Silicon Supply Improves Leaf Gas Exchange, Antioxidant Defense System and Growth in Saccharum officinarum Responsive to Water Limitation

 ,
,  ,
,  ,
,  ,
,
Abstract
:
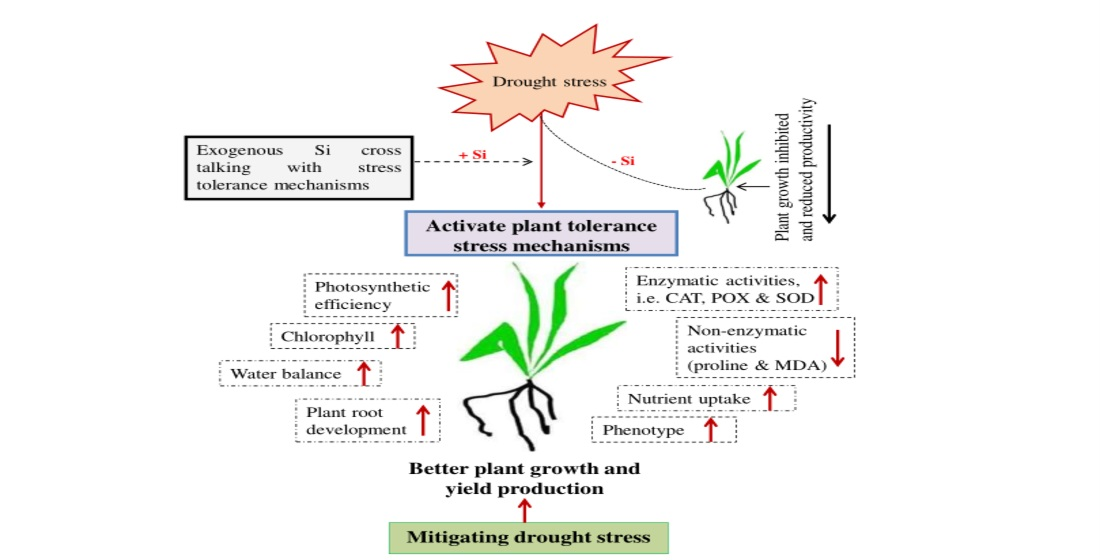
1. Introduction
2. Materials and Methods
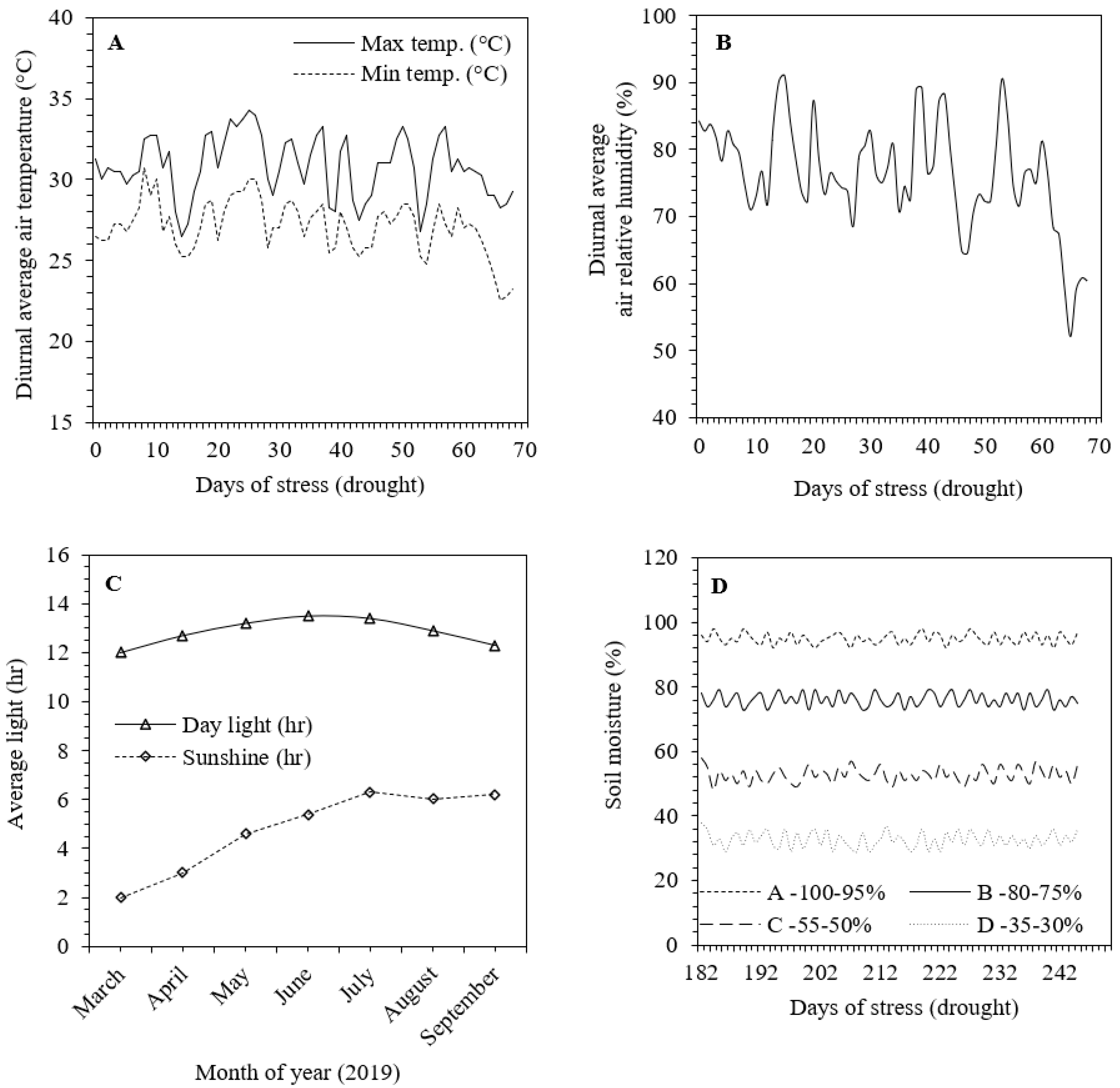
2.1. Experimental Site and Treatment Design
2.2. Determination of Relative Water Content, Chlorophyll and Chlorophyll Stability Index
2.3. Photosynthetic Characteristics
2.4. Measurement of EnzymeActivities
2.5. Determination of Proline and Malondialdehyde Content
2.6. Determination of Silicon Content
2.7. Measurement of Growth and Biomass Traits
2.8. The Model
2.9. Governing Equation
2.10. Solution
2.11. Statistical Analysis
3. Results
