PIF3’s ability to bind EBFs under light or dark conditions, the recruitment of PIF3-EBFs to the SCFEBF1/2 only occurs under light and after phosphorylation of PIF3 [84]. LRB, on the other hand, only targets phyB and PIF3 under high-light environments without affecting the stability of phyB [83]. PIF3 and phyB also interact with PPKs in a light-dependent fashion, which is required for PIF3′s phosphorylation [85]. With the migration of phyB to the nucleus upon activation by light, it binds to PIF3, followed by the activation of PPK with consequent phosphorylation of PIF3. The phosphorylated PIF3 form is recognized by the LRB E3 ligases, resulting in the degradation by ubiquitination of both PIF3 and phyB [83,85].
Other PIFs are also involved in the control of germination. Specifically, PIF1 represses germination and de-etiolation [86]. This negative regulation is achieved through the action of the KELCH F-BOX protein COLD TEMPERATURE GERMINATION (CTG10), which recognizes, binds, and destabilizes PIF1, hence stimulating the completion of germination and seedling de-etiolation. This activation also shows a feedback loop where PIF1 downregulates the transcription of CTG10 [87]. Light quality also affects seed germination through the regulation of gibberelins (GA) and abscisic acid (ABA) levels. Whereas GA induces germination, ABA inhibits it. Valstij et al. [88] showed that under far-red conditions (usually shade environments), the gene MOTHER-OF-FT-AND-TFL1 (MFT) represses germination by regulating both ABA and GA signaling pathways. In the dark, PIF1 promotes the expression of ABA-INSENSITIVE 5 (ABI5), DELLA-encoding GA-INSENSITIVE (GAI), and REPRESSOR-OF-GA1 (RGA) [89]. The control of ABA and GA accumulation by PIF1 is also exerted by promoting the activation of SOMNUS (SOM), which is responsible for the increasing of ABA and decreasing of GA levels [90].
3.2. Flowering Control by CO/FT Regulation
Flowering is a fundamental phase of plant’s development and is of great importance when dealing with species that are the sources of seeds which are essential for feeding purposes. Flowering regulation depends on internal and external signals such as temperature and daylength, which are converted into the regulation of two major floral integrator genes: FT and CO [91]. FT is upregulated by the protein CO in the photoperiodic pathway, which is initiated by the photoreceptors CRY1, CRY2, and FKF1 [27].
Day length must reach a threshold in order to promote the stabilization of the CO protein. Only under LD, CO is not degraded, picking its accumulation during the night [92,93]. CO, alone or combined with other factors, activates the transcription of FT gene (Figure 2). The flowering time is dependent on the amount of FT protein present in the apical meristems, which was previously transported from leaves, where the FT gene is regulated by a high number of factors. CO presents a tight transcriptional and post-transcriptional regulation, which is important to measure the day length and calculate when to promote flowering. The circadian clock FKF1, GI, and CDF are major factors in the regulation of CO [71]. In the case of CDF1 protein, it directly binds to the CO promoter and represses its expression, which results in a reduced level of CO transcripts in the morning [94] (Figure 2a). Conversely, CDF1 is positively regulated by CCA1 and LFY at dawn and negatively regulated by PRR9, PRR7, and PRR5 in the afternoon [95].
Under LD days, the FKF1-GI complex degrades CDF proteins, preventing its repression on the CO promoter [96]. As days shorten, FKF1 and GI transcription levels no longer pick at the same time. Hence, the FKF1-GI complex cannot be established, and CDF proteins are not degraded, remaining present on the CO promoter. In this way, CO transcription is prevented, which, in turn, does not activate FT expression during the day under SD [93]. Other genes are involved in the regulation of the CO-FT pathway. ZTL and LKP2 interact with FKF1 and GI to help in the destabilization of the CDF2 protein [22]. The bHLH (basic helix-loop-helix) transcription factors FLOWERING BHLH1 (FBH1), FBH2, FBH3, and FBH4 promote the activation of CO once the CDF repressor is removed from the CO promoter region [97]. Albeit CO transcripts accumulate at night both under LD and SD, their effects on the FT gene only take place at dusk under LD. CO proteins are degraded by COP1-SPA1 complex during the night (Figure 2a) [98], while during the first hours of the day, CO degradation is carried out by the HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE1 (HOS1), a RING finger containing E3 ubiquitin ligase [99]. PHYB also mediates CO protein degradation in the morning, contrarily to PHYA, which stabilizes CO later in the day [100]. CRY1 and CRY2 are another group of photoreceptors involved in CO protein regulation [55,101] with CRY2-SPA1 upon light activation, promoting the interaction between CRY2 and COP1 and inhibiting the capacity that COP1 has to degrade CO (Figure 2a) [55].
The transcriptional activation/inhibition of FT is the final approach for flowering regulation initiated by light sensing. The heterodimer comprising FLC and SHORT VEGETATIVE PHASE (SVP) directly suppress FT and SOC1 expression [102]. FT transcription is also inhibited by the FLC-FLM (FLOWERWING LOCUS M)-MAFs (MADS AFFECTING FLOWERING) complex [103] and by the Polycomb group complex (PRC 1-like complex) composed by the EMBRYONIC C FLOWER 1 (EMF1), LHP1 and histone E3 lysine-4 [92]. FT is negatively regulated by three more transcription factors: SCHLAFMÜTZE (SMZ), TEMPRANILLO1 (TEM1), and TEM2 (Figure 2b) [104,105]. As FT activators, one finds CIBs and CO proteins. CIBs interact directly with FT promoter, activating its expression, but the CIB1 function is restricted from late afternoon to early night and forms a complex with CRY2 after photoexcitement by blue light (Figure 2b). In the absence of blue light, CIB1 is targeted for degradation [106], while its stabilization is mediated by ZTL and LKP2 under blue light with further FT expression [101].
