
Understanding Tendon Fibroblast Biology and Heterogeneity
 ,
, 
Abstract
1. Introduction

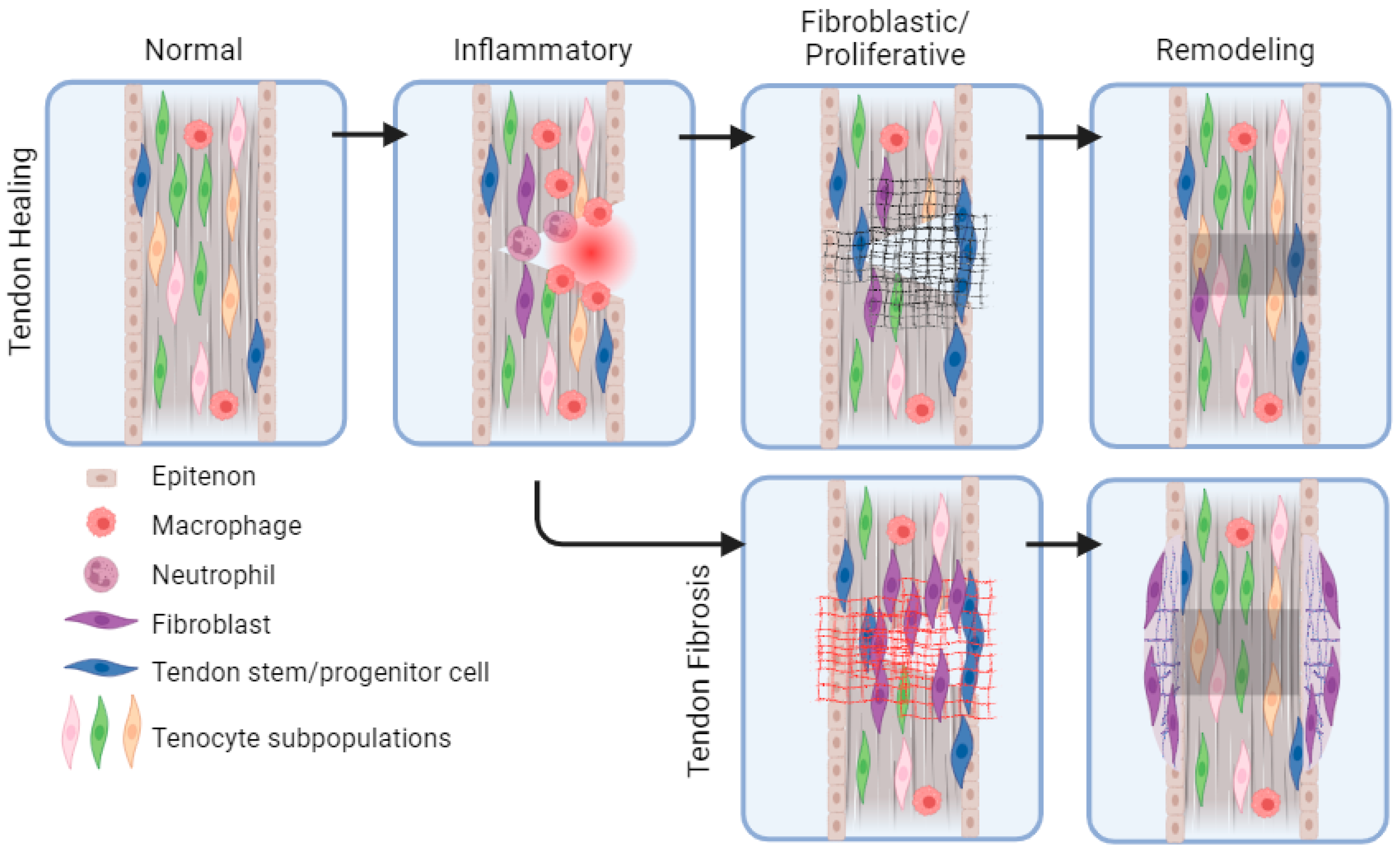{kind=link}
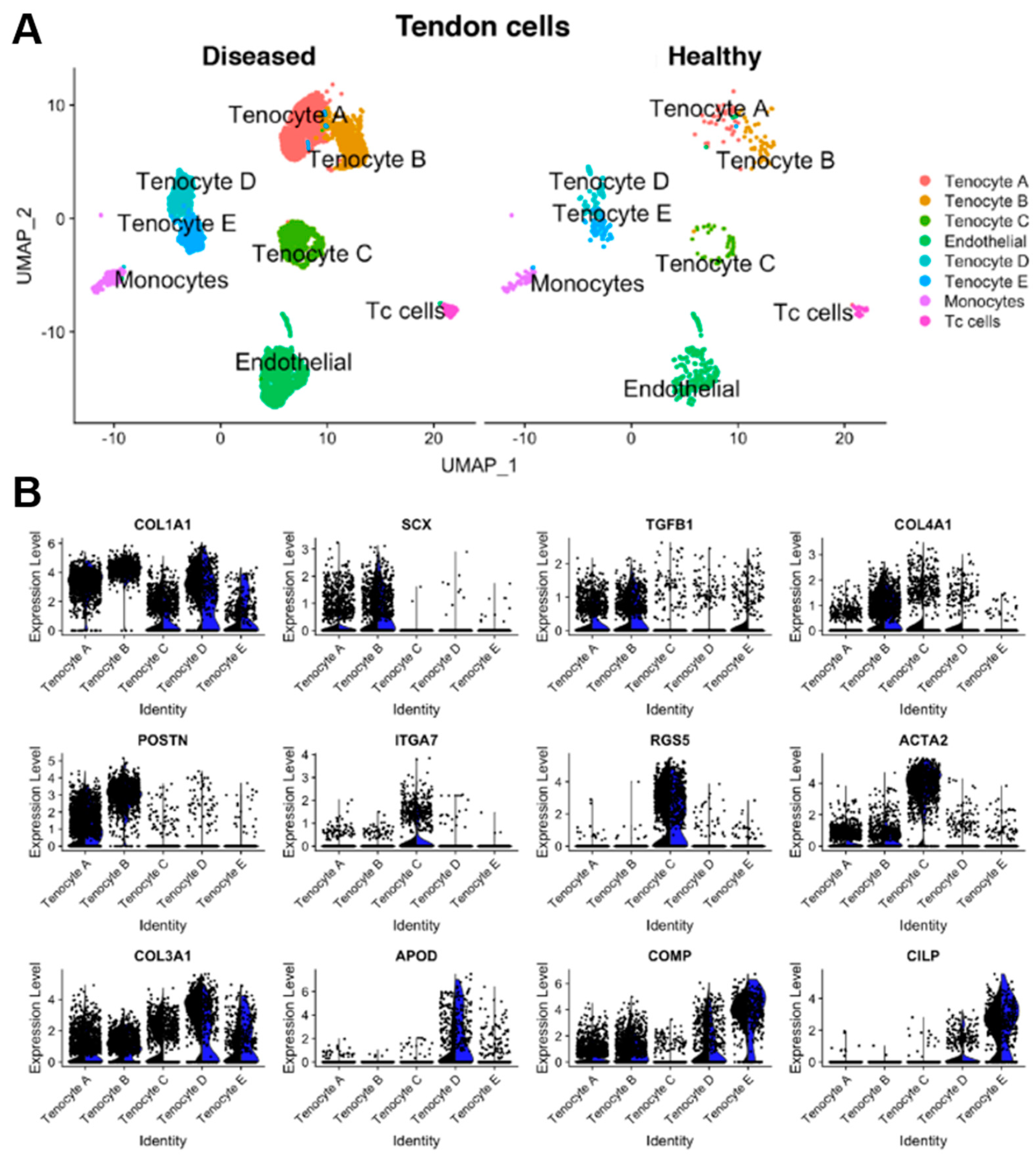{kind=link}
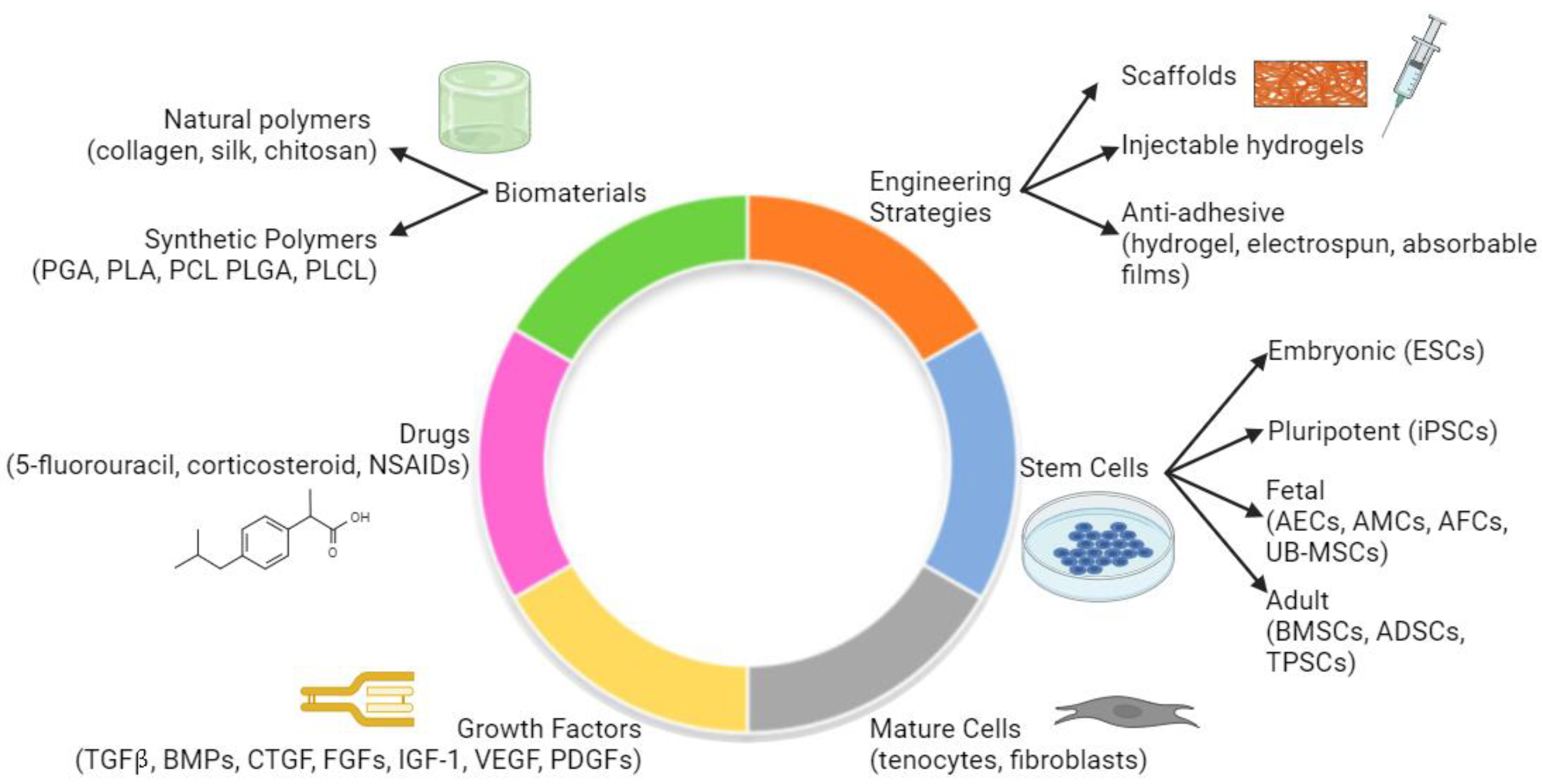{kind=link}
| Type of Tendon | Definition/Function | Key Examples |
|---|---|---|
| Synovial | Surrounded by synovial sheath and bathed in synovial fluid | Flexor tendons of hands and feet |
| Non-synovial | Surrounded by paratenon | Achilles, rotator cuff tendons |
| Energy-storing | Provide elasticity | Achilles, patellar |
| Positional | Remain stiff to transfer force to bone | Supraspinatus |
2. Overview of Tendon Healing: Cells and Signals
3. Intrinsic and Extrinsic Cell Types in Tendon Healing
4. Heterogeneity in Fibroblast Function
4.1. Fibroblast Heterogeneity in Skin
4.2. Fibroblast Heterogeneity in Tendons
5. Tendon Heterogeneity across the Body
6. Tendon Fibrosis
7. Biomaterials and Novel Solutions to Tendon Fibrosis
7.1. Overview
7.2. Anti-Adhesive Biomaterials
8. Future Directions
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Miller, K.; Hsu, J.E.; Soslowsky, L.J. Materials in Tendon and Ligament Repair. Compr. Biomater. 2011, 6, 257–279. [Google Scholar] [CrossRef]
- Screen, H.R.C.; Berk, D.E.; Kadler, K.E.; Ramirez, F.; Young, M.F. Tendon Functional Extracellular Matrix. J. Orthop. Res. 2015, 33, 793. [Google Scholar] [CrossRef]
- Harvey, T.; Flamenco, S.; Fan, C.M. A Tppp3+Pdgfra+ Tendon Stem Cell Population Contributes to Regeneration and Reveals a Shared Role for PDGF Signalling in Regeneration and Fibrosis. Nat. Cell Biol. 2019, 21, 1490–1503. [Google Scholar] [CrossRef]
- Kannus, P. Structure of the Tendon Connective Tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Thomopoulos, S.; Parks, W.C.; Rifkin, D.B.; Derwin, K.A. Mechanisms of Tendon Injury and Repair. J. Orthop. Res. 2015, 33, 832. [Google Scholar] [CrossRef]
- Scott, A.; Ashe, M.C. Common Tendinopathies in the Upper and Lower Extremities. Curr. Sports Med. Rep. 2006, 5, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Andarawis-Puri, N.; Flatow, E.L.; Soslowsky, L.J. Tendon Basic Science: Development, Repair, Regeneration, and Healing. J. Orthop. Res. 2015, 33, 780–784. [Google Scholar] [CrossRef]
- Abate, M.; Gravare Silbernagel, K.; Siljeholm, C.; Di Iorio, A.; De Amicis, D.; Salini, V.; Werner, S.; Paganelli, R. Pathogenesis of Tendinopathies: Inflammation or Degeneration? Arthritis Res. Ther. 2009, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Maffulli, N.; Khan, K.M.; Puddu, G. Overuse Tendon Conditions: Time to Change a Confusing Terminology. Arthrosc. J. Arthrosc. Relat. Surg. 1998, 14, 840–843. [Google Scholar] [CrossRef]
- Gibbon, W.W.; Cooper, J.R.; Radcliffe, G.S. Sonographic Incidence of Tendon Microtears in Athletes with Chronic Achilles Tendinosis. Br. J. Sports Med. 1999, 33, 129–130. [Google Scholar] [CrossRef]
- Bobzin, L.; Roberts, R.R.; Chen, H.J.; Crump, J.G.; Merrill, A.E. Development and Maintenance of Tendons and Ligaments. Development 2021, 148, dev186916. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.R.; Kerwin, S. Common Orthopedic Conditions and Their Physical Rehabilitation. In Canine Rehabilitation and Physical Therapy, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 543–581. [Google Scholar] [CrossRef]
