
Animals Experimentally Infected with SARS-CoV-2 Generate Functional Autoantibodies against G-Protein-Coupled Receptors
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Serum Sample Preparation
2.2. Cell Preparation
2.3. Bioassay for Measurement of Functional Autoantibodies
2.4. Identification of the fAAB Binding Site on the Receptors
2.5. Statistical Analyses
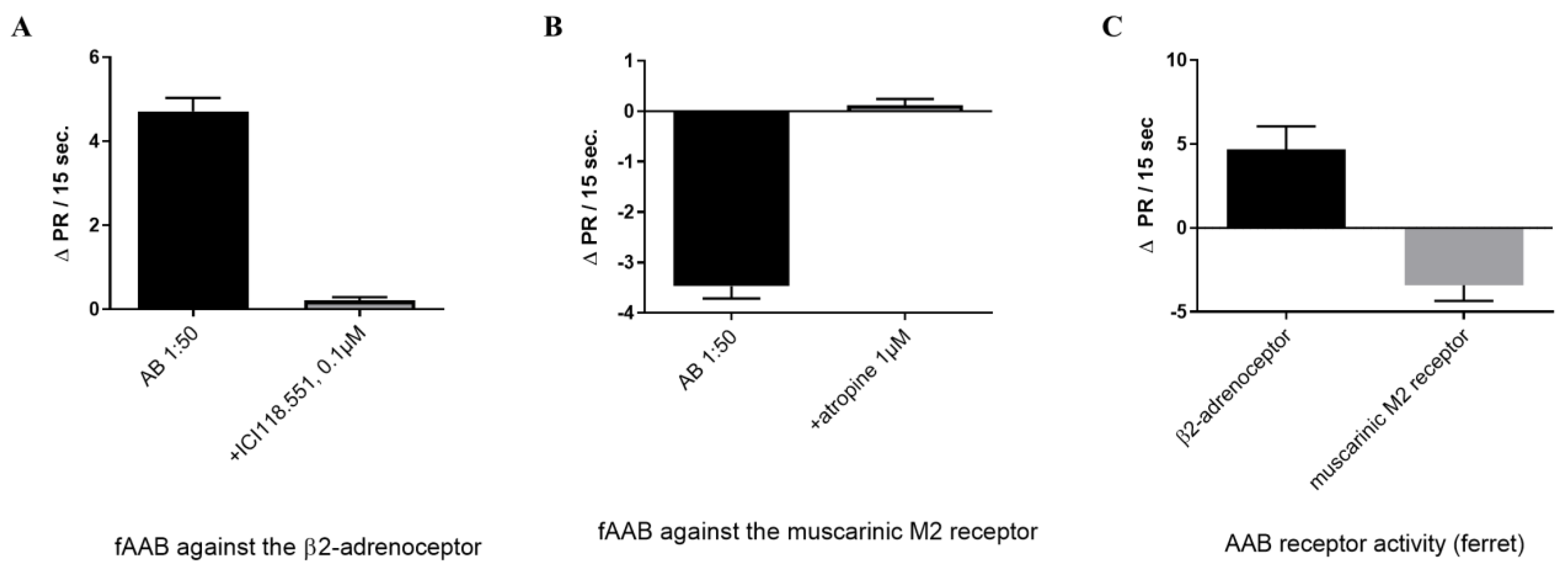
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L.; et al. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EBioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- Townsend, L.; Dyer, A.H.; Jones, K.; Dunne, J.; Mooney, A.; Gaffney, F.; O’Connor, L.; Leavy, D.; O’Brien, K.; Dowds, J.; et al. Persistent fatigue following SARS-CoV-2 infection is common and independent of severity of initial infection. PLoS ONE 2020, 15, e0240784. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; Assaf, G.S.; McCorkell, L.; Wei, H.; Low, R.J.; Re’Em, Y.; Redfield, S.; Austin, J.P.; Akrami, A. Characterizing long COVID in an international cohort: 7 months of symptoms and their impact. EClinicalMedicine 2021, 38, 101019. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Jaycox, J.R.; Liu, F.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse Functional Autoantibodies in Patients with COVID-19. Nature 2021, 595, 283–288. [Google Scholar] [CrossRef]
- Cabral-Marques, O.; Halpert, G.; Schimke, L.F.; Ostrinski, Y.; Vojdani, A.; Baiocchi, G.C.; Freire, P.P.; Filgueiras, I.S.; Zyskind, I.; Lattin, M.T.; et al. Autoantibodies targeting GPCRs and RAS-related molecules associate with COVID-19 severity. Nat. Commun. 2022, 13, 1220. [Google Scholar] [CrossRef] [PubMed]
- Wallukat, G.; Hohberger, B.; Wenzel, K.; Fürst, J.; Schulze-Rothe, S.; Wallukat, A.; Hönicke, A.-S.; Müller, J. Functional autoantibodies against G-protein coupled receptors in patients with persistent Long-COVID-19 symptoms. J. Transl. Autoimmun. 2021, 4, 100100. [Google Scholar] [CrossRef]
- Wallukat, G.; Wollenberger, A.; Morwinski, R.; Pitschner, H. Anti-beta 1-adrenoceptor autoantibodies with chronotropic activity from the serum of patients with dilated cardiomyopathy: Mapping of epitopes in the first and second extracellular loops. J. Mol. Cell. Cardiol. 1995, 27, 397–406, Correction in J. Mol. Cell. Cardiol. 1995, 27, 2529. [Google Scholar] [CrossRef] [PubMed]
