
Platelet-Rich Fibrin Increases CXCL8 Expression in Gingival Fibroblasts

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Gingival Fibroblasts
2.2. PRF Lysates and PRF Serum
2.3. Treatments of Cells
2.4. Reverse Transcription-Quantitative Real-Time PCR and Immunoassay
2.5. Immunofluorescence Analysis
2.6. Western Blot
2.7. Statistical Analysis
3. Results
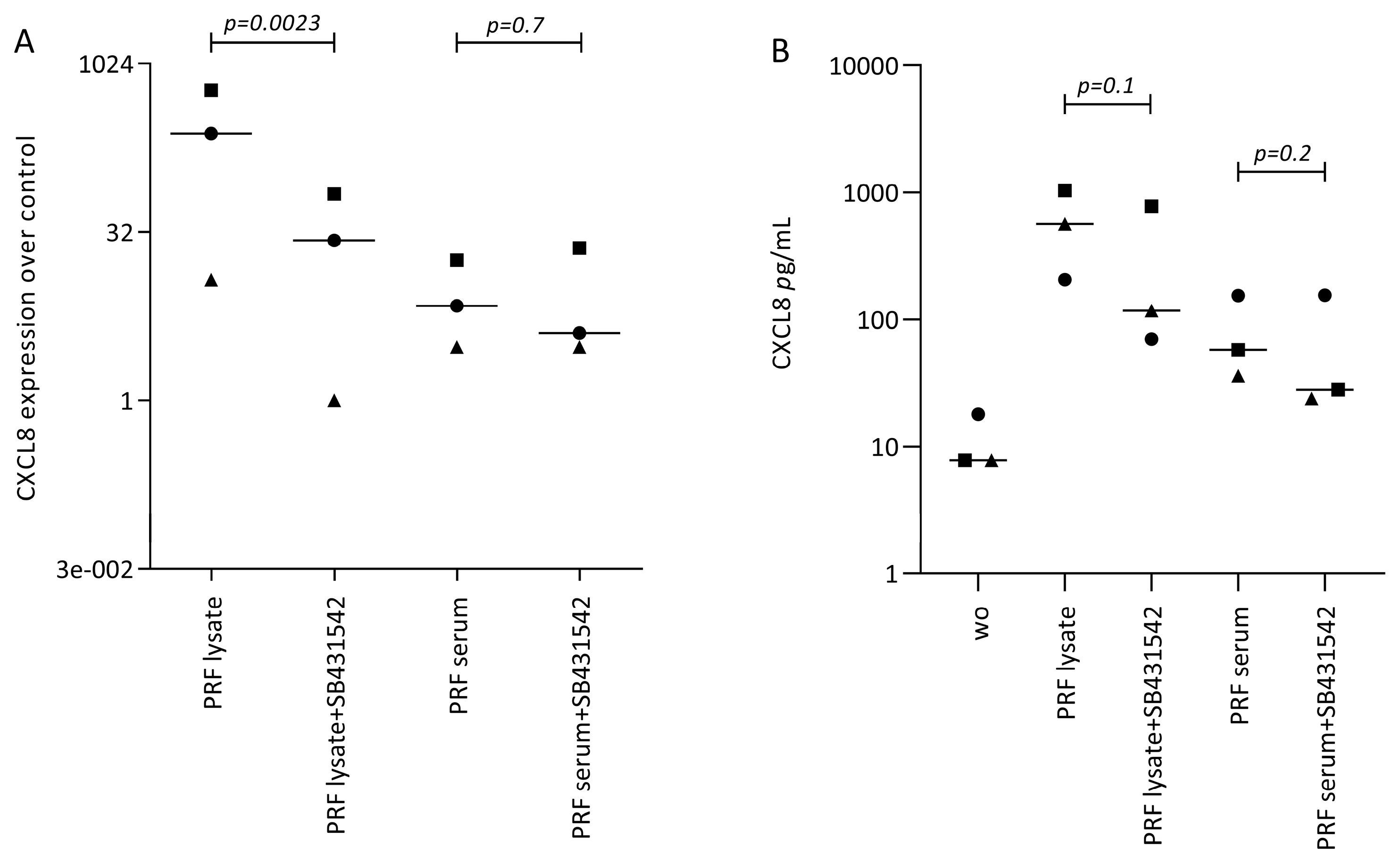
3.1. PRF Lysates and PRF Serum Increase CXCL8 in Gingival Fibroblasts
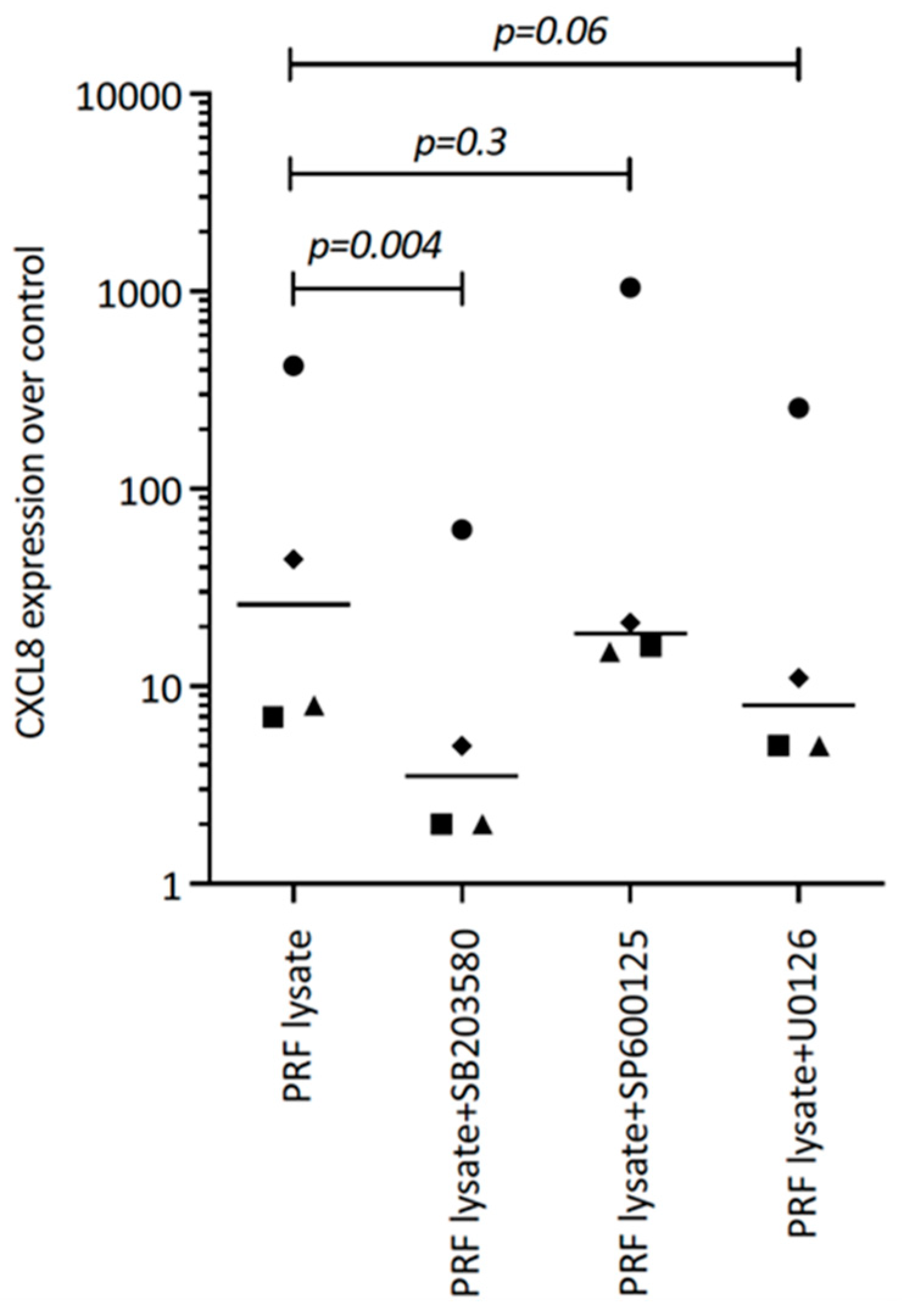
3.2. PRF Lysate Requires TGF-β, p38 and PI3K Signaling to Drive CXCL8 Expression
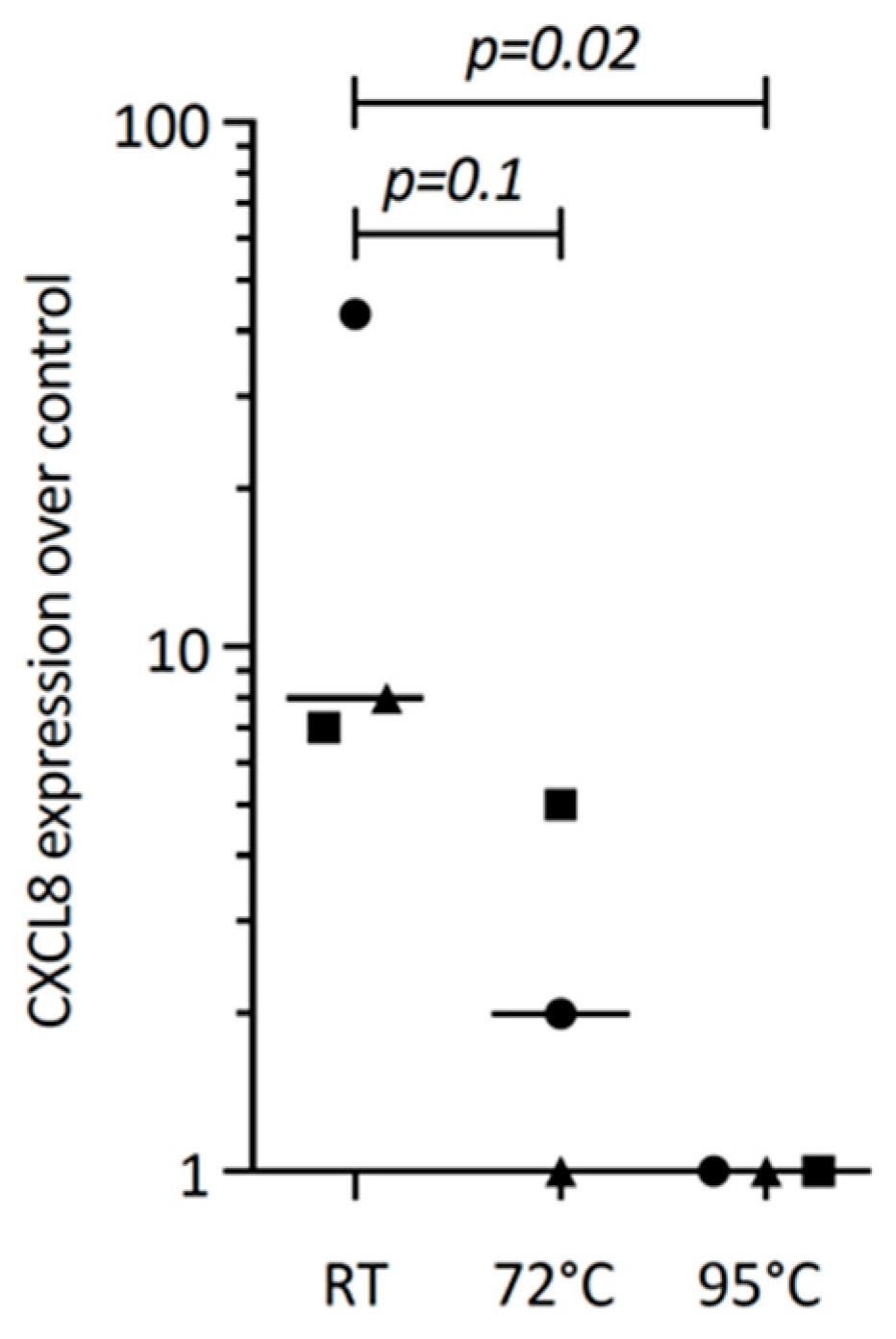
3.3. PRF Lysates Contain Heat-Sensitive Molecules Enhancing CXCL8 Expression
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miron, R.J.; Fujioka-Kobayashi, M.; Sculean, A.; Zhang, Y. Optimization of platelet-rich fibrin. Periodontol. 2000 2023, 94, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.B.; Van Dessel, J.; Temmerman, A.; Jacobs, R.; Quirynen, M. Effect of different platelet-rich fibrin matrices for ridge preservation in multiple tooth extractions: A split-mouth randomized controlled clinical trial. J. Clin. Periodontol. 2021, 48, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Chauca-Bajana, L.; Caponio, V.C.A.; Cueva, K.A.S.; Velasquez-Ron, B.; Padin-Iruegas, M.E.; Almeida, L.L.; Lorenzo-Pouso, A.I.; Suarez-Penaranda, J.M.; Perez-Sayans, M. Regeneration of periodontal intrabony defects using platelet-rich fibrin (PRF): A systematic review and network meta-analysis. Odontology 2024. [Google Scholar] [CrossRef]
- Tavelli, L.; Chen, C.J.; Barootchi, S.; Kim, D.M. Efficacy of biologics for the treatment of periodontal infrabony defects: An American Academy of Periodontology best evidence systematic review and network meta-analysis. J. Periodontol. 2022, 93, 1803–1826. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, H.; Zhang, Z.; Yan, Z.; Lv, H.; Zhang, Y.; Wu, B. Platelet-rich fibrin exudate promotes the proliferation and osteogenic differentiation of human periodontal ligament cells in vitro. Mol. Med. Rep. 2018, 18, 4477–4485. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.B.; Cortellini, S.; Temmerman, A.; Li, X.; Pinto, N.; Teughels, W.; Quirynen, M. Characterization of the Leukocyte- and Platelet-Rich Fibrin Block: Release of Growth Factors, Cellular Content, and Structure. Int. J. Oral. Maxillofac. Implant. 2019, 34, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Cambier, S.; Gouwy, M.; Proost, P. The chemokines CXCL8 and CXCL12: Molecular and functional properties, role in disease and efforts towards pharmacological intervention. Cell. Mol. Immunol. 2023, 20, 217–251. [Google Scholar] [CrossRef]
- Frasheri, I.; Heym, R.; Ern, C.; Summer, B.; Hennessen, T.G.; Hogg, C.; Reichl, F.X.; Folwaczny, M. Salivary and gingival CXCL8 correlation with periodontal status, periodontal pathogens, and smoking. Oral Dis. 2022, 28, 2267–2276. [Google Scholar] [CrossRef]
- Finoti, L.S.; Nepomuceno, R.; Pigossi, S.C.; Corbi, S.C.; Secolin, R.; Scarel-Caminaga, R.M. Association between interleukin-8 levels and chronic periodontal disease: A PRISMA-compliant systematic review and meta-analysis. Medicine 2017, 96, e6932. [Google Scholar] [CrossRef]
- Williams, D.W.; Greenwell-Wild, T.; Brenchley, L.; Dutzan, N.; Overmiller, A.; Sawaya, A.P.; Webb, S.; Martin, D.; Genomics, N.N.; Computational Biology, C.; et al. Human oral mucosa cell atlas reveals a stromal-neutrophil axis regulating tissue immunity. Cell 2021, 184, 4090–4104.e15. [Google Scholar] [CrossRef]
- Mo, J.J.; Lai, Y.R.; Huang, Q.R.; Li, Y.R.; Zhang, Y.J.; Chen, R.Y.; Qian, S.J. Single-cell sequencing identifies inflammation-promoting fibroblast-neutrophil interaction in peri-implantitis. J. Clin. Periodontol. 2024, 51, 196–208. [Google Scholar] [CrossRef]
- Andia, I.; Rubio-Azpeitia, E. Angiogenic and innate immune responses triggered by PRP in tendon cells are not modified by hyperuricemia. Muscles Ligaments Tendons J. 2014, 4, 292–297. [Google Scholar] [CrossRef]