- Kaya, M.; Karahan, N.; Yılmaz, B. Tendon Structure and Classification. Tendons 2019. [Google Scholar] [CrossRef]
- Liu, S.; Kang, Q.; Zhang, R.; Li, Y.; Bao, R.; Liu, S.; Kang, Q.; Zhang, R.; Li, Y.; Bao, R. Tendon Adhesion and Novel Solutions. In Tendons—Trauma, Inflammation, Degeneration, and Treatment; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Titan, A.L.; Foster, D.S.; Chang, J.; Longaker, M.T. Flexor Tendon: Development, Healing, Adhesion Formation, and Contributing Growth Factors. Plast Reconstr. Surg. 2019, 144, 639e. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.B.; Valencia, C.; Akras, D.; DiIorio, S.E.; Griffin, M.F.; Longaker, M.T.; Wan, D.C. Understanding Fibroblast Heterogeneity in Form and Function. Biomedicines 2023, 11, 2264. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, J.E.; Best, K.T.; O’Keefe, R.J.; Loiselle, A.E. Deletion of EP4 in S100a4-Lineage Cells Reduces Scar Tissue Formation during Early but Not Later Stages of Tendon Healing. Sci. Rep. 2017, 7, 8658. [Google Scholar] [CrossRef]
- Connizzo, B.K.; Yannascoli, S.M.; Soslowsky, L.J. Structure-Function Relationships of Postnatal Tendon Development: A Parallel to Healing. Matrix Biol. 2013, 32, 106. [Google Scholar] [CrossRef] [PubMed]
- Lomas, A.J.; Ryan, C.N.M.; Sorushanova, A.; Shologu, N.; Sideri, A.I.; Tsioli, V.; Fthenakis, G.C.; Tzora, A.; Skoufos, I.; Quinlan, L.R.; et al. The Past, Present and Future in Scaffold-Based Tendon Treatments. Adv. Drug Deliv. Rev. 2015, 84, 257–277. [Google Scholar] [CrossRef]
- Marsolais, D.; Côté, C.H.; Frenette, J. Neutrophils and Macrophages Accumulate Sequentially Following Achilles Tendon Injury. J. Orthop. Res. 2001, 19, 1203–1209. [Google Scholar] [CrossRef]
- Nichols, A.E.C.; Best, K.T.; Loiselle, A.E. The Cellular Basis of Fibrotic Tendon Healing: Challenges and Opportunities. Transl. Res. 2019, 209, 156–168. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, L.; Lyu, K.; Lu, J.; Long, L.; Wang, X.; Liu, T.; Li, S. A Promising Candidate in Tendon Healing Events—PDGF-BB. Biomolecules 2022, 12, 1518. [Google Scholar] [CrossRef]
- Miescher, I.; Rieber, J.; Calcagni, M.; Buschmann, J. In Vitro and In Vivo Effects of IGF-1 Delivery Strategies on Tendon Healing: A Review. Int. J. Mol. Sci. 2023, 24, 2370. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Li, W.; Ni, X.; Sui, Y.; Li, H.; Chen, X.; Lu, Y.; Jiang, M.; Wang, C. Growth Factors in the Treatment of Achilles Tendon Injury. Front. Bioeng. Biotechnol. 2023, 11, 1250533. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, S.; Wang, H.; Chen, Y.; Ding, M.Z.; Lu, J.; Jiang, L.; Lyu, K.; Huang, S.; Shi, H.; et al. The Mechanisms and Functions of TGF-Β1 in Tendon Healing. Injury 2023, 54, 111052. [Google Scholar] [CrossRef]
- Lu, J.; Jiang, L.; Chen, Y.; Lyu, K.; Zhu, B.; Li, Y.; Liu, X.; Liu, X.; Long, L.; Wang, X.; et al. The Functions and Mechanisms of Basic Fibroblast Growth Factor in Tendon Repair. Front. Physiol. 2022, 13, 852795. [Google Scholar] [CrossRef] [PubMed]