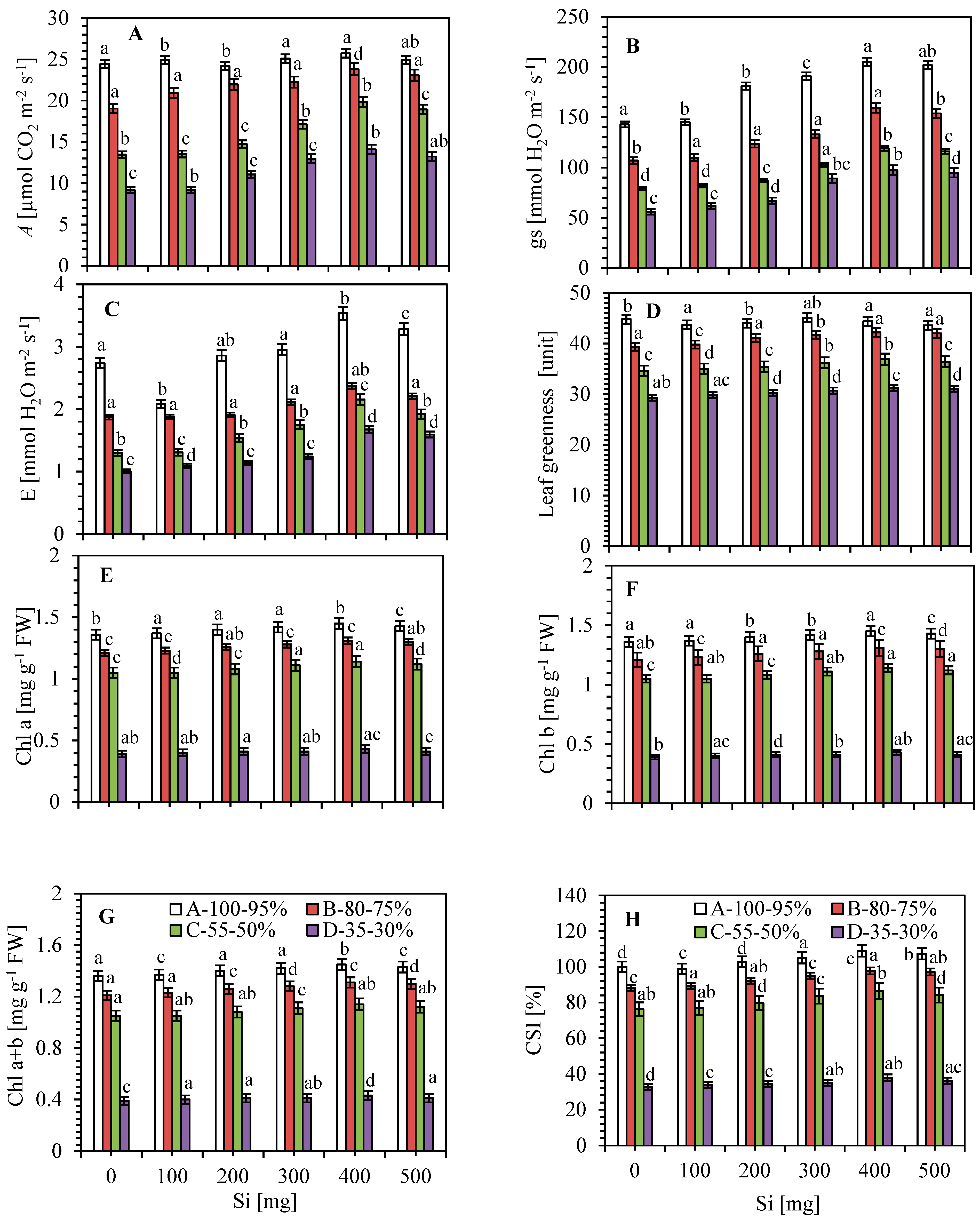
3.1. Impact of Silicon on Growth and Biomass under Limited Water Irrigation
3.2. Impact of Leaf Gas Exchange Characteristics
3.3. Photosynthetic Pigments, Leaf Greenness (SPAD Value) and Relative Water Content
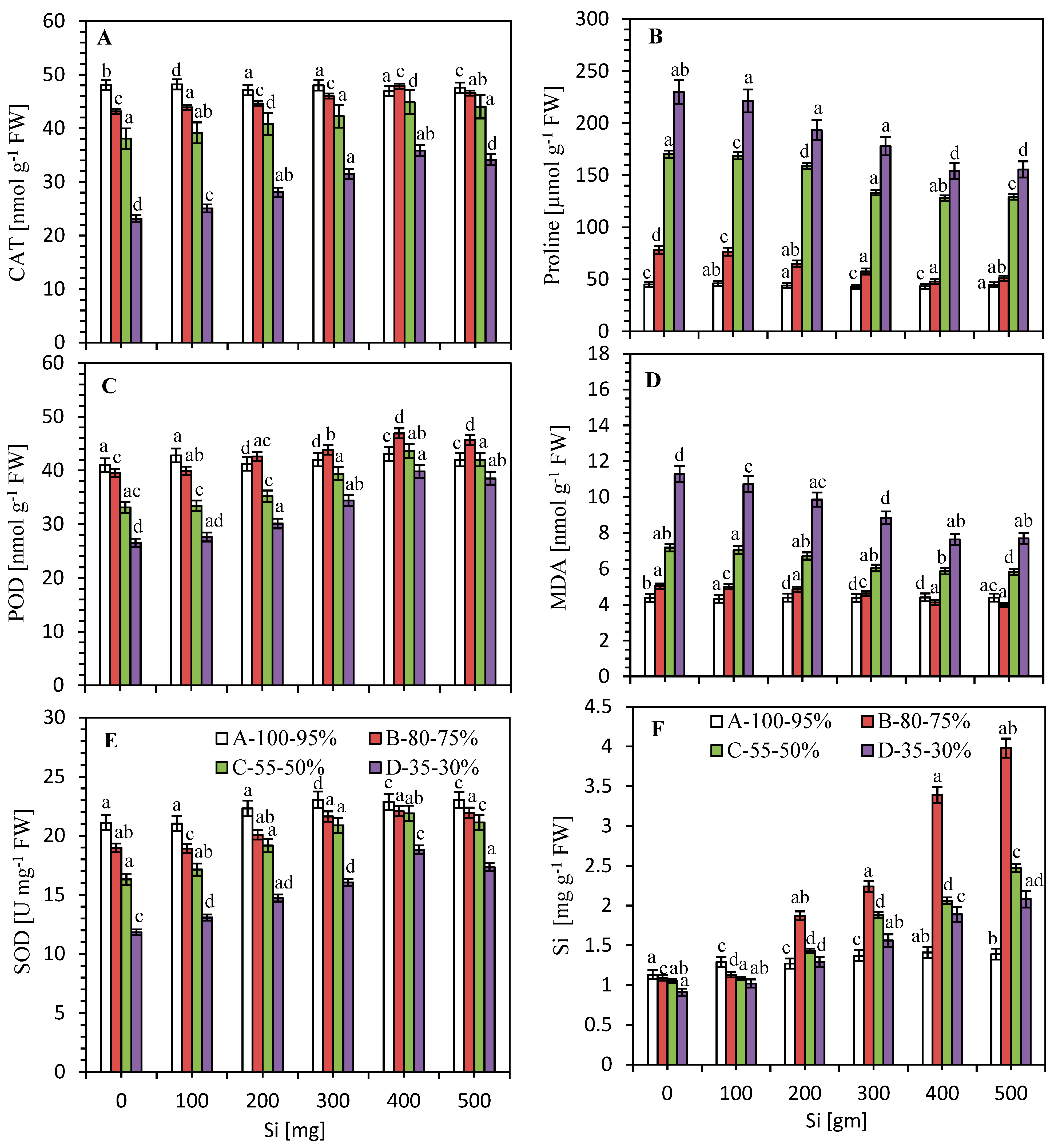
3.4. Impact of Si on the Activities of Antioxidants against Limited Water Irrigation
3.5. Impact of Si on Proline and MDA Content
3.6. Impact of Silicon on Si Accumulation under Limited Water Irrigation
3.7. Regression Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, D.; Wang, S.; Yin, L.; Deng, X. How does silicon mediate plant water uptake and loss under water deficiency? Front. Plant Sci. 2018, 9, 281. [Google Scholar] [CrossRef]
- Verma, K.K.; Liu, X.H.; Wu, K.C.; Singh, R.K.; Song, Q.Q.; Malviya, M.K.; Song, X.P.; Singh, P.; Verma, C.L.; Li, Y.R. The impact of silicon on photosynthetic and biochemical responses of sugarcane under different soil moisture levels. Silicon 2019. [Google Scholar] [CrossRef]
- Vilela, R.D.; Bezerra, B.K.L.; Froehlich, A.; Endres, L. Antioxidant system is essential to increase drought tolerance of sugarcane. Ann. Appl. Biol. 2017, 171, 451–463. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef] [PubMed]
- United Nations. World Population Prospects: Highlights. 2019. Available online: https://www.un.org/development/desa/publications/world-population-prospects-2019highlights.html (accessed on 30 March 2020).