- Assirelli, E.; Filardo, G.; Mariani, E.; Kon, E.; Roffi, A.; Vaccaro, F.; Marcacci, M.; Facchini, A.; Pulsatelli, L. Effect of two different preparations of platelet-rich plasma on synoviocytes. Knee Surg. Sports Traumatol. Arthrosc. 2015, 23, 2690–2703. [Google Scholar] [CrossRef] [PubMed]
- Kasnak, G.; Fteita, D.; Jaatinen, O.; Kononen, E.; Tunali, M.; Gursoy, M.; Gursoy, U.K. Regulatory effects of PRF and titanium surfaces on cellular adhesion, spread, and cytokine expressions of gingival keratinocytes. Histochem. Cell Biol. 2019, 152, 63–73. [Google Scholar] [CrossRef]
- Di Summa, F.; Kargarpour, Z.; Nasirzade, J.; Stahli, A.; Mitulovic, G.; Panic-Jankovic, T.; Koller, V.; Kaltenbach, C.; Muller, H.; Panahipour, L.; et al. TGFbeta activity released from platelet-rich fibrin adsorbs to titanium surface and collagen membranes. Sci. Rep. 2020, 10, 10203. [Google Scholar] [CrossRef] [PubMed]
- Fujioka-Kobayashi, M.; Miron, R.J.; Hernandez, M.; Kandalam, U.; Zhang, Y.; Choukroun, J. Optimized Platelet-Rich Fibrin With the Low-Speed Concept: Growth Factor Release, Biocompatibility, and Cellular Response. J. Periodontol. 2017, 88, 112–121. [Google Scholar] [CrossRef]
- Qi, W.; Chen, X.; Polhill, T.S.; Sumual, S.; Twigg, S.; Gilbert, R.E.; Pollock, C.A. TGF-beta1 induces IL-8 and MCP-1 through a connective tissue growth factor-independent pathway. Am. J. Physiol. Renal Physiol. 2006, 290, F703–F709. [Google Scholar] [CrossRef]
- Lu, S.; Dong, Z. Characterization of TGF-beta-regulated interleukin-8 expression in human prostate cancer cells. Prostate 2006, 66, 996–1004. [Google Scholar] [CrossRef]
- Fong, Y.C.; Maa, M.C.; Tsai, F.J.; Chen, W.C.; Lin, J.G.; Jeng, L.B.; Yang, R.S.; Fu, W.M.; Tang, C.H. Osteoblast-derived TGF-beta1 stimulates IL-8 release through AP-1 and NF-kappaB in human cancer cells. J. Bone Miner. Res. 2008, 23, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Krick, S.; Baumlin, N.; Aller, S.P.; Aguiar, C.; Grabner, A.; Sailland, J.; Mendes, E.; Schmid, A.; Qi, L.; David, N.V.; et al. Klotho Inhibits Interleukin-8 Secretion from Cystic Fibrosis Airway Epithelia. Sci. Rep. 2017, 7, 14388. [Google Scholar] [CrossRef]