- Molloy, T.; Wang, Y.; Murrell, G.A.C. The Roles of Growth Factors in Tendon and Ligament Healing. Sports Med. 2003, 33, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Aspenberg, P.; Forslund, C. Enhanced Tendon Healing with GDF 5 and 6. Acta Orthop. Scand. 1999, 70, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Aslan, H.; Kimelman-Bleich, N.; Pelled, G.; Gazit, D. Molecular Targets for Tendon Neoformation. J. Clin. Investg. 2008, 118, 439. [Google Scholar] [CrossRef] [PubMed]
- Chisari, E.; Rehak, L.; Khan, W.S.; Maffulli, N. Tendon Healing Is Adversely Affected by Low-Grade Inflammation. J. Orthop. Surg. Res. 2021, 16, 700. [Google Scholar] [CrossRef]
- Robertson, C.M.; Chen, C.T.; Shindle, M.K.; Cordasco, F.A.; Rodeo, S.A.; Warren, R.F. Failed Healing of Rotator Cuff Repair Correlates with Altered Collagenase and Gelatinase in Supraspinatus and Subscapularis Tendons. Am. J. Sports Med. 2012, 40, 1993–2001. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, Y.; Yildirimer, L.; Xu, T.; Zhao, X. Advanced Technology-Driven Therapeutic Interventions for Prevention of Tendon Adhesion: Design, Intrinsic and Extrinsic Factor Considerations. Acta Biomater. 2021, 124, 15–32. [Google Scholar] [CrossRef]
- Wong, J.K.F.; Lui, Y.H.; Kapacee, Z.; Kadler, K.E.; Ferguson, M.W.J.; McGrouther, D.A. The Cellular Biology of Flexor Tendon Adhesion Formation: An Old Problem in a New Paradigm. Am. J. Pathol. 2009, 175, 1938–1951. [Google Scholar] [CrossRef] [PubMed]
- Muhl, L.; Genové, G.; Leptidis, S.; Liu, J.; He, L.; Mocci, G.; Sun, Y.; Gustafsson, S.; Buyandelger, B.; Chivukula, I.V.; et al. Single-Cell Analysis Uncovers Fibroblast Heterogeneity and Criteria for Fibroblast and Mural Cell Identification and Discrimination. Nat. Commun. 2020, 11, 3953. [Google Scholar] [CrossRef] [PubMed]
- Talbott, H.E.; Mascharak, S.; Griffin, M.; Wan, D.C.; Longaker, M.T. Wound Healing, Fibroblast Heterogeneity, and Fibrosis. Cell Stem. Cell 2022, 29, 1161. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jiang, S.; Liu, S.; Chen, S.; Lin, Z.Y.W.; Pan, G.; He, F.; Li, F.; Fan, C.; Cui, W. Optimization of Intrinsic and Extrinsic Tendon Healing through Controllable Water-Soluble Mitomycin-C Release from Electrospun Fibers by Mediating Adhesion-Related Gene Expression. Biomaterials 2015, 61, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Dong, N.; Zheng, Z. Small Leucine-Rich Proteoglycans in Skin Wound Healing. Front. Pharmacol. 2020, 10, 501915. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Vaughan-Thomas, A.; Clements, D.N.; Pinchbeck, G.; MacRory, L.C.; Smith, R.K.; Clegg, P.D. Gene Expression Markers of Tendon Fibroblasts in Normal and Diseased Tissue Compared to Monolayer and Three Dimensional Culture Systems. BMC Musculoskelet. Disord. 2009, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Sugg, K.B.; Markworth, J.F.; Disser, N.P.; Rizzi, A.M.; Talarek, J.R.; Sarver, D.C.; Brooks, S.V.; Mendias, C.L. Postnatal Tendon Growth and Remodeling Require Platelet-Derived Growth Factor Receptor Signaling. Am. J. Physiol. Cell Physiol. 2018, 314, C389–C403. [Google Scholar] [CrossRef] [PubMed]