- Sahebi, M.; Hanafi, M.M.; Akmar, A.S.N.; Rafii, M.Y.; Azizi, P.; Tengoua, F.F.; Azwa, J.N.M.; Shabanimofrad, M. Importance of silicon and mechanisms of biosilica formation in plants. BioMed Res. Int. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K. Abiotic stress responses and microbe mediated mitigation in plants: The Omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef]
- Rasool, S.; Hameed, A.; Azooz, M.; Siddiqi, T.; Ahmad, P. Salt Stress: Causes, Types and Responses of Plants. In Ecophysiology and Responses of Plants under Salt Stress; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–24. [Google Scholar]
- Bodner, G.; Nakhforoosh, A.; Kaul, H.P. Management of crop water under drought: A review. Agron. Sustain. Dev. 2015, 35, 401–442. [Google Scholar] [CrossRef]
- Verma, K.K.; Singh, R.K.; Song, Q.Q.; Singh, P.; Zhang, B.Q.; Song, X.P.; Chen, G.L.; Li, Y.R. Silicon alleviates drought stress of sugarcane plants by improving antioxidant responses. Biomed. J. Sci. Tech. Res. 2019, 17, 002957. [Google Scholar]
- Verma, K.K.; Wu, K.-C.; Singh, P.; Malviya, M.K.; Singh, R.K.; Song, X.-P.; Li, Y.R. The protective role of silicon in sugarcane under water stress: Photosynthesis and antioxidant enzymes. Biomed. J. Sci. Tech. Res. 2019, 15, 002685. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, N.O.; Medici, L.O.; Olivares, F.L.; Dobbss, L.B.; Torres-Netto, A.; Silva, S.F.; Canellas, L.P. Metabolic profile and antioxidant responses during drought stress recovery in sugarcane treated with humic acids and endophytic diazotrophic bacteria. Ann. Appl. Biol. 2016, 168, 203–213. [Google Scholar] [CrossRef]
- Bassi, B.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.O.S.; Cunhaa, M.P.; Jesusa, C.D.F.; Rochaa, G.J.M.; Pradellaa, J.G.C.; Rossella, C.E.V.; Filhoab, R.M.; Bonomia, A. Second generation ethanol in Brazil: Can it compete with electricity production? Bioresour. Technol. 2011, 102, 8964–8971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, S.C.; Maehara, L.; Machado, C.M.M.; Farinas, C.S. 2G ethanol from the whole sugarcane lignocellulosic biomass. Biotechnol. Biofuels 2015, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Belanger, R.R. The controversies of silicon’s role in plants biology. New Phytol. 2019, 221, 67–85. [Google Scholar] [CrossRef]
- Griffin, M.; Hogan, B.; Schmidt, O. Silicon reduces slug feeding on wheat seedlings. J. Pest Sci. 2015, 88, 17–24. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Dallagnol, L.J.; Duarte, H.S.S.; Datnoff, L.E. Silicon Control of Foliar Diseases in Monocots and Dicots. In Silicon and Plant Diseases; Rodrigues, F.A., Datnoff, L.E., Eds.; Springer: Cham, The Netherlands, 2015; pp. 67–108. [Google Scholar]
- Reynolds, O.L.; Padula, M.P.; Zeng, R.; Gurr, G.M. Silicon: Potential to promote direct and indirect effects on plant defense against arthropod pests in agriculture. Front. Plant Sci. 2016, 7, 744. [Google Scholar] [CrossRef] [Green Version]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates: Sunderland, UK, 2005. [Google Scholar]
- Liang, Y.; Nikolic, M.; Belanger, R.; Gong, H.; Song, A. Effect of silicon on crop growth, yield and quality. In Silicon in agriculture: From Theory to Practice; Liang, Y., Nikolic, M., Belanger, R., Gong, H., Song, A., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 209–233. [Google Scholar]
- Epstein, E. Silicon: Its manifold roles in plants. Ann. Appl. Biol. 2009, 155, 155–160. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Mitani, N. Transport of silicon from roots to panicles in plants. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2011, 87, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, S.; Dillenburg, L.C. Measurements of leaf relative water content in Araucaria angustifolia. Rev. Brasilian Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop. Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Azevedo, M.M.; Carvalho, A.; Pascoal, C.; Rodrigues, F.; Caassio, F. Responses of antioxidant defenses to Cu and Zn stress in two aquatic fungi. Sci. Total. Environ. 2007, 377, 233–243. [Google Scholar] [CrossRef]