- Fu, Y.; Yip, A.; Seah, P.G.; Blasco, F.; Shi, P.Y.; Herve, M. Modulation of inflammation and pathology during dengue virus infection by p38 MAPK inhibitor SB203580. Antiviral Res. 2014, 110, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Sunil, Y.; Ramadori, G.; Raddatzc, D. Influence of NFkappaB inhibitors on IL-1beta-induced chemokine CXCL8 and -10 expression levels in intestinal epithelial cell lines: Glucocorticoid ineffectiveness and paradoxical effect of PDTC. Int. J. Color. Dis. 2010, 25, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Higham, A.; Singh, D. Dexamethasone and p38 MAPK inhibition of cytokine production from human lung fibroblasts. Fundam. Clin. Pharmacol. 2021, 35, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A. Nf-kappa B, chemokine gene transcription and tumour growth. Nat. Rev. Immunol. 2002, 2, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Mao, P.; Wu, S.; Li, J.; Fu, W.; He, W.; Liu, X.; Slutsky, A.S.; Zhang, H.; Li, Y. Human alveolar epithelial type II cells in primary culture. Physiol. Rep. 2015, 3, e12288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.E. Non-Smad pathways in TGF-beta signaling. Cell Res. 2009, 19, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Schafer, S.; Viswanathan, S.; Widjaja, A.A.; Lim, W.W.; Moreno-Moral, A.; DeLaughter, D.M.; Ng, B.; Patone, G.; Chow, K.; Khin, E.; et al. IL-11 is a crucial determinant of cardiovascular fibrosis. Nature 2017, 552, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Sterczala, B.; Chwilkowska, A.; Szwedowicz, U.; Kobielarz, M.; Chwilkowski, B.; Dominiak, M. Impact of APRF+ in Combination with Autogenous Fibroblasts on Release Growth Factors, Collagen, and Proliferation and Migration of Gingival Fibroblasts: An In Vitro Study. Materials 2022, 15, 796. [Google Scholar] [CrossRef] [PubMed]
- Bucur, M.; Constantin, C.; Neagu, M.; Zurac, S.; Dinca, O.; Vladan, C.; Cioplea, M.; Popp, C.; Nichita, L.; Ionescu, E. Alveolar blood clots and platelet-rich fibrin induce in vitro fibroblast proliferation and migration. Exp. Ther. Med. 2019, 17, 982–989. [Google Scholar] [CrossRef]