4. Conclusions
The influence of light on plant development, curiously, starts with the lack of light during seed germination until soil surface is reached. At this time, plant development suffers a transition from the heterotrophic growth, where maternally deposited energy reserves are used to fuel an autotrophic photosynthesis-dependent growth. As soon seedlings are exposed to light, fully functional chloroplasts develop, enabling photosynthesis to take place in reactions dependent on light-absorbing pigment molecules present in the thylakoids. Here, light energy is converted to chemical energy, which involves a series of chemical reactions known as the light-dependent reactions. Plants also possess five classes of photoreceptors that are differentially photoactivated based on the type of wavelengths they absorb (from far-red to UV-B). This distinguished capacity of the different photoreceptors to absorb in narrow zone of the solar spectrum allows a precise physiological response in the face of diverse signals from light, such as intensity, quality, and duration. Therefore, throughout plant’s lifespan, we find mechanisms such as the circadian clock, which can be summarized as endogenous genetic pathways that allow plants to anticipate and prepare for environmental daily and seasonal changes. Among the circadian clock, we can find biological processes such as hypocotyl elongation, photosynthesis, stomatal opening, leaf movement, cell cycle progression, and flowering.
Photomorphogenesis, also defined as light-regulated plant development molecular signaling intermediates, are formed in response to the activation of photoreceptors by light. The photomorphogenic physiological responses are germination, de-etiolation, shade avoidance, circadian rhythm, and flowering. Flowering was singled out in this review given its importance for plant multiplication and seed production, which is vital for animal feeding.
The present work intended to show, in a relatively brief way, that the balance between light-promoter development and growth under darkness is delicate and complex, with the capacity to influence all aspects of plant’s life. If we consider the wider view of all life on Earth, having plants as the base of the food chain, it is easy to understand the importance of having a deep knowledge of how light perception is mediated by all photosynthetic organisms.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Karpiński, S.; Szechyńska-Hebda, M.; Wituszyńska, W.; Burdiak, P. Light acclimation, retrograde signalling, cell death and immune defences in plants. Plant Cell Environ. 2013, 36, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Terashima, I.; Hanba, Y.T.; Tazoe, Y.; Vyas, P.; Yano, S. Irradiance and phenotype: Comparative eco-development of sun and shade leaves in relation to photosynthetic CO2 diffusion. J. Exp. Bot. 2006, 57, 343–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kono, M.; Terashima, I. Long-term and short-term responses of the photosynthetic electron transport to fluctuating light. J. Photochem. Photobiol. B Biol. 2014, 137, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light-regulated plant growth and development. In Current Topics in Developmental Biology; Academic Press Inc.: Cambridge, MA, USA, 2010; Volume 91, pp. 29–66. [Google Scholar]
- Higa, T.; Suetsugu, N.; Kong, S.G.; Wada, M. Actin-dependent plastid movement is required for motive force generation in directional nuclear movement in plants. Proc. Natl. Acad. Sci. USA 2014, 111, 4327–4331. [Google Scholar] [CrossRef] [Green Version]
- Yamori, W. Photosynthetic response to fluctuating environments and photoprotective strategies under abiotic stress. J. Plant. Res. 2016, 129, 379–395. [Google Scholar] [CrossRef]
- Murchie, E.H.; Hubbart, S.; Peng, S.; Horton, P. Acclimation of photosynthesis to high irradiance in rice: Gene expression and interactions with leaf development. J. Exp. Bot. 2005, 56, 449–460. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Muchow, R.C. Radiation Use Efficiency. Adv. Agron. 1999, 65, 215–265. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Pierik, R.; Ruban, A.; Wingler, A. The dynamic plant: Capture, transformation, and management of energy. Plant Physiol. 2018, 176, 961–966. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.P. Photosynthesis. Essays Biochem. 2016, 60, 255–273. [Google Scholar] [CrossRef]
- Song, Y.H.; Ito, S.; Imaizumi, T. Similarities in the circadian clock and photoperiodism in plants. Curr. Opin. Plant. Biol. 2010, 13, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Harmer, S.L. The Circadian System in Higher Plants. Ann. Rev. Plant Biol. 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farré, E.M.; Harmer, S.L.; Harmon, F.G.; Yanovsky, M.J.; Kay, S.A. Overlapping and distinct roles of PRR7 and PRR9 in the Arabidopsis circadian clock. Curr. Biol. 2005, 15, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.L.; Newton, L.; Liu, M.J.; Shiu, S.H.; Farré, E.M. A G-box-like motif is necessary for transcriptional regulation by circadian pseudo-response regulators in arabidopsis. Plant Physiol. 2016, 170, 528–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruneda-Paz, J.L.; Breton, G.; Para, A.; Kay, S.A. A functional genomics approach reveals CHE as a component of the Arabidopsis circadian clock. Science 2009, 323, 1481–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, N.; Kita, M.; Ito, S.; Yamashino, T.; Mizuno, T. PSEUDO-RESPONSE REGULATORS, PRR9, PRR7 and PRR5, Together Play Essential Roles Close to the Circadian Clock of Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 686–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, N.; Kiba, T.; Henriques, R.; Mizuno, T.; Chua, N.H.; Sakakibara, H. PSEUDO-RESPONSE ReGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell 2010, 22, 594–605. [Google Scholar] [CrossRef] [Green Version]