- Jünemann, A.; Hohberger, B.; Rech, J.; Sheriff, A.; Fu, Q.; Schlötzer-Schrehardt, U.; Voll, R.E.; Bartel, S.; Kalbacher, H.; Hoebeke, J.; et al. Agonistic Autoantibodies to the β2-Adrenergic Receptor Involved in the Pathogenesis of Open-Angle Glaucoma. Front. Immunol. 2018, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Wallukat, G.; Schimke, I. Agonistic autoantibodies directed against G-protein-coupled receptors and their relationship to cardiovascular diseases. Semin. Immunopathol. 2014, 36, 351–363. [Google Scholar] [CrossRef]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental Infection of Cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979–2981. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, S.; Buckley, A.; Laverack, M.; Martins, M.; Palmer, M.V.; Lager, K.; Diel, D.G. Experimental Inoculation of Young Calves with SARS-CoV-2. Viruses 2021, 13, 441. [Google Scholar] [CrossRef] [PubMed]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, e218–e225. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.; Kok, A.; De Meulder, D.; Bestebroer, T.M.; Lamers, M.M.; Okba, N.M.A.; Van Vlissingen, M.F.; Rockx, B.; Haagmans, B.L.; Koopmans, M.; et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat. Commun. 2020, 11, 3496. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef]
- Pulit-Penaloza, J.A.; Belser, J.A.; Sun, X.; Pappas, C.; Brock, N.; Kieran, T.J.; Ritter, J.M.; Seixas, J.N.; Jones, J.; Thakur, P.B.; et al. Comparative Assessment of Severe Acute Respiratory Syndrome Coronavirus 2 Variants in the Ferret Model. mBio 2022, 13, e0242122. [Google Scholar] [CrossRef]
- Barut, G.T.; Halwe, N.J.; Taddeo, A.; Kelly, J.N.; Schön, J.; Ebert, N.; Ulrich, L.; Devisme, C.; Steiner, S.; Trüeb, B.S.; et al. The spike gene is a major determinant for the SARS-CoV-2 Omicron-BA.1 phenotype. Nat. Commun. 2022, 13, 5929. [Google Scholar] [CrossRef]
- Rodriguez-Perez, A.I.; Labandeira, C.M.; Pedrosa, M.A.; Valenzuela, R.; Suarez-Quintanilla, J.A.; Cortes-Ayaso, M.; Mayán-Conesa, P.; Labandeira-Garcia, J.L. Autoantibodies against ACE2 and angiotensin type-1 receptors increase severity of COVID-19. J. Autoimmun. 2021, 122, 102683. [Google Scholar] [CrossRef] [PubMed]
- Vakil, M.K.; Mansoori, Y.; Al-Awsi, G.R.L.; Hosseinipour, A.; Ahsant, S.; Ahmadi, S.; Ekrahi, M.; Montaseri, Z.; Pezeshki, B.; Mohaghegh, P.; et al. Individual genetic variability mainly of Proinflammatory cytokines, cytokine receptors, and toll-like receptors dictates pathophysiology of COVID-19 disease. J. Med. Virol. 2022, 94, 4088–4096. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Hojyo, S.; Uchida, M.; Tanaka, K.; Hasebe, R.; Tanaka, Y.; Murakami, M.; Hirano, T. How COVID-19 induces cytokine storm with high mortality. Inflamm. Regen. 2020, 40, 37. [Google Scholar] [CrossRef] [PubMed]
- Montazersaheb, S.; Khatibi, S.M.H.; Hejazi, M.S.; Tarhriz, V.; Farjami, A.; Sorbeni, F.G.; Farahzadi, R.; Ghasemnejad, T. COVID-19 infection: An overview on cytokine storm and related interventions. Virol. J. 2022, 19, 92. [Google Scholar] [CrossRef]