- Best, K.T.; Korcari, A.; Mora, K.E.; Nichols, A.E.C.; Muscat, S.N.; Knapp, E.; Buckley, M.R.; Loiselle, A.E. Scleraxis-Lineage Cell Depletion Improves Tendon Healing and Disrupts Adult Tendon Homeostasis. Elife 2021, 10, e62203. [Google Scholar] [CrossRef]
- Manske, P.R.; Lesker, P.A. Biochemical Evidence of Flexor Tendon Participation in the Repair Process—An In Vitro Study. J. Hand Surg. Br. Eur. Vol. 1984, 9, 117–120. [Google Scholar] [CrossRef]
- Lendahl, U.; Muhl, L.; Betsholtz, C. Identification, Discrimination and Heterogeneity of Fibroblasts. Nat. Commun. 2022, 13, 3409. [Google Scholar] [CrossRef]
- Gauthier, V.; Kyriazi, M.; Nefla, M.; Pucino, V.; Raza, K.; Buckley, C.D.; Alsaleh, G. Fibroblast Heterogeneity: Keystone of Tissue Homeostasis and Pathology in Inflammation and Ageing. Front. Immunol. 2023, 14, 1137659. [Google Scholar] [CrossRef] [PubMed]
- Vorstandlechner, V.; Laggner, M.; Kalinina, P.; Haslik, W.; Radtke, C.; Shaw, L.; Lichtenberger, B.M.; Tschachler, E.; Ankersmit, H.J.; Mildner, M. Deciphering the Functional Heterogeneity of Skin Fibroblasts Using Single-Cell RNA Sequencing. FASEB J. 2020, 34, 3677–3692. [Google Scholar] [CrossRef] [PubMed]
- Janson, D.G.; Saintigny, G.; Van Adrichem, A.; Mahé, C.; El Ghalbzouri, A. Different Gene Expression Patterns in Human Papillary and Reticular Fibroblasts. J. Investig. Dermatol. 2012, 132, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Mascharak, S.; des Jardins-Park, H.E.; Davitt, M.F.; Griffin, M.; Borrelli, M.R.; Moore, A.L.; Chen, K.; Duoto, B.; Chinta, M.; Foster, D.S.; et al. Preventing Engrailed-1 Activation in Fibroblasts Yields Wound Regeneration without Scarring. Science 2021, 372, eaba2374. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Identification and Isolation of a Dermal Lineage with Intrinsic Fibrogenic Potential. Science 2015, 348, aaa2151. [Google Scholar] [CrossRef] [PubMed]
- de Micheli, A.J.; Swanson, J.B.; Disser, N.P.; Martinez, L.M.; Walker, N.R.; Oliver, D.J.; Cosgrove, B.D.; Mendias, C.L. Single-Cell Transcriptomic Analysis Identifies Extensive Heterogeneity in the Cellular Composition of Mouse Achilles Tendons. Am. J. Physiol. Cell Physiol. 2020, 319, C885–C894. [Google Scholar] [CrossRef] [PubMed]
- Kendal, A.R.; Layton, T.; Al-Mossawi, H.; Appleton, L.; Dakin, S.; Brown, R.; Loizou, C.; Rogers, M.; Sharp, R.; Carr, A. Multi-Omic Single Cell Analysis Resolves Novel Stromal Cell Populations in Healthy and Diseased Human Tendon. Sci. Rep. 2020, 10, 13939. [Google Scholar] [CrossRef] [PubMed]
- Still, C.; Chang, W.T.; Sherman, S.L.; Sochacki, K.R.; Dragoo, J.L.; Qi, L.S. Single-Cell Transcriptomic Profiling Reveals Distinct Mechanical Responses between Normal and Diseased Tendon Progenitor Cells. Cell Rep. Med. 2021, 2, 100343. [Google Scholar] [CrossRef]
- Guo, J.; Tang, H.; Huang, P.; Ye, X.; Tang, C.; Shu, Z.; Guo, J.; Kang, X.; Shi, Y.; Zhou, B.; et al. Integrative Single-Cell RNA and ATAC Sequencing Reveals That the FOXO1-PRDX2-TNF Axis Regulates Tendinopathy. Front. Immunol. 2023, 14, 1092778. [Google Scholar] [CrossRef]