- Imaizumi, T.; Tran, H.G.; Swartz, T.E.; Briggs, W.R.; Kay, S.A. FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis. Nature 2003, 426, 302–306. [Google Scholar] [CrossRef]
- Nakamichi, N.; Kita, M.; Niinuma, K.; Ito, S.; Yamashino, T.; Mizoguchi, T.; Mizuno, T. Arabidopsis Clock-Associated Pseudo-Response Regulators PRR9, PRR7 and PRR5 Coordinately and Positively Regulate Flowering Time Through the Canonical CONSTANS-dependent Photoperiodic Pathway. Plant Cell Physiol. 2007, 48. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Bueno, G.; Romero-Campero, F.J.; Lucas-Reina, E.; Romero, J.M.; Valverde, F. Evolution of photoperiod sensing in plants and algae. Curr. Opin. Plant. Biol. 2017, 37, 10–17. [Google Scholar] [CrossRef]
- Song, Y.H.; Estrada, D.A.; Johnson, R.S.; Kim, S.K.; Lee, S.Y.; MacCoss, M.J.; Imaizumi, T. Distinct roles of FKF1, GIGANTEA, and ZEITLUPE proteins in the regulation of constans stability in Arabidopsis photoperiodic flowering. Proc. Natl. Acad. Sci. USA 2014, 111, 17672–17677. [Google Scholar] [CrossRef] [Green Version]
- Fornara, F.; Panigrahi, K.C.S.; Gissot, L.; Sauerbrunn, N.; Rühl, M.; Jarillo, J.A.; Coupland, G. Arabidopsis DOF Transcription Factors Act Redundantly to Reduce CONSTANS Expression and Are Essential for a Photoperiodic Flowering Response. Dev. Cell 2009, 17, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Geng, R.; Gallenstein, R.A.; Somers, D.E. The F-box protein ZEITLUPE controls stability and nucleocytoplasmic partitioning of GIGANTEA. Development 2013, 140, 4060–4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanovsky, M.J.; Kay, S.A. Molecular basis of seasonal time measurement in Arabidopsis. Nature 2002, 419, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Payyavula, R.S.; Chen, L.; Zhang, J.; Zhang, C.; Turgeon, R. FLOWERING LOCUS T mRNA is synthesized in specialized companion cells in Arabidopsis and Maryland Mammoth tobacco leaf veins. Proc. Natl. Acad. Sci. USA 2018, 115, 2830–2835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeger, K.E.; Wigge, P.A. FT Protein Acts as a Long-Range Signal in Arabidopsis. Curr. Biol. 2007, 17, 1050–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbesier, L.; Vincent, C.; Jang, S.; Fornara, F.; Fan, Q.; Searle, I.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C.; et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-Y.; Adams, J.P.; Kim, H.; No, K.; Ma, C.; Strauss, S.H.; Drnevich, J.; Vandervelde, L.; Ellis, J.D.; Rice, B.M.; et al. FLOWERING LOCUS T duplication coordinates reproductive and vegetative growth in perennial poplar. Proc. Natl. Acad. Sci. USA 2011, 108, 10756–10761. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, R.T.; Sheng, X.; Brunner, A.M. Activity of the shoot apical and cambial meristems: Coordination and responses to environmental signals. In Advances in Botanical Research; Academic Press Inc.: Cambridge, MA, USA, 2019; Volume 89, pp. 185–199. ISBN 9780128154656. [Google Scholar]
- Maurya, J.P.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of growth cessation and dormancy in trees: A molecular perspective. Ann. Bot. 2017, 120, 351–360. [Google Scholar] [CrossRef]
- Burgie, E.S.; Vierstra, R.D. Phytochromes: An atomic perspective on photoactivation and signaling. Plant Cell 2014, 26, 568–4583. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, S.E.; Rugnone, M.L.; Kay, S.A. Light Perception: A Matter of Time. Mol. Plant 2020, 13, 363–385. [Google Scholar] [CrossRef]
- Saijo, Y.; Sullivan, J.A.; Wang, H.; Yang, J.; Shen, Y.; Rubio, V.; Ma, L.; Hoecker, U.; Deng, X.W. The COP1-SPA1 interaction defines a critical step in phytochrome A-mediated regulation of HY5 activity. Genes Dev. 2003, 17, 2642–2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Lory, N.; Stauber, J.; Hoecker, U. Photoreceptor Specificity in the Light-Induced and COP1-Mediated Rapid Degradation of the Repressor of Photomorphogenesis SPA2 in Arabidopsis. PLoS Genet. 2015, 11. [Google Scholar] [CrossRef] [Green Version]
- Podolec, R.; Ulm, R. Photoreceptor-mediated regulation of the COP1/SPA E3 ubiquitin ligase. Curr. Opin. Plant Biol. 2018, 45, 18–25. [Google Scholar] [CrossRef]