- Dong, B.; Zhu, J.; Chen, X.; Jiang, H.; Deng, Y.; Xu, L.; Wang, Y.; Li, S. The Emerging Role of Interleukin-(IL)-11/IL-11R in Bone Metabolism and Homeostasis: From Cytokine to Osteokine. Aging Dis. 2023, 14, 2113–2126. [Google Scholar] [CrossRef]
- Cook, S.A. Understanding interleukin 11 as a disease gene and therapeutic target. Biochem. J. 2023, 480, 1987–2008. [Google Scholar] [CrossRef] [PubMed]
- Kargarpour, Z.; Nasirzade, J.; Panahipour, L.; Miron, R.J.; Gruber, R. Platelet-Rich Fibrin Decreases the Inflammatory Response of Mesenchymal Cells. Int. J. Mol. Sci. 2021, 22, 1333. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Zhang, X.; Zhao, Q.; Chai, J.; Zhang, Y. Liquid platelet-rich fibrin promotes the regenerative potential of human periodontal ligament cells. Oral Dis. 2020, 26, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X. Lipopolysaccharide-regulated production of bone sialoprotein and interleukin-8 in human periodontal ligament fibroblasts: The role of toll-like receptors 2 and 4 and the MAPK pathway. J. Periodontal Res. 2015, 50, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, P.; Nelson, V.M.; Manley, S.; Wellner, M.; Floer, M.; Binion, D.G.; Shaker, R. Effect of curcumin on acidic pH-induced expression of IL-6 and IL-8 in human esophageal epithelial cells (HET-1A): Role of PKC, MAPKs, and NF-kappaB. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G388–G398. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Yang, C.H.; Li, T.T.; Zouboulis, C.C.; Hsu, H.C. Cell-free extracts of Propionibacterium acnes stimulate cytokine production through activation of p38 MAPK and Toll-like receptor in SZ95 sebocytes. Life Sci. 2015, 139, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Gawaz, M. Platelet-derived CXCL12 (SDF-1alpha): Basic mechanisms and clinical implications. J. Thromb. Haemost. 2013, 11, 1954–1967. [Google Scholar] [CrossRef]