- Bai, L.; Sui, F.; Ge, T.; Sun, Z.; Lu, Y.; Zhou, G. Effect of soil drought stress on leaf water status, membrane permeability and enzymatic antioxidant system of maize. Pedosphere 2006, 16, 326–332. [Google Scholar] [CrossRef]
- Esposito, J.B.; Esposito, B.P.; Azevedo, R.A.; Cruz, L.S.; da Silva, L.C.; de Souza, S.R. Protective effect of Mn (III)-desferrioxamine B upon oxidative stress caused by ozone and acid rain in the Brazilian soybean cultivar Glycine max ‘Sambaiba’. Environ. Sci. Pollut. Res. Int. 2015, 22, 5315–5324. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Come, D. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiol. Plant. 1996, 97, 104–110. [Google Scholar] [CrossRef]
- Wang, L.; Huang, X.; Zhou, Q. Effects of rare earth elements on the distribution of mineral elements and heavy metals in horseradish. Chemosphere 2008, 73, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Ahmed, M.; Fayyaz-Ul-Hassen; Khurshid, Y. Does silicon and irrigation have impact on drought tolerance mechanism of sorghum? Agric. Water Manag. 2011, 98, 1808–1812. [Google Scholar] [CrossRef]
- Hameed, A.; Sheikh, M.A.; Jamil, A.; Basra, S.M.A. Seed priming with sodium silicate enhances seed germination and seedling growth in wheat (Triticum aestivum L.) under water deficit stress induced by polyethylene glycol. Pak. J. Life Soc. Sci. 2013, 11, 19–24. [Google Scholar]
- Hattori, T.; Inanaga, S.; Tanimoto, E.; Lux, A.; Luxova, M.; Sugimoto, Y. Silicon-induced changes in viscoelastic properties of sorghum root cell walls. Plant Cell Physiol. 2003, 44, 743–749. [Google Scholar] [CrossRef] [Green Version]
- Hattori, T.; Sonobe, K.; Inanaga, S.; An, P.; Morita, S. Effects of silicon on photosynthesis of young cucumber seedlings under osmotic tress. J. Plant Nut. 2008, 31, 1046–1058. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Element Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2019, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.J.; Chen, K.M. The regulatory role of silicon on water relations, photosynthetic gas exchange, and carboxylation activities of wheat leaves in field drought conditions. Acta Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Karmollachaab, A.; Bakhshandeh, A.; Gharineh, M.; Moradi Telavat, M.; Fathi, G. Effect of silicon application on physiological characteristics and grain yield of wheat under drought stress condition. Int. J. Agro. Plant Prod. 2013, 4, 30–37. [Google Scholar]
- Shi, Y.; Zhang, Y.; Han, W.; Feng, R.; Hu, Y.; Guo, J.; Gong, H. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.; Ahmed, S.; Ayub, M.; Naeem, M.; Sarwar, M. Impact of planting time and silicon levels on yield and yield components of sunflower (Helianthus annuus L.). Pak. J. Life Soc. Sci. 2013, 11, 60–64. [Google Scholar]
- Amin, M.; Ahmad, R.; Ali, A.; Hussain, I.; Mahmood, R.; Aslam, M.; Lee, D.J. Influence of silicon fertilization on maize performance under limited water supply. Silicon 2016, 10, 177–183. [Google Scholar] [CrossRef]
- Yahdjian, L.; Sala, O.E. Vegetation structure constrains primary production response to water availability in the Patagonian Steppe. Ecology 2006, 87, 952–962. [Google Scholar] [CrossRef]
- Hu, L.; Wang, Z.; Huang, B. Diffusion limitations and metabolic factors associated with inhibition and recovery of photosynthesis from drought stress in a C3 perennial grass species. Physiol. Plant. 2010, 139, 93–106. [Google Scholar] [CrossRef]
- Zhang, X.; Du, Y.; Wang, L.; Zhou, Q.; Huang, X.; Sun, Z. Combined effects of lanthanum (III) and acid rain on antioxidant enzyme system in soybean roots. PLoS ONE 2015, 10, e0134546. [Google Scholar] [CrossRef]
- Hossain, M.T.; Mori, R.; Soga, K.; Wakabayashi, K.; Kamisaka, S.; Fujii, S.; Yamamoto, R.; Hoson, T. Growth promotion and an increase in cell wall extensibility by silicon in rice and some other Poaceae seedlings. J. Plant Res. 2002, 115, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Akram, H.M.; Sattar, A.; Ali, A.; Nadeem, M.A. Agro-physiological performance of wheat genotypes under moisture stress conditions. J. Agric. Res. 2010, 48, 361–368. [Google Scholar]