- Lau, O.S.; Deng, X.W. The photomorphogenic repressors COP1 and DET1: 20 years later. Trends Plant Sci. 2012, 17, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.-G.; Okajima, K. Diverse photoreceptors and light responses in plants. J. Plant Res. 2016, 129, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgie, E.S.; Bussell, A.N.; Walker, J.M.; Dubiel, K.; Vierstra, R.D. Crystal structure of the photosensing module from a red/far-red light-absorbing plant phytochrome. Proc. Natl. Acad. Sci. USA 2014, 111, 10179–10184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, J.M.; Blackwood, L.; Petersen, J.; Sullivan, S. Plant Flavoprotein Photoreceptors. Plant Cell Physiol. 2015, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paik, I.; Huq, E. Plant photoreceptors: Multi-functional sensory proteins and their signaling networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Lau, O.S.; Deng, X.W. Plant hormone signaling lightens up: Integrators of light and hormones. Curr. Opin. Plant Biol. 2010, 13, 571–577. [Google Scholar] [CrossRef]
- Leivar, P.; Quail, P.H. PIFs: Pivotal components in a cellular signaling hub. Trends Plant Sci. 2011, 16, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Sakuraba, Y.; Jeong, J.; Kang, M.Y.; Kim, J.; Paek, N.C.; Choi, G. Phytochrome-interacting transcription factors PIF4 and PIF5 induce leaf senescence in Arabidopsis. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leivar, P.; Monte, E. PIFs: Systems integrators in plant development. Plant Cell 2014, 26, 56–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvão, V.C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Ouyang, X.; Deng, X.W. Beyond repression of photomorphogenesis: Role switching of COP/DET/FUS in light signaling. Curr. Opin. Plant Biol. 2014, 21, 96–103. [Google Scholar] [CrossRef]
- Medzihradszky, M.; Bindics, J.; Ádám, É.; Viczián, A.; Klement, É.; Lorrain, S.; Gyula, P.; Mérai, Z.; Fankhauser, C.; Medzihradszky, K.F.; et al. Phosphorylation of phytochrome B inhibits light-induced signaling via accelerated dark reversion in Arabidopsis. Plant Cell 2013, 25, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Galvão, R.M.; Li, M.; Burger, B.; Bugea, J.; Bolado, J.; Chory, J. Arabidopsis HEMERA/pTAC12 Initiates photomorphogenesis by phytochromes. Cell 2010, 141, 1230–1240. [Google Scholar] [CrossRef] [Green Version]
- Kaiserli, E.; Páldi, K.; O’Donnell, L.; Batalov, O.; Pedmale, U.V.; Nusinow, D.A.; Kay, S.A.; Chory, J. Integration of Light and Photoperiodic Signaling in Transcriptional Nuclear Foci. Dev. Cell 2015, 35, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Van Buskirk, E.K.; Decker, P.V.; Chen, M. Photobodies in light signaling. Plant Physiol. 2012, 158, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, R.; Nakamura, M.; Mochizuki, N.; Kay, S.A.; Nagatani, A. Light-dependent translocation of a phytochrome B-GFP fusion protein to the nucleus in transgenic Arabidopsis. J. Cell Biol. 1999, 145, 437–445. [Google Scholar] [CrossRef]
- Lin, C.; Ahmad, M.; Cashmore, A.R. Arabidopsis cryptochrome 1 is a soluble protein mediating blue light-dependent regulation of plant growth and development. Plant J. 1996, 10, 893–902. [Google Scholar] [CrossRef]
- Yu, X.; Klejnot, J.; Zhao, X.; Shalitin, D.; Maymon, M.; Yang, H.; Lee, J.; Liu, X.; Lopez, J.; Lina, C. Arabidopsis cryptochrome 2 completes its posttranslational life cycle in the nucleus. Plant Cell 2007, 19, 3146–3156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, I.; Pokorny, R.; Byrdin, M.; Hoang, N.; Ritz, T.; Brettel, K.; Essen, L.-O.; van der Horst, G.T.J.; Batschauer, A.; Ahmad, M. The Cryptochromes: Blue Light Photoreceptors in Plants and Animals. Annu. Rev. Plant. Biol. 2011, 62, 335–364. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Liu, H.; Liu, B.; Liu, X.; Lin, C. Blue light-dependent interaction of CRY2 with SPA1 regulates COP1 activity and floral initiation in arabidopsis. Curr. Biol. 2011, 21, 841–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Zuo, Z.; Liu, H.; Liu, X.; Lin, C. Arabidopsis cryptochrome 1 interacts with SPA1 to suppress COP1 activity in response to blue light. Genes Dev. 2011, 25, 1029–1034. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, X.; Li, K.; Liu, H.; Lin, C. Multiple bHLH Proteins form Heterodimers to Mediate CRY2-Dependent Regulation of Flowering-Time in Arabidopsis. PLoS Genet. 2013, 9, e1003861. [Google Scholar] [CrossRef] [Green Version]
- Lian, H.L.; He, S.B.; Zhang, Y.C.; Zhu, D.M.; Zhang, J.Y.; Jia, K.P.; Sun, S.X.; Li, L.; Yang, H.Q. Blue-light-dependent interaction of cryptochrome 1 with SPA1 defines a dynamic signaling mechanism. Genes Dev. 2011, 25, 1023–1028. [Google Scholar] [CrossRef] [Green Version]
- Demarsy, E.; Fankhauser, C. Higher plants use LOV to perceive blue light. Curr. Opin. Plant Biol. 2009, 12, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Suetsugu, N.; Wada, M. Evolution of Three LOV Blue Light Receptor Families in Green Plants and Photosynthetic Stramenopiles: Phototropin, ZTL/FKF1/LKP2 and Aureochrome. Plant Cell Physiol. 2013, 54, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.-I.; Kinoshita, T.; Matsumoto, M.; Nakayama, K.I.; Doi, M.; Shimazaki, K.-I. Blue light-induced autophosphorylation of phototropin is a primary step for signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 5626–5631. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Oses-Prieto, J.A.; Kutschera, U.; Tseng, T.S.; Hao, L.; Burlingame, A.L.; Wang, Z.Y.; Briggs, W.R. Blue light-induced proteomic changes in etiolated Arabidopsis seedlings. J. Proteome Res. 2014, 13, 2524–2533. [Google Scholar] [CrossRef]