- Korcari, A.; Nichols, A.E.C.; Buckley, M.R.; Loiselle, A.E. Scleraxis-Lineage Cells Are Required for Tendon Homeostasis and Their Depletion Induces an Accelerated Extracellular Matrix Aging Phenotype. Elife 2023, 12, e84194. [Google Scholar] [CrossRef]
- Masur, S.K.; Dewal, H.S.; Dinh, T.T.; Erenburg, I.; Petridou, S. Myofibroblasts Differentiate from Fibroblasts When Plated at Low Density. Proc. Natl. Acad. Sci. USA 1996, 93, 4219. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, G.G.; Rinkevich, Y.; Hu, M.S.; Montoro, D.T.; Lo, D.D.; McArdle, A.; Maan, Z.N.; Morrison, S.D.; Duscher, D.; Whittam, A.J.; et al. Live Fibroblast Harvest Reveals Surface Marker Shift In Vitro. Tissue Eng. Part C Methods 2015, 21, 314. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, M.; Kaiser, E.; Milz, S. Structure-Function Relationships in Tendons: A Review. J. Anat. 2008, 212, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Michener, L.A.; Kulig, K. Not All Tendons Are Created Equal: Implications for Differing Treatment Approaches. J. Orthop. Sports Phys. Ther. 2015, 45, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Scheffler, S.U.; Unterhauser, F.N.; Weiler, A. Graft Remodeling and Ligamentization after Cruciate Ligament Reconstruction. Knee Surg. Sports Traumatol. Arthrosc. 2008, 16, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Claes, S.; Verdonk, P.; Forsyth, R.; Bellemans, J. The “Ligamentization” Process in Anterior Cruciate Ligament Reconstruction. Am. J. Sports Med. 2011, 39, 2476–2483. [Google Scholar] [CrossRef]
- Thorpe, C.T.; Birch, H.L.; Clegg, P.D.; Screen, H.R.C. Tendon Physiology and Mechanical Behavior: Structure–Function Relationships. In Tendon Regeneration: Understanding Tissue Physiology and Development to Engineer Functional Substitutes; Elsevier: Amsterdam, The Netherlands, 2015; pp. 3–39. [Google Scholar] [CrossRef]
- Thorpe, C.T.; Godinho, M.S.C.; Riley, G.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R.C. The Interfascicular Matrix Enables Fascicle Sliding and Recovery in Tendon, and Behaves More Elastically in Energy Storing Tendons. J. Mech. Behav. Biomed. Mater. 2015, 52, 85–94. [Google Scholar] [CrossRef]
- Godinho, M.S.C.; Thorpe, C.T.; Greenwald, S.E.; Screen, H.R.C. Elastin Is Localised to the Interfascicular Matrix of Energy Storing Tendons and Becomes Increasingly Disorganised with Ageing. Sci. Rep. 2017, 7, 9713. [Google Scholar] [CrossRef]
- Thorpe, C.T.; Udeze, C.P.; Birch, H.L.; Clegg, P.D.; Screen, H.R.C. Specialization of Tendon Mechanical Properties Results from Interfascicular Differences. J. R. Soc. Interface 2012, 9, 3108–3117. [Google Scholar] [CrossRef]
- Ishii, Y.; Deie, M.; Adachi, N.; Hazeki, O.; Okamura, N.; Sharman, P.; Yasunaga, Y.; Ochi, M. Comparison of the Protein Profiles between the Achilles and Patella Tendon in Rats. Mater. Sci. Eng. C 2008, 28, 1319–1321. [Google Scholar] [CrossRef]
- Steffen, D.; Avey, A.; Mienaltowski, M.J.; Baar, K. The Rat Achilles and Patellar Tendons Have Similar Increases in Mechanical Properties but Become Transcriptionally Divergent during Postnatal Development. J. Physiol. 2023, 601, 3869–3884. [Google Scholar] [CrossRef] [PubMed]
