Varespladib Inhibits the Phospholipase A2 and Coagulopathic Activities of Venom Components from Hemotoxic Snakes


 , and
, and 
Abstract
1. Introduction
2. Experimental Section
2.1. Chemicals
2.2. LC with Parallel Nanofractionation and MS Detection
2.3. Phospholipase A2 Activity Assay
2.4. Plasma Coagulation Activity Assay
2.5. Correlation of Biological and MS Data
3. Results and Discussion
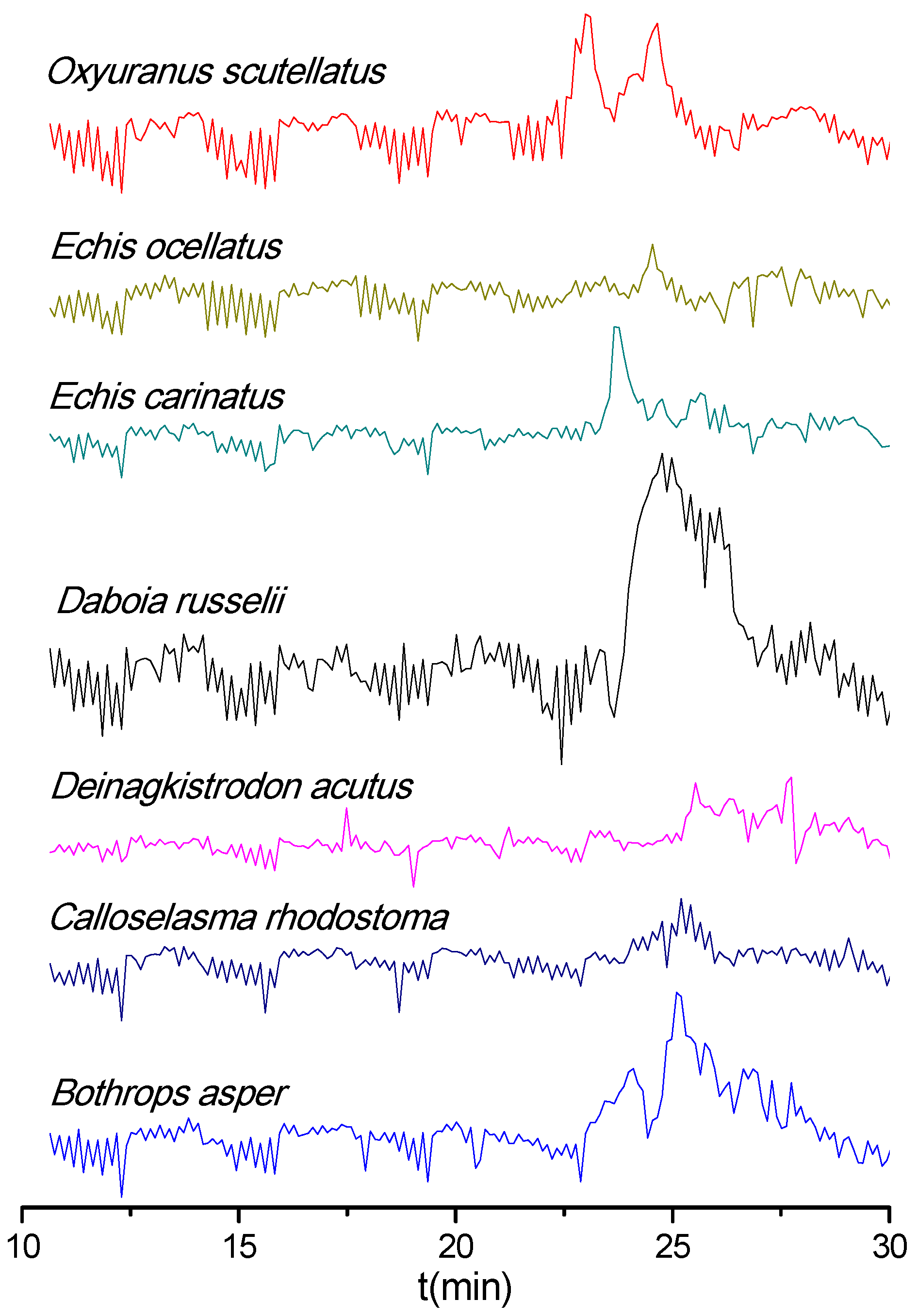
3.1. PLA2 Bioactivity Profiles of Nanofractionated Venom Toxins
3.2. Coagulopathic Bioactivity Profiles of Nanofractionated Venom Toxins