- Ullah, R.; Khan, J.; Basharat, N.; Huo, D.; Din, A.U.; Wang, G. Evaluation of Cardiac Biomarkers and Expression Analysis of IL-1, IL-6, IL-10, IL-17, and IL-25 among COVID-19 Patients from Pakistan. Viruses 2022, 14, 2149. [Google Scholar] [CrossRef]
- LaMarca, B.; Speed, J.; Ray, L.F.; Cockrell, K.; Wallukat, G.; Dechend, R.; Granger, J. Hypertension in response to IL-6 during pregnancy: Role of AT1-receptor activation. Int. J. Interf. Cytokine Mediat. Res. 2011, 2011, 65–70. [Google Scholar] [CrossRef]
- Csobonyeiova, M.; Smolinska, V.; Harsanyi, S.; Ivantysyn, M.; Klein, M. The Immunomodulatory Role of Cell-Free Approaches in SARS-CoV-2-Induced Cytokine Storm—A Powerful Therapeutic Tool for COVID-19 Patients. Biomedicines 2023, 11, 1736. [Google Scholar] [CrossRef] [PubMed]
- Dhillion, P.; Wallace, K.; Herse, F.; Scott, J.; Wallukat, G.; Heath, J.; Mosely, J.; Martin, J.N.; Dechend, R.; LaMarca, B.; et al. IL-17-mediated oxidative stress is an important stimulator of AT1-AA and hypertension during pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R353–R358. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, D.C.; Hogg, J.P.; Scott, J.; Wallace, K.; Herse, F.; Moseley, J.; Wallukat, G.; Dechend, R.; LaMarca, B. Administration of interleukin-17 soluble receptor C suppresses TH17 cells, oxidative stress, and hypertension in response to placental ischemia during pregnancy. Hypertension 2013, 62, 1068–1073. [Google Scholar] [CrossRef]
- Cornelius, D.C.; Lamarca, B. TH17- and IL-17- mediated autoantibodies and placental oxidative stress play a role in the pathophysiology of pre-eclampsia. Minerva Ginecol. 2014, 66, 243–249. [Google Scholar]
- Fitzgerald, S.; Deer, E.; Hogg, J.; Cornelius, D.C.; Turner, T.; Amaral, L.M.; Hoang, N.; Edwards, K.; Herrock, O.; Campbell, N.; et al. RUPP Th17s cause hypertension and mitochondrial dysfunction in the kidney and placenta during pregnancy. Pregnancy Hypertens. 2023, 32, 50–56. [Google Scholar] [CrossRef]
- Cornelius, D.C.; Amaral, L.M.; Wallace, K.; Campbell, N.; Thomas, A.J.; Scott, J.; Herse, F.; Wallukat, G.; Dechend, R.; LaMarca, B.; et al. Reduced uterine perfusion pressure T-helper 17 cells cause pathophysiology associated with preeclampsia during pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R1192–R1199. [Google Scholar] [CrossRef] [PubMed]
- Velloso, E.P.; Pimentel, R.L.; Braga, J.F.; Cabral, A.C.V.; Reis, Z.S.N.; Bader, M.; Santos, R.A.S.; Wallukat, G. Identification of a Novel Agonist-Like Autoantibody in Preeclamptic Patients. Am. J. Hypertens. 2016, 29, 405–412. [Google Scholar] [CrossRef]