- Sabater, B.; Rodriquez, M.I. Control of chlorophyll degradation indetached leaves of barley and oat through effect of kinetin on chlorophyllase levels. Physiol. Plant. 1978, 43, 274–276. [Google Scholar] [CrossRef]
- Gong, H.J.; Zhu, X.Y.; Chen, K.M.; Wang, S.M.; Zhang, C.L. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 2005, 169, 313–321. [Google Scholar] [CrossRef]
- Ruiz-Espinoza, F.H.; Murillo-Amador, B.; Garcia-Hernandez, J.L.; Fenech-Larios, L.; Rueda-Puente, E.O.; Troyo-Dieguez, E.; Kaya, C.; Beltran-Morales, A. Field evaluation of the relationship between chlorophyll content in basil leaves and a portable chlorophyll meter (SPAD-502) readings. J. Plant Nut. 2010, 33, 423–438. [Google Scholar] [CrossRef]
- Cornic, G. Drought stress inhibits photosynthesis by decreasing stomatal aperture-not by affecting ATP synthesis. Trends Plant Sci. 2000, 5, 187–188. [Google Scholar] [CrossRef]
- Yin, L.N.; Wang, S.W.; Li, J.Y.; Tanaka, K.; Oka, M. Application of silicon improves salt tolerance through ameliorating osmotic and ionic stresses in the seedling of Sorghum bicolor. Acta Physiol. Plant. 2013, 35, 3099–3107. [Google Scholar] [CrossRef]
- Liu, H.; Sultan, M.A.; Liu, X.L.; Zhang, J.; Yu, F.; Zhao, H.X. Physiological and comparative proteomic analysis reveals different drought responses in roots and leaves of drought tolerant wild wheat (Triticum boeoticum). PLoS ONE 2015, 10, e0121852. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K. Effect of moisture stress on physiological activities of two contrasting wheat genotypes. Ind. J. Exp. Biol. 1994, 32, 584–593. [Google Scholar]
- Rajput, V.D.; Chen, Y.; Ayup, M. Effects of high salinity on physiological and anatomical indices in the early stages of Populus euphratica growth. Russ. J. Plant Physiol. 2015, 62, 229–236. [Google Scholar] [CrossRef]
- Li, C.; Nong, Q.; Solanki, M.K.; Liang, Q.; Xie, J.; Liu, X.; Li, Y.; Wang, W.; Yang, L.; Li, Y. Differential expression profiles and pathways of genes in sugarcane leaf at elongation stage in response to drought stress. Sci. Rep. 2016, 6, 25698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.J.; Chen, K.M.; Chen, G.C.; Wang, S.M.; Zhang, C.L. Effects of silicon on growth of wheat under drought. J. Plant Nutr. 2003, 26, 1055–1063. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant Nutr. 2006, 29, 1637–1647. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Cia, M.C.; Guimaraes, A.C.R.; Medici, L.O.; Chabregas, S.M.; Azevedo, R.A. Antioxidant responses to water deficit by drought-tolerant and sensitive sugarcane varieties. Ann. Appl. Biol. 2012, 161, 313–324. [Google Scholar] [CrossRef]
- Molinari, H.B.C.; Marur, C.J.; Daros, E.; Campos, M.K.F.; Carvalho, J.F.R.P.; Bespalhok Filho, J.C.; Pereira, L.F.P.; Vieira, L.G.E. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): Osmotic adjustment, chlorophyll fluorescence and antioxidative stress. Physiol. Plant. 2007, 130, 218–225. [Google Scholar] [CrossRef]
- Wahid, A.; Close, T.J. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol. Plant. 2007, 51, 104–109. [Google Scholar] [CrossRef]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, Q.I.N.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt stressed barley (Hordium vulgare L.). Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef]
- Moussa, H.R. Influence of exogenous application of silicon on physiological response of salt-stressed maize (Zea mays L.). Int. J. Agric. Biol. 2006, 8, 293–297. [Google Scholar]
- Soylemezoglu, G.; Demir, K.; Inal, A.; Gunes, A. Effect of silicon on antioxidant and stomatal response of two grapevine (Vitis vinifera L.) rootstocks grown in boron toxic, saline and boron toxic-saline soil. Sci. Hortic. 2009, 123, 240–246. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, K.K.; Singh, P.; Song, X.P.; Malviya, M.K.; Singh, R.K.; Chen, G.L.; Solomon, S.; Li, Y.R. Mitigating climatic change for sugarcane improvement: Role of silicon in alleviating abiotic stresses. Sugar Tech. 2020. [Google Scholar] [CrossRef]