- Disser, N.P.; Ghahramani, G.C.; Swanson, J.B.; Wada, S.; Chao, M.L.; Rodeo, S.A.; Oliver, D.J.; Mendias, C.L. Widespread Diversity in the Transcriptomes of Functionally Divergent Limb Tendons. J. Physiol. 2020, 598, 1537. [Google Scholar] [CrossRef] [PubMed]
- Thomopoulos, S.; Genin, G.M.; Galatz, L.M. The Development and Morphogenesis of the Tendon-to-Bone Insertion What Development Can Teach Us about Healing. J. Musculoskelet. Neuronal. Interact. 2010, 10, 35. [Google Scholar] [PubMed]
- Carmont, M.R.; Knutsson, S.B.; Brorsson, A.; Karlsson, J.; Nilsson-Helander, K. The Release of Adhesions Improves Outcome Following Minimally Invasive Repair of Achilles Tendon Rupture. Knee Surg. Sports Traumatol. Arthrosc. 2022, 30, 1109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, J.H.C. Mechanobiological Response of Tendon Stem Cells: Implications of Tendon Homeostasis and Pathogenesis of Tendinopathy. J. Orthop. Res. 2010, 28, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.Y.; Gelberman, R.H.; Cobb, N.G.; Amiel, D.; Lothringer, K.; Akeson, W.H. The Importance of Controlled Passive Mobilization on Flexor Tendon Healing: A Biomechanical Study. Acta Orthop. Scand. 1981, 52, 615–622. [Google Scholar] [CrossRef]
- Citeroni, M.R.; Ciardulli, M.C.; Russo, V.; Porta, G.D.; Mauro, A.; Khatib, M.E.; Di Mattia, M.; Galesso, D.; Barbera, C.; Forsyth, N.R.; et al. In Vitro Innovation of Tendon Tissue Engineering Strategies. Int. J. Mol. Sci. 2020, 21, 6726. [Google Scholar] [CrossRef]
- Müller, S.A.; Todorov, A.; Heisterbach, P.E.; Martin, I.; Majewski, M. Tendon Healing: An Overview of Physiology, Biology, and Pathology of Tendon Healing and Systematic Review of State of the Art in Tendon Bioengineering. Knee Surg. Sports Traumatol. Arthrosc. 2015, 23, 2097–2105. [Google Scholar] [CrossRef] [PubMed]
- Leong, N.L.; Kator, J.L.; Clemens, T.L.; James, A.; Enamoto-Iwamoto, M.; Jiang, J. Tendon and Ligament Healing and Current Approaches to Tendon and Ligament Regeneration. J. Orthop. Res. 2020, 38, 7–12. [Google Scholar] [CrossRef]
- Manning, C.N.; Schwartz, A.G.; Liu, W.; Xie, J.; Havlioglu, N.; Sakiyama-Elbert, S.E.; Silva, M.J.; Xia, Y.; Gelberman, R.H.; Thomopoulos, S. Controlled Delivery of Mesenchymal Stem Cells and Growth Factors Using a Nanofiber Scaffold for Tendon Repair. Acta Biomater. 2013, 9, 6905. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Y.; Ha, P.; Chen, Y.; Li, C.; Yen, E.; Bai, Y.; Chen, R.; Wu, B.M.; Da Lio, A.; et al. A Novel Injectable Fibromodulin-releasing Granular Hydrogel for Tendon Healing and Functional Recovery. Bioeng. Transl. Med. 2023, 8, 10355. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhu, Y.; Xu, J.; Tong, W.; Hu, S.; Chen, Y.F.; Deng, S.; Yao, H.; Li, J.; Lee, C.W.; et al. Injectable Bioactive Glass/Sodium Alginate Hydrogel with Immunomodulatory and Angiogenic Properties for Enhanced Tendon Healing. Bioeng. Transl. Med. 2022, 8, 10345. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Liang, Y.; Zhang, J.; Bai, L.; Xu, M.; Han, Q.; Han, X.; Xiu, J.; Li, M.; Zhou, X.; et al. Synergistic Enhancement of Tendon-to-Bone Healing via Anti-Inflammatory and pro-Differentiation Effects Caused by Sustained Release of Mg2+/Curcumin from Injectable Self-Healing Hydrogels. Theranostics 2021, 11, 5911. [Google Scholar] [CrossRef]