- Motchoulski, A.; Liscum, E. Arabidopsis NPH3: A NPH1 photoreceptor-interacting protein essential for phototropism. Science 1999, 286, 961–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esmon, C.A.; Tinsley, A.G.; Ljung, K.; Sandberg, G.; Hearne, L.B.; Liscum, E. A gradient of auxin and auxin-dependent transcription precedes tropic growth responses. Proc. Natl. Acad. Sci. USA 2006, 103, 236–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Li, J.; Qu, L.; Hager, J.; Chen, Z.; Zhao, H.; Deng, X.W. Light Control of Arabidopsis Development Entails Coordinated Regulation of Genome Expression and Cellular Pathways. Plant Cell 2001, 13, 2589–2607. [Google Scholar] [CrossRef] [PubMed]
- Inada, S.; Ohgishi, M.; Mayama, T.; Okada, K.; Sakai, T. RPT2 is a signal transducer involved in phototropic response and stomatal opening by association with phototropin 1 in Arabidopsis thaliana. Plant Cell 2004, 16, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Baudry, A.; Ito, S.; Song, Y.H.; Strait, A.A.; Kiba, T.; Lu, S.; Henriques, R.; Pruneda-Paz, J.L.; Chua, N.H.; Tobin, E.M.; et al. F-Box proteins FKF1 and LKP2 act in concert with ZEITLUPE to control Arabidopsis clock progression. Plant Cell 2010, 22, 606–622. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.C.; Lasswell, J.; Rogg, L.E.; Cohen, M.A.; Bartel, B. FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis. Cell 2000, 101, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Fujiwara, S.; Somers, D.E. PRR5 regulates phosphorylation, nuclear import and subnuclear localization of TOC1 in the Arabidopsis circadian clock. EMBO J. 2010, 29, 1903–1915. [Google Scholar] [CrossRef] [Green Version]
- Andrade, M.A.; González-Guzmán, M.; Serrano, R.; Rodríguez, P.L. A combination of the F-box motif and kelch repeats defines a large Arabidopsis family of F-box proteins. Plant Mol. Biol. 2001, 46, 603–614. [Google Scholar] [CrossRef]
- Imaizumi, T.; Schultz, T.; Harmon, F.; Ho, L.A.; Kay, A.S. FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science 2005, 309, 293–297. [Google Scholar] [CrossRef]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schäfer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Tossi, V.; Lamattina, L.; Jenkins, G.I.; Cassia, R.O. Ultraviolet-B-induced stomatal closure in arabidopsis is regulated by the UV RESISTANCE LOCUS8 photoreceptor in a nitric oxide-dependent mechanism. Plant Physiol. 2014, 164, 2220–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehér, B.; Kozma-Bognár, L.; Kevei, É.; Hajdu, A.; Binkert, M.; Davis, S.J.; Schäfer, E.; Ulm, R.; Nagy, F. Functional interaction of the circadian clock and UV RESISTANCE LOCUS 8-controlled UV-B signaling pathways in Arabidopsis thaliana. Plant J. 2011, 67, 37–48. [Google Scholar] [CrossRef]
- Wu, D.; Hu, Q.; Yan, Z.; Chen, W.; Yan, C.; Huang, X.; Zhang, J.; Yang, P.; Deng, H.; Wang, J.; et al. Structural basis of ultraviolet-B perception by UVR8. Nature 2012, 484, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Cloix, C.; Kaiserli, E.; Heilmann, M.; Baxter, K.J.; Brown, B.A.; O’Hara, A.; Smith, B.O.; Christie, J.M.; Jenkins, G.I. C-terminal region of the UV-B photoreceptor UVR8 initiates signaling through interaction with the COP1 protein. Proc. Natl. Acad. Sci. USA 2012, 109, 16366–16370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binkert, M.; Kozma-Bognár, L.; Terecskei, K.; De Veylder, L.; Nagy, F.; Ulm, R. UV-B-Responsive association of the Arabidopsis bZIP transcription factor ELONGATED HYPOCOTYL5 with target genes, including its own promoter. Plant Cell 2014, 26, 4200–4213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heijde, M.; Ulm, R. UV-B photoreceptor-mediated signalling in plants. Trends Plant. Sci. 2012, 17, 230–237. [Google Scholar] [CrossRef]
- Ulm, R.; Nagy, F. Signalling and gene regulation in response to ultraviolet light. Curr. Opin. Plant. Biol. 2005, 8, 477–482. [Google Scholar] [CrossRef]
- Ballaré, C.L. Light Regulation of Plant Defense. Annu. Rev. Plant. Biol. 2014, 65, 335–363. [Google Scholar] [CrossRef]
- Hideg, É.; Jansen, M.A.K.; Strid, Å. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Lau, O.S.; Deng, X.W. Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 2007, 8, 217–230. [Google Scholar] [CrossRef]
- Ni, W.; Xu, S.L.; Tepperman, J.M.; Stanley, D.J.; Maltby, D.A.; Gross, J.D.; Burlingame, A.L.; Wang, Z.Y.; Quail, P.H. A mutually assured destruction mechanism attenuates light signaling in Arabidopsis. Science 2014, 344, 1160–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Ni, W.; Yu, R.; Deng, X.W.; Chen, H.; Wei, N. Light-Dependent Degradation of PIF3 by SCFEBF1/2 Promotes a Photomorphogenic Response in Arabidopsis. Curr. Biol. 2017, 27, 2420–2430. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Xu, S.L.; González-Grandío, E.; Chalkley, R.J.; Huhmer, A.F.R.; Burlingame, A.L.; Wang, Z.Y.; Quail, P.H. PPKs mediate direct signal transfer from phytochrome photoreceptors to transcription factor PIF3. Nat. Commun. 2017, 8, 15236. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Kim, J.; Park, E.; Kim, J.I.; Kang, C.; Choi, G. PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis thaliana. Plant Cell 2004, 16, 3045–3058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majee, M.; Kumar, S.; Kathare, P.K.; Wu, S.; Gingerich, D.; Nayak, N.R.; Salaita, L.; Dinkins, R.; Martin, K.; Goodin, M.; et al. KELCH F-BOX protein positively influences Arabidopsis seed germination by targeting PHYTOCHROME-INTERACTING FACTOR1. Proc. Natl. Acad. Sci. USA 2018, 115, E4120–E4129. [Google Scholar] [CrossRef] [PubMed]