- Liang, Y. Effects of silicon on enzyme activity, and sodium, potassium and calcium concentration in barley under salt stress. Plant Soil 1999, 209, 217–224. [Google Scholar] [CrossRef]
- Ju, S.; Yin, N.; Wang, L.; Zhang, C.; Wang, Y. Effects of silicon on Oryza sativa L. seedling roots under simulated acid rain stress. PLoS ONE 2017, 12, e0173378. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.J.; Chen, K.M.; Zhao, Z.G.; Chen, G.C.; Zhou, W.J. Effects of silicon on defense of wheat against oxidative stress under drought at different developmental stages. Biol. Plant 2008, 52, 592–596. [Google Scholar] [CrossRef]
- Demidchik, V. Reactive Oxygen Species and Oxidative Stress in Plants. In Plant Stress Physiology; CAB International: Oxford, UK, 2012; pp. 24–58. [Google Scholar]
- Gunes, A.; Inal, A.; Bagci, E.G.; Pilbeam, D.J. Silicon-mediated changes of some physiological and enzymatic parameters symptomatic for oxidative stress in spinach and tomato grown in sodic-B toxic soil. Plant Soil 2007, 290, 103–114. [Google Scholar] [CrossRef]
- Peng, Q.; Zhou, Q. Antioxidant capacity of flavonoid in soybean seedlings under the joint actions of rare earth element la (III) and ultraviolet-B stress. Biol. Trace Element Res. 2009, 127, 69–80. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Silicon [mg L−1] | SML [%] | PL [cm] | LN | LAE [m2 plant−1] | LWC [%] |
|---|---|---|---|---|---|
| 0 | 100–95 | 112 ± 10af | 18 ± 3ac | 0.7504 ± 0.006aj | 92.85 ± 2.8ah |
| 100 | 117 ± 13e | 16 ± 2ej | 0.7622 ± 0.003h | 93.03 ± 1.9g | |
| 200 | 114 ± 8d | 19 ± 2aeg | 0.7501 ± 0.007al | 93.08 ± 3.6aef | |
| 300 | 122 ± 5ij | 18 ± 3be | 0.7553 ± 0.008ad | 93.01 ± 2.5ade | |
| 400 | 120 ± 9hi | 18 ± 2ad | 0.7789 ± 0.010cl | 92.98 ± 2.0ah | |
| 500 | 118 ± 11akj | 16 ± 3ai | 0.7698 ± 0.008n | 93.03 ± 3.1adi | |
| 0 | 80–75 | 83 ± 5adp | 11 ± 1d | 0.7019 ± 0.011ap | 76.03 ± 2.7fn |
| 100 | 85 ± 7al | 11 ± 2eh | 0.7141 ± 0.004o | 76.45 ± 1.7ac | |
| 200 | 89 ± 11hi | 13 ± 2fi | 0.7188 ± 0.014ak | 77.61 ± 2.0hi | |
| 300 | 95 ± 10aok | 14 ± 3ghi | 0.7250 ± 0.009ap | 79.33 ± 2.3adg | |
| 400 | 101 ± 6am | 14 ± 2d | 0.7318 ± 0.006egi | 79.87 ± 2.7ac | |
| 500 | 100 ± 8aeh | 13 ± 2f | 0.7276 ± 0.011gh | 78.92 ± 1.8e | |
| 0 | 55–50 | 48 ± 3ef | 8 ± 1ae | 0.5009 ± 0.016d | 63.01 ± 1.1ai |
| 100 | 48 ± 5ad | 8 ± 3i | 0.5091 ± 0.019ef | 64.11 ± 1.3n | |
| 200 | 50 ± 5an | 9 ± 2aj | 0.5108 ± 0.009ae | 64.06 ± 0.9f | |
| 300 | 52 ± 7f | 9 ± 1af | 0.5209 ± 0.011mn | 66.29 ± 1.0de | |
| 400 | 53 ± 4ab | 11 ± 2n | 0.5301 ± 0.013an | 69.87 ± 1.3pc | |
| 500 | 53 ± 3a | 10 ± 2ade | 0.5248 ± 0.009ci | 69.02 ± 0.8m | |
| 0 | 35–30 | 37 ± 2ce | 6 ± 1aj | 0.3678 ± 0.007fg | 40.11 ± 0.5gh |
| 100 | 37 ± 4l | 6 ± 2cj | 0.3706 ± 0.005hi | 40.83 ± 0.8op | |
| 200 | 37 ± 4ki | 6 ± 1n | 0.3817 ± 0.007ae | 41.51 ± 1.1ahi | |
| 300 | 40 ± 3aj | 7 ± 2l | 0.3873 ± 0.009afi | 41.96 ± 0.7l | |
| 400 | 42 ± 2h | 8 ± 2ap | 0.4006 ± 0.005aj | 43.07 ± 1.3aj | |
| 500 | 42 ± 3eg | 8 ± 1ace | 0.3903 ± 0.013ade | 42.63 ± 1.2ace | |
| Variation factor | |||||
| Si | ** | NS | ** | ** | |
| SML | NS | ** | NS | NS | |
| Si × SML | ** | ** | ** | ** | |
| Silicon [mg L−1 ] | SML [%] | Biomass [g] | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Lfm | Ldm | Sfm | Sdm | Rfm | Rdm | Tfm | Tdm | ||
| 0 | 100–95 | 212.55 ± 10.1ac | 69.96 ± 3.3la | 969.71 ± 9.3ad | 229.78 ± 13.1ai | 129.79 ± 4.1bc | 51.11 ± 1.8bce | 1312.05 ± 22.5gh | 350.85 ± 5.8ae |
| 100 | 210.85 ± 7.3pi | 68.11 ± 2.3j | 979.45 ± 7.1p | 230.19 ± 9.8cd | 132.59 ± 3.7ef | 51.33 ± 1.1ci | 1322.89 ± 19.1aei | 349.63 ± 6.3bgi | |
| 200 | 215.25 ± 13.3od | 70.18 ± 2.1kl | 974.04 ± 7.7ci | 230.08 ± 7.6bde | 134.11 ± 3.3cd | 52.08 ± 1.4de | 1323.4 ± 17.8hi | 352.34 ± 9.1ac | |
| 300 | 213.71 ± 6.9mn | 68.98 ± 1.8ac | 978.87 ± 6.8op | 231.19 ± 10.1ae | 137.03 ± 3.9a | 53.21 ± 0.9e | 1329.61 ± 23.3ef | 353.38 ± 5.8de | |
| 400 | 218.13 ± 10.1ca | 71.05 ± 1.3ac | 979.89 ± 5.3ai | 231.33 ± 12.5f | 138.89 ± 2.8ac | 53.95 ± 1.1ad | 1336.91 ± 17.5dei | 356.33 ± 7.4e | |