- Takekoshi, T.; Ziarek, J.J.; Volkman, B.F.; Hwang, S.T. A locked, dimeric CXCL12 variant effectively inhibits pulmonary metastasis of CXCR4-expressing melanoma cells due to enhanced serum stability. Mol. Cancer Ther. 2012, 11, 2516–2525. [Google Scholar] [CrossRef]
- Lin, T.J.; Issekutz, T.B.; Marshall, J.S. SDF-1 induces IL-8 production and transendothelial migration of human cord blood-derived mast cells. Int. Arch. Allergy Immunol. 2001, 124, 142–145. [Google Scholar] [CrossRef]
- Scupoli, M.T.; Donadelli, M.; Cioffi, F.; Rossi, M.; Perbellini, O.; Malpeli, G.; Corbioli, S.; Vinante, F.; Krampera, M.; Palmieri, M.; et al. Bone marrow stromal cells and the upregulation of interleukin-8 production in human T-cell acute lymphoblastic leukemia through the CXCL12/CXCR4 axis and the NF-kappaB and JNK/AP-1 pathways. Haematologica 2008, 93, 524–532. [Google Scholar] [CrossRef]
- Lin, C.H.; Shih, C.H.; Lin, Y.C.; Yang, Y.L.; Chen, B.C. MEKK1, JNK, and SMAD3 mediate CXCL12-stimulated connective tissue growth factor expression in human lung fibroblasts. J. Biomed. Sci. 2018, 25, 19. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imani, A.; Panahipour, L.; dos Santos Sanches, N.; Wang, L.; Gruber, R. Platelet-Rich Fibrin Increases CXCL8 Expression in Gingival Fibroblasts. Biomedicines 2024, 12, 1326. https://doi.org/10.3390/biomedicines12061326
Imani A, Panahipour L, dos Santos Sanches N, Wang L, Gruber R. Platelet-Rich Fibrin Increases CXCL8 Expression in Gingival Fibroblasts. Biomedicines. 2024; 12(6):1326. https://doi.org/10.3390/biomedicines12061326
Chicago/Turabian StyleImani, Atefe, Layla Panahipour, Natalia dos Santos Sanches, Lei Wang, and Reinhard Gruber. 2024. "Platelet-Rich Fibrin Increases CXCL8 Expression in Gingival Fibroblasts" Biomedicines 12, no. 6: 1326. https://doi.org/10.3390/biomedicines12061326
APA StyleImani, A., Panahipour, L., dos Santos Sanches, N., Wang, L., & Gruber, R. (2024). Platelet-Rich Fibrin Increases CXCL8 Expression in Gingival Fibroblasts. Biomedicines, 12(6), 1326. https://doi.org/10.3390/biomedicines12061326