- Liu, C.; Yu, K.; Bai, J.; Tian, D.; Liu, G. Experimental Study of Tendon Sheath Repair via Decellularized Amnion to Prevent Tendon Adhesion. PLoS ONE 2018, 13, e0205811. [Google Scholar] [CrossRef]
- Zhou, H.; Lu, H. Advances in the Development of Anti-Adhesive Biomaterials for Tendon Repair Treatment. Tissue Eng. Regen. Med. 2021, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Menderes, A.; Mola, F.; Tayfur, V.; Vayvada, H.; Barutçu, A. Prevention of Peritendinous Adhesions Following Flexor Tendon Injury with Seprafilm. Ann. Plast Surg. 2004, 53, 560–564. [Google Scholar] [CrossRef]
- Yilmaz, E.; Avci, M.; Bulut, M.; Kelestimur, H.; Karakurt, L.; Ozercan, I. The Effect of Seprafilm on Adhesion Formation and Tendon Healing after Flexor Tendon Repair in Chicken. Orthopedics 2010, 33, 164–170. [Google Scholar] [CrossRef]
- Karaaltin, M.V.; Ozalp, B.; Dadaci, M.; Kayikcioglu, A.; Kecik, A.; Oner, F. The Effects of 5-Fluorouracil on Flexor Tendon Healing by Using a Biodegradable Gelatin, Slow Releasing System: Experimental Study in a Hen Model. J. Hand Surg. Eur. Vol. 2013, 38, 651–657. [Google Scholar] [CrossRef]
- Freedman, B.R.; Kuttler, A.; Beckmann, N.; Nam, S.; Kent, D.; Schuleit, M.; Ramazani, F.; Accart, N.; Rock, A.; Li, J.; et al. Enhanced Tendon Healing by a Tough Hydrogel with an Adhesive Side and High Drug-Loading Capacity. Nat. Biomed. Eng. 2022, 6, 1167. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A. Electrospun Nanofibers as Carriers of Microorganisms, Stem Cells, Proteins, and Nucleic Acids in Therapeutic and Other Applications. Front. Bioeng. Biotechnol. 2020, 8, 511400. [Google Scholar] [CrossRef]
- Turner, J.B.; Corazzini, R.L.; Butler, T.J.; Garlick, D.S.; Rinker, B.D. Evaluating Adhesion Reduction Efficacy of Type I/III Collagen Membrane and Collagen-GAG Resorbable Matrix in Primary Flexor Tendon Repair in a Chicken Model. Hand 2015, 10, 482. [Google Scholar] [CrossRef] [PubMed]
- Capella-Monsonís, H.; Kelly, J.; Kearns, S.; Zeugolis, D.I. Decellularised Porcine Peritoneum as a Tendon Protector Sheet. Biomed. Mater. 2019, 14, 044102. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.H.; Galloway, J.L. Current and Emerging Technologies for Defining and Validating Tendon Cell Fate. J. Orthop. Res. 2023, 41, 2082–2092. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ning, B.; Shi, T. Single-Cell RNA-Seq Technologies and Related Computational Data Analysis. Front. Genet. 2019, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, J.E.; Best, K.T.; Muscat, S.N.; Pritchett, E.M.; Nichols, A.E.C.; Wu, C.L.; Loiselle, A.E. Defining the Spatial-Molecular Map of Fibrotic Tendon Healing and the Drivers of Scleraxis-Lineage Cell Fate and Function. Cell Rep. 2022, 41, 111706. [Google Scholar] [CrossRef]
- Goltsev, Y.; Nolan, G. CODEX Multiplexed Tissue Imaging. Nat. Rev. Immunol. 2023, 23, 613. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.; Li, P.; Gao, C.; Fu, L.; Liao, Z.; Tian, G.; Yin, H.; Li, M.; Sui, X.; Yuan, Z.; et al. Recent Advances in Tendon Tissue Engineering Strategy. Front. Bioeng. Biotechnol. 2023, 11, 1115312. [Google Scholar] [CrossRef]