- Vaistij, F.E.; Barros-Galvão, T.; Cole, A.F.; Gilday, A.D.; He, Z.; Li, Y.; Harvey, D.; Larson, T.R.; Graham, I.A. MOTHER-OF-FT-AND-TFL1 represses seed germination under far-red light by modulating phytohormone responses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2018, 115, 8442–8447. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Yamaguchi, S.; Hu, J.; Yusuke, J.; Jung, B.; Paik, I.; Lee, H.S.; Sun, T.P.; Kamiya, Y.; Choi, G. PIL5, a phytochrome-interacting bHLH protein, regulates gibberellin responsiveness by binding directly to the GAI and RGA promoters in Arabidopsis seeds. Plant Cell 2007, 19, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.K.; Yamaguchi, S.; Lim, S.; Oh, E.; Park, J.; Hanada, A.; Kamiya, Y.; Choi, G. SOMNUS, a CCCH-type zinc finger protein in Arabidopsis, negatively regulates light-dependent seed germination downstream of PIL5. Plant Cell 2008, 20, 1260–1277. [Google Scholar] [CrossRef] [Green Version]
- Parcy, F. Flowering: A time for integration. Int. J. Dev. Biol. 2005, 49, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.S.; Imaizumi, T. Circadian clock and photoperiodic response in arabidopsis: From seasonal flowering to redox homeostasis. Biochemistry 2015, 54, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 2007, 318, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.H.; Smith, R.W.; To, B.J.; Millar, A.J.; Imaizumi, T. FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering. Science 2012, 336, 1045–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomoto, Y.; Kubozono, S.; Yamashino, T.; Nakamichi, N.; Mizuno, T. Circadian Clock- and PIF4-Controlled Plant Growth: A Coincidence Mechanism Directly Integrates a Hormone Signaling Network into the Photoperiodic Control of Plant Architectures in Arabidopsis thaliana. Plant Cell Physiol. 2012, 53, 1950–1964. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.S.; Kubota, A.; Imaizumi, T. Circadian clock and photoperiodic flowering in arabidopsis: CONSTANS is a Hub for Signal integration. Plant Physiol. 2017, 173, 5–15. [Google Scholar] [CrossRef]
- Ito, S.; Song, Y.H.; Josephson-Day, A.R.; Miller, R.J.; Breton, G.; Olmstead, R.G.; Imaizumi, T. FLOWERING BHLH transcriptional activators control expression of the photoperiodic flowering regulator CONSTANS in Arabidopsis. Proc. Natl. Acad Sci. USA 2012, 109, 3582–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Marchal, V.; Panigrahi, K.C.S.; Wenkel, S.; Soppe, W.; Deng, X.W.; Valverde, F.; Coupland, G. Arabidopsis COP1 shapes the temporal pattern of CO accumulation conferring a photoperiodic flowering response. EMBO J. 2008, 27, 1277–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazaro, A.; Valverde, F.; Piñ, M.; Jarillo, J.A. The Arabidopsis E3 Ubiquitin Ligase HOS1 Negatively Regulates CONSTANS Abundance in the Photoperiodic Control of Flowering W. Plant Cell 2012, 24, 982–999. [Google Scholar] [CrossRef] [Green Version]
- Valverde, F.; Mouradov, A.; Soppe, W.; Ravenscroft, D.; Samach, A.; Coupland, G. Photoreceptor Regulation of CONSTANS Protein in Photoperiodic Flowering. Science 2004, 303, 1003–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, Q.; Liu, Y.; Zhao, X.; Imaizumi, T.; Somers, D.E.; Tobin, E.M.; Lin, C. Arabidopsis CRY2 and ZTL mediate blue-light regulation of the transcription factor CIB1 by distinct mechanisms. Proc. Natl. Acad. Sci. USA 2013, 110, 17582–17587. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Liu, C.; Shen, L.; Wu, Y.; Chen, H.; Robertson, M.; Helliwell, C.A.; Ito, T.; Meyerowitz, E.; Yu, H. A Repressor Complex Governs the Integration of Flowering Signals in Arabidopsis. Dev. Cell 2008, 15, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Le, C.; Wang, Y.; Li, Z.; Jiang, D.; Wang, Y.; He, Y. Arabidopsis FLC clade members form flowering-repressor complexes coordinating responses to endogenous and environmental cues. Nat. Commun. 2013, 4, 1–10. [Google Scholar] [CrossRef]
- Castillejo, C.; Pelaz, S. The Balance between CONSTANS and TEMPRANILLO Activities Determines FT Expression to Trigger Flowering. Curr. Biol. 2008, 18, 1338–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, J.; Yant, L.J.; Mürdter, F.; Küttner, F.; Schmid, M. Repression of flowering by the miR172 target SMZ. PLoS Biol. 2009, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Yu, X.; Li, K.; Klejnot, J.; Yang, H.; Lisiero, D.; Lin, C. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 2008, 322, 1535–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
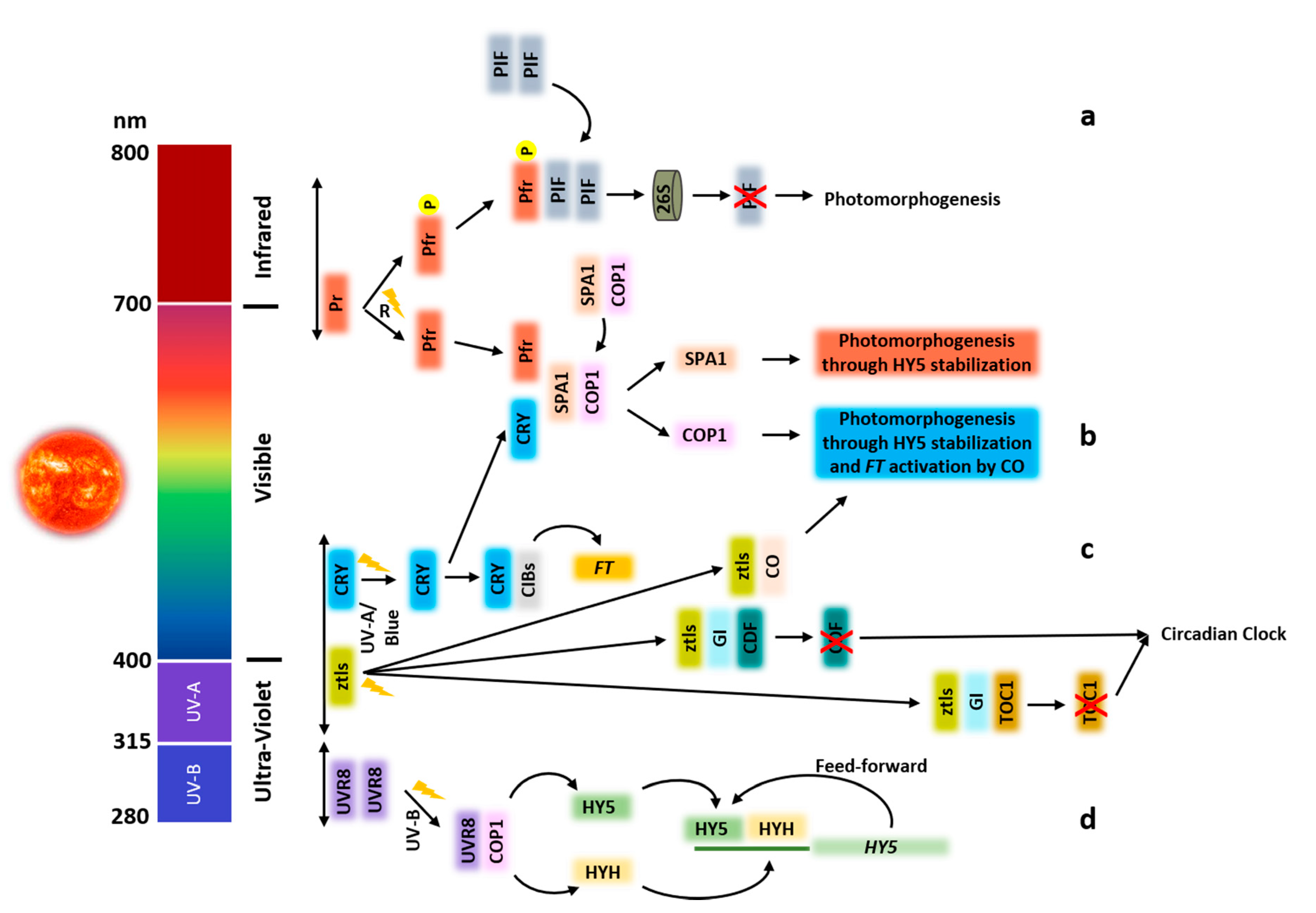
Figure 1.