| 500 | 216.01 ± 7.9df | 69.59 ± 1.9b | 986.66 ± 8.1m | 232.06 ± 11.3ad | 141.07 ± 3.5ec | 95.24 ± 1.7g | 1343.74 ± 21.2e | 396.89 ± 10.1bc | |
| 0 | 80–75 | 191.91 ± 3.9ad | 61.85 ± 1.2fi | 765.95 ± 5.8no | 183.08 ± 7.6i | 122.37 ± 4.1aei | 43.08 ± 0.7af | 1080.23 ± 13.1aef | 288.01 ± 8.4ai |
| 100 | 192.05 ± 4.2bl | 62.03 ± 1.3ce | 766.03 ± 6.3ad | 183.23 ± 7.1bc | 123.98 ± 2.2g | 43.70 ± 0.9ig | 1082.06 ± 9.8bc | 288.96 ± 7.9ef | |
| 200 | 195.81 ± 4.1ai | 63.81 ± 1.7g | 769.07 ± 4.8l | 183.87 ± 8.5g | 124.08 ± 2.9j | 44.01 ± 1.3kn | 1088.96 ± 17.2ae | 291.69 ± 4.7ak | |
| 300 | 196.64 ± 5.6ace | 84.54 ± 2.3h | 774.25 ± 5.1ai | 184.11 ± 6.2ab | 125.81 ± 3.1l | 45.49 ± 1.1ah | 1096.7 ± 10.2k | 314.14 ± 5.8ajk | |
| 400 | 201.03 ± 4.1cn | 85.76 ± 1.6bc | 778.01 ± 4.9k | 185.23 ± 5.3h | 128.13 ± 2.2ah | 46.33 ± 1.2al | 1107.17 ± 17.8lm | 317.32 ± 6.1agh | |
| 500 | 198.63 ± 3.9op | 85.03 ± 2.1d | 775.81 ± 6.3b | 184.82 ± 7.1k | 127.44 ± 1.8acn | 46.08 ± 1.7an | 1101.88 ± 13.3ao | 315.93 ± 4.1al | |
| 0 | 55–50 | 112.55 ± 3.2ai | 39.96 ± 2.3ef | 543.30 ± 4.1c | 113.80 ± 4.3lm | 98.21 ± 1.1m | 30.70 ± 0.9jk | 754.06 ± 11.8an | 184.46 ± 4.3aeh |
| 100 | 112.85 ± 2.9ai | 40.01 ± 1.1ac | 544.08 ± 3.9ef | 113.95 ± 3.6j | 98.46 ± 1.9ajk | 31.11 ± 0.3o | 755.39 ± 13.3jl | 185.07 ± 5.1ai | |
| 200 | 115.31 ± 2.6gh | 40.64 ± 1.7fh | 545.89 ± 5.5d | 114.25 ± 2.8ao | 99.03 ± 1.6ae | 31.68 ± 0.9am | 760.23 ± 9.1ai | 186.57 ± 6.1aop | |
| 300 | 118.76 ± 3.3ah | 41.18 ± 1.3ae | 546.81 ± 2.8fh | 114.53 ± 4.1acn | 101.13 ± 1.5ao | 33.03 ± 0.5ai | 766.7 ± 13.5hik | 188.74 ± 3.8n | |
| 400 | 123.96 ± 2.1k | 41.91 ± 1.4a | 551.11 ± 3.7h | 115.08 ± 3.9p | 104.28 ± 2.1ci | 34.23 ± 0.5aeg | 779.35 ± 17.4am | 191.22 ± 5.2ace | |
| 500 | 120.72 ± 2.7j | 40.88 ± 1.2m | 548.25 ± 3.2aj | 114.68 ± 5.7ace | 102.34 ± 1.3ab | 34.08 ± 0.4p | 771.31 ± 9.6ae | 189.64 ± 4.1am | |
| 0 | 35–30 | 55.85 ± 1.8ai | 18.16 ± 0.8ap | 413.15 ± 5.8g | 83.67 ± 2.1ai | 68.05 ± 1.1ai | 15.95 ± 0.3ai | 537.05 ± 5.8eg | 117.78 ± 3.7ai |
| 100 | 56.36 ± 2.3bc | 18.21 ± 0.3lm | 413.65 ± 4.2abe | 84.03 ± 4.7bc | 68.63 ± 1.3ap | 16.16 ± 0.2ab | 538.64 ± 9.2p | 118.4 ± 3.7fgh | |
| 200 | 57.82 ± 1.9ac | 18.91 ± 0.5ai | 415.58 ± 3.3i | 83.11 ± 2.7af | 69.11 ± 1.4ac | 16.93 ± 0.2ce | 542.51 ± 9.3ce | 118.95 ± 3.1ap | |
| 300 | 58.70 ± 1.1de | 19.02 ± 0.4m | 416.ac41 ± 3.9 | 83.29 ± 1.9ac | 69.80 ± 0.9cd | 17.21 ± 0.3a | 544.91 ± 9.1ab | 119.52 ± 1.9ac | |
| 400 | 61.08 ± 1.1af | 19.93 ± 0.4ad | 419.73ai ± 3.9 | 84.02 ± 1.1ai | 71.23 ± 0.9ci | 18.16 ± 0.3ao | 552.04 ± 7.2ef | 122.11 ± 2.3ae | |
| 500 | 61.01 ± 1.9ae | 19.34 ± 0.2aeg | 417.50 ± 4.3ade | 83.91 ± 0.9adg | 71.08 ± 1.2aef | 18.11 ± 0.2de | 549.59 ± 5.6de | 121.36 ± 1.7ace | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verma, K.K.; Anas, M.; Chen, Z.; Rajput, V.D.; Malviya, M.K.; Verma, C.L.; Singh, R.K.; Singh, P.; Song, X.-P.; Li, Y.-R. Silicon Supply Improves Leaf Gas Exchange, Antioxidant Defense System and Growth in Saccharum officinarum Responsive to Water Limitation. Plants 2020, 9, 1032. https://doi.org/10.3390/plants9081032
Verma KK, Anas M, Chen Z, Rajput VD, Malviya MK, Verma CL, Singh RK, Singh P, Song X-P, Li Y-R. Silicon Supply Improves Leaf Gas Exchange, Antioxidant Defense System and Growth in Saccharum officinarum Responsive to Water Limitation. Plants. 2020; 9(8):1032. https://doi.org/10.3390/plants9081032
Chicago/Turabian StyleVerma, Krishan K., Muhammad Anas, Zhongliang Chen, Vishnu D. Rajput, Mukesh Kumar Malviya, Chhedi Lal Verma, Rajesh Kumar Singh, Pratiksha Singh, Xiu-Peng Song, and Yang-Rui Li. 2020. "Silicon Supply Improves Leaf Gas Exchange, Antioxidant Defense System and Growth in Saccharum officinarum Responsive to Water Limitation" Plants 9, no. 8: 1032. https://doi.org/10.3390/plants9081032
APA StyleVerma, K. K., Anas, M., Chen, Z., Rajput, V. D., Malviya, M. K., Verma, C. L., Singh, R. K., Singh, P., Song, X.-P., & Li, Y.-R. (2020). Silicon Supply Improves Leaf Gas Exchange, Antioxidant Defense System and Growth in Saccharum officinarum Responsive to Water Limitation. Plants, 9(8), 1032. https://doi.org/10.3390/plants9081032