- Connizzo, B.K.; Grodzinsky, A.J. Release of Pro-Inflammatory Cytokines from Muscle and Bone Causes Tenocyte Death in a Novel Rotator Cuff In Vitro Explant Culture Model. Connect. Tissue Res. 2018, 59, 423–436. [Google Scholar] [CrossRef]



| Key Players in Tendon Healing | |||
|---|---|---|---|
| Cell Type | Primary Phase Present | Primary Function | Reference |
| Erythrocytes | Early inflammatory | Can be present from broken vessels | Connizzo et al. [18] |
| Platelets | Early inflammatory | Clot blood vessels broke in injury, secrete PDGF | Chen et al. [22] |
| Neutrophils | First 24 h | Phagocytose necrotic tissue | Marsolais et al. [20] |
| Macrophages | Inflammatory phase, typically after neutrophils | Phagocytosis; release of growth factors to stimulate fibroblasts, increase ECM synthesis, and decrease ECM degradation | Marsolais et al. [20] |
| Fibroblasts | Inflammatory | Migrate into site of injury | Lomas et al. [19]; Parker et al. [16] |
| Proliferative | Deposit collagen III | ||
| Remodeling | Deposit collagen I | ||
| Tenocytes | Present in tendon for all phases | Contribute to intrinsic tendon healing and collagen formation | Nichols et al. [21] |
| Tendon Stem and Progenitor Cells | Present in sheath for all phases | Differentiate to form mature tenocytes for tissue regeneration | Harvey et al. [3] |
| Growth Factor or Enzyme | Primary Phase Present | Primary Function | Reference |
| PDGF | Inflammatory and remodeling | Induces synthesis of IGF-I and IGF receptors; stimulate collagen, protein, and DNA production; stimulate macrophage migration | Chen et al. [22] |
| IGF-I | Inflammatory and proliferative | Stimulate ECM formation; stimulates migration and proliferation of fibroblasts and tendon stem cells | Miescher et al. [23] |
| VEGF | Proliferative and remodeling | Promotes angiogenesis | Lin et al. [24] |
| TGF-β | All phases | Many functions including collagen production, cell viability, promoting Scx expression | Li et al. [25] |
| bFGF | All phases | Angiogenesis, cellular migration and proliferation; Promotes expression of other growth factors | Lu et al. [26] |
| MMPs | All phases | Degradation and turnover of collagen, glycoproteins, proteoglycans | Andarawis-Puri et al. [7] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DiIorio, S.E.; Young, B.; Parker, J.B.; Griffin, M.F.; Longaker, M.T. Understanding Tendon Fibroblast Biology and Heterogeneity. Biomedicines 2024, 12, 859. https://doi.org/10.3390/biomedicines12040859
DiIorio SE, Young B, Parker JB, Griffin MF, Longaker MT. Understanding Tendon Fibroblast Biology and Heterogeneity. Biomedicines. 2024; 12(4):859. https://doi.org/10.3390/biomedicines12040859
Chicago/Turabian StyleDiIorio, Sarah E., Bill Young, Jennifer B. Parker, Michelle F. Griffin, and Michael T. Longaker. 2024. "Understanding Tendon Fibroblast Biology and Heterogeneity" Biomedicines 12, no. 4: 859. https://doi.org/10.3390/biomedicines12040859
APA StyleDiIorio, S. E., Young, B., Parker, J. B., Griffin, M. F., & Longaker, M. T. (2024). Understanding Tendon Fibroblast Biology and Heterogeneity. Biomedicines, 12(4), 859. https://doi.org/10.3390/biomedicines12040859