Proposed explanatory model of plants photoreceptors activity upon activated by light. (a) The biological active Pfr form of the phytochrome is activated by red light (R). The interaction between Pfr and PIFs triggers the rapid phosphorylation of PIFs entering protein degradation via the ubiquitin 26S proteasome pathway promoting photomorphogenesis. Pfr also induces COP1-SPA1 complex dissociation. All these pathways lead to the accumulation of major transcription factors (TFs) triggering photomorphogenesis such as HY5 (b) Two mechanisms are represented for cryptochrome signal transduction. Transcription regulation of light-activated Cry after interaction with TF CIB1 and its relatives (CIBs) leads to the activation of FT transcription, promoting floral initiation. The other mechanisms involve the interaction of cryptochromes with SPA1 proteins to suppress SPA1 activation of COP1 activity necessary for the degradation of HY5, HYH, CO, and other transcription regulators promoting photomorphogenesis. (c) ztls are activated by UV-A/blue light. Once activated, they interact with GI, resulting in the stabilization of these receptors, enabling them to act as E3 ligases which target the transcriptional regulators CDF and TOC1 for degradation. In addition, light-activated FKF1 interacts with and stabilizes CO. (d) The inactive dimeric form of UVR8 is activated to the monomeric form by UV-B, which interacts with the E3 ubiquitin ligase COP1. With the suppression of COP1, HY5 and HYH are stabilized. These two proteins, in a feed-forward loop, bind to the HY5 promoter region with further activation of transcription.
Figure 1.
Proposed explanatory model of plants photoreceptors activity upon activated by light. (a) The biological active Pfr form of the phytochrome is activated by red light (R). The interaction between Pfr and PIFs triggers the rapid phosphorylation of PIFs entering protein degradation via the ubiquitin 26S proteasome pathway promoting photomorphogenesis. Pfr also induces COP1-SPA1 complex dissociation. All these pathways lead to the accumulation of major transcription factors (TFs) triggering photomorphogenesis such as HY5 (b) Two mechanisms are represented for cryptochrome signal transduction. Transcription regulation of light-activated Cry after interaction with TF CIB1 and its relatives (CIBs) leads to the activation of FT transcription, promoting floral initiation. The other mechanisms involve the interaction of cryptochromes with SPA1 proteins to suppress SPA1 activation of COP1 activity necessary for the degradation of HY5, HYH, CO, and other transcription regulators promoting photomorphogenesis. (c) ztls are activated by UV-A/blue light. Once activated, they interact with GI, resulting in the stabilization of these receptors, enabling them to act as E3 ligases which target the transcriptional regulators CDF and TOC1 for degradation. In addition, light-activated FKF1 interacts with and stabilizes CO. (d) The inactive dimeric form of UVR8 is activated to the monomeric form by UV-B, which interacts with the E3 ubiquitin ligase COP1. With the suppression of COP1, HY5 and HYH are stabilized. These two proteins, in a feed-forward loop, bind to the HY5 promoter region with further activation of transcription.

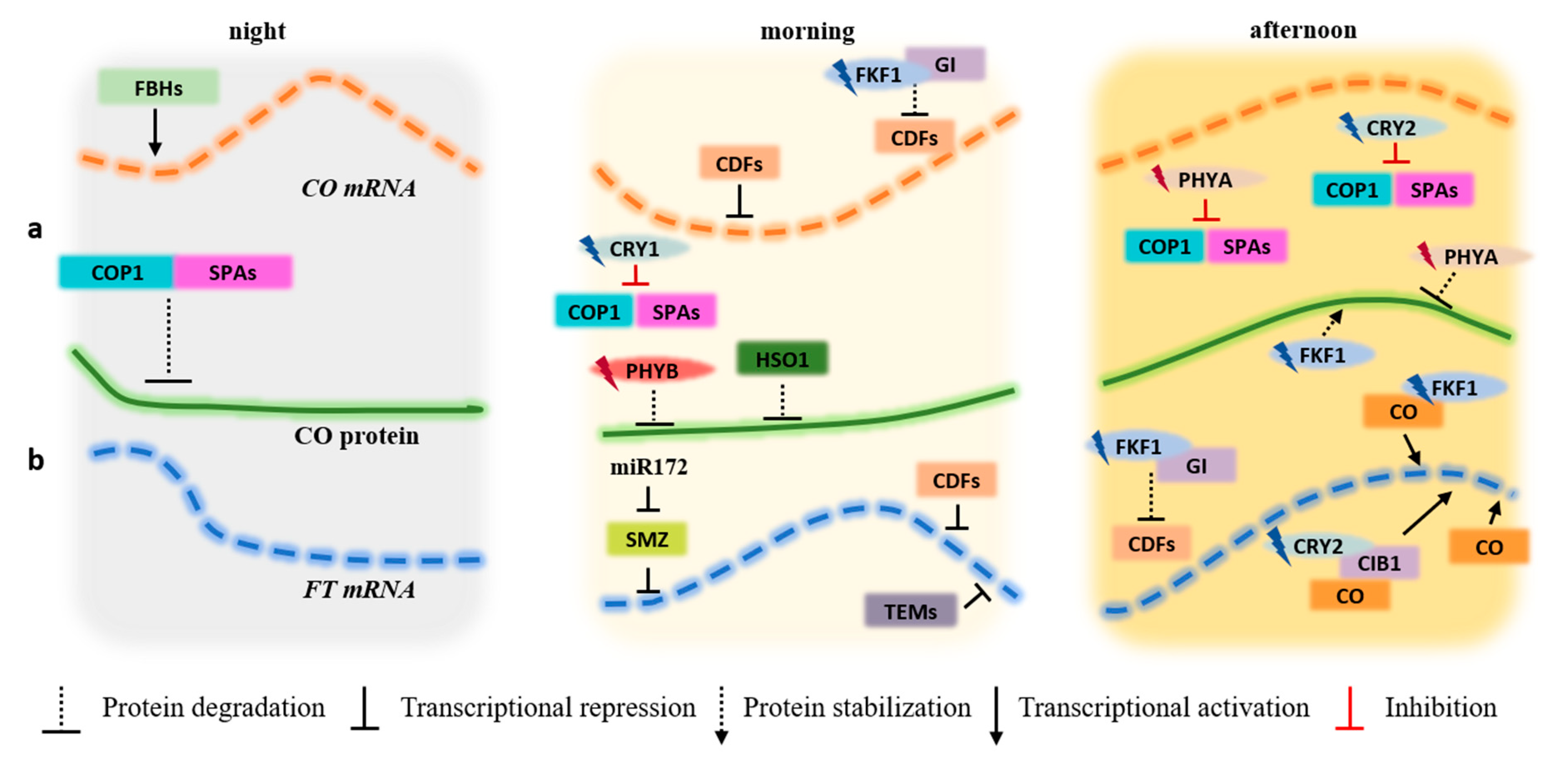
Figure 2.