- Larsen, S.B.; Wallukat, G.; Schimke, I.; Sandager, A.; Christensen, T.T.; Uldbjerg, N.; Tørring, N. Functional autoantibodies against Endothelin-1 receptor type A and Angiotensin II receptor type 1 in patients with preeclampsia. Pregnancy Hypertens. 2018, 14, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-I.; Kim, S.-G.; Kim, S.-M.; Kim, E.-H.; Park, S.-J.; Yu, K.-M.; Chang, J.-H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709.e2. [Google Scholar] [CrossRef] [PubMed]
- Hohberger, B.; Harrer, T.; Mardin, C.; Kruse, F.; Hoffmanns, J.; Rogge, L.; Heltmann, F.; Moritz, M.; Szewczykowski, C.; Schottenhamml, J.; et al. Case Report: Neutralization of Autoantibodies Targeting G-Protein-Coupled Receptors Improves Capillary Impairment and Fatigue Symptoms After COVID-19 Infection. Front. Med. 2021, 8, 754667. [Google Scholar] [CrossRef] [PubMed]
- Haberland, A.; Holtzhauer, M.; Schlichtiger, A.; Bartel, S.; Schimke, I.; Müller, J.; Dandel, M.; Luppa, P.B.; Wallukat, G. Aptamer BC 007—A broad spectrum neutralizer of pathogenic autoantibodies against G-protein-coupled receptors. Eur. J. Pharmacol. 2016, 789, 37–45. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cattle Number | Day 0 | Day 20 |
|---|---|---|
| Inoculated cattle | ||
| 776 | 0.17 | 4.69 |
| 768 | 0.59 | 5.32 |
| 766 | −0.25 | 0.00 |
| 770 | 0.17 | 0.00 |
| 771 | 0.00 | 0.50 |
| 842 | 0.00 | 0.17 |
| In−contact cattle | ||
| 774 | −0.17 | 0.17 |
| 773 | −0.33 | 0.42 |
| 777 | 0.42 | −0.09 |
| Ferret | Day 0 | Day 21 |
|---|---|---|
| 1 | 0.25 | 5.07 |
| 2 | 0.09 | 4.95 |
| 3 | 0.09 | 4.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallukat, G.; Wernike, K.; Bachamanda Somesh, D.; Mettenleiter, T.C.; Müller, J. Animals Experimentally Infected with SARS-CoV-2 Generate Functional Autoantibodies against G-Protein-Coupled Receptors. Biomedicines 2023, 11, 2668. https://doi.org/10.3390/biomedicines11102668
Wallukat G, Wernike K, Bachamanda Somesh D, Mettenleiter TC, Müller J. Animals Experimentally Infected with SARS-CoV-2 Generate Functional Autoantibodies against G-Protein-Coupled Receptors. Biomedicines. 2023; 11(10):2668. https://doi.org/10.3390/biomedicines11102668
Chicago/Turabian StyleWallukat, Gerd, Kerstin Wernike, Dipthi Bachamanda Somesh, Thomas C. Mettenleiter, and Johannes Müller. 2023. "Animals Experimentally Infected with SARS-CoV-2 Generate Functional Autoantibodies against G-Protein-Coupled Receptors" Biomedicines 11, no. 10: 2668. https://doi.org/10.3390/biomedicines11102668
APA StyleWallukat, G., Wernike, K., Bachamanda Somesh, D., Mettenleiter, T. C., & Müller, J. (2023). Animals Experimentally Infected with SARS-CoV-2 Generate Functional Autoantibodies against G-Protein-Coupled Receptors. Biomedicines, 11(10), 2668. https://doi.org/10.3390/biomedicines11102668