CO and FT photoperiodic regulation under LD during the night, morning, and afternoon periods. (a) Transcriptional (CO mRNA) and post-translational (CO protein) regulation. (b) Transcriptional regulation of FT (FT mRNA). At night, the FBHs (bHLH transcription factors) bind to the CO promoter, activating its transcription. In the morning, the presence of high amounts of CDF proteins on the CO and FT promoters repress the transcription of both genes. As afternoon is reached, FKF1 is photoactivated by blue light. At this point, it forms a complex with GI, promoting the degradation of the CDFs proteins which bind to the CO and FT promoters, freeing these promoter regions for binding to other gene activators. At the protein regulation level, the COP1-SPAs complex degrades CO protein during the night hours. When CRY1 and CRY2 are photoexcited by blue light, they acquire the capacity to interact with COP1 and SPAs, inhibiting the COP1-SPAs complex activity and increasing the stability of CO. PHYA also inhibits the COP1-SPAs complex. During the first hours of the day, HOS1, ubiquitin ligase, and red light activated PHYB mediate CO degradation. By the time CO picks late in the afternoon, blue light-activated FKF1 and far-red photo-activated PHYA stabilize CO. The amount of CO protein in the morning is low due to interactions with other proteins that prevents the CO-dependent activation of FT. Throughout the day, as CO becomes stable upon interaction with blue light-activated FKF1 and CRY2, activates the transcription of FT gene by directly binding to the FT promoter. The blue light-activated form of CRY2 also interacts with CIB1 and directly binds to the FT promoter, activating the transcription of the gene late in the afternoon. miR172 is a negative regulator of SMZ and SMZ protein and is a powerful repressor of FT expression under LD. TEMs transcription factors (TEM1 and TEM2) bind to FT promoter, inhibiting its transcription in the morning.
Figure 2.
CO and FT photoperiodic regulation under LD during the night, morning, and afternoon periods. (a) Transcriptional (CO mRNA) and post-translational (CO protein) regulation. (b) Transcriptional regulation of FT (FT mRNA). At night, the FBHs (bHLH transcription factors) bind to the CO promoter, activating its transcription. In the morning, the presence of high amounts of CDF proteins on the CO and FT promoters repress the transcription of both genes. As afternoon is reached, FKF1 is photoactivated by blue light. At this point, it forms a complex with GI, promoting the degradation of the CDFs proteins which bind to the CO and FT promoters, freeing these promoter regions for binding to other gene activators. At the protein regulation level, the COP1-SPAs complex degrades CO protein during the night hours. When CRY1 and CRY2 are photoexcited by blue light, they acquire the capacity to interact with COP1 and SPAs, inhibiting the COP1-SPAs complex activity and increasing the stability of CO. PHYA also inhibits the COP1-SPAs complex. During the first hours of the day, HOS1, ubiquitin ligase, and red light activated PHYB mediate CO degradation. By the time CO picks late in the afternoon, blue light-activated FKF1 and far-red photo-activated PHYA stabilize CO. The amount of CO protein in the morning is low due to interactions with other proteins that prevents the CO-dependent activation of FT. Throughout the day, as CO becomes stable upon interaction with blue light-activated FKF1 and CRY2, activates the transcription of FT gene by directly binding to the FT promoter. The blue light-activated form of CRY2 also interacts with CIB1 and directly binds to the FT promoter, activating the transcription of the gene late in the afternoon. miR172 is a negative regulator of SMZ and SMZ protein and is a powerful repressor of FT expression under LD. TEMs transcription factors (TEM1 and TEM2) bind to FT promoter, inhibiting its transcription in the morning.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Teixeira, R.T. Distinct Responses to Light in Plants. Plants 2020, 9, 894. https://doi.org/10.3390/plants9070894
AMA Style
Teixeira RT. Distinct Responses to Light in Plants. Plants. 2020; 9(7):894. https://doi.org/10.3390/plants9070894
Chicago/Turabian StyleTeixeira, Rita Teresa. 2020. "Distinct Responses to Light in Plants" Plants 9, no. 7: 894. https://doi.org/10.3390/plants9070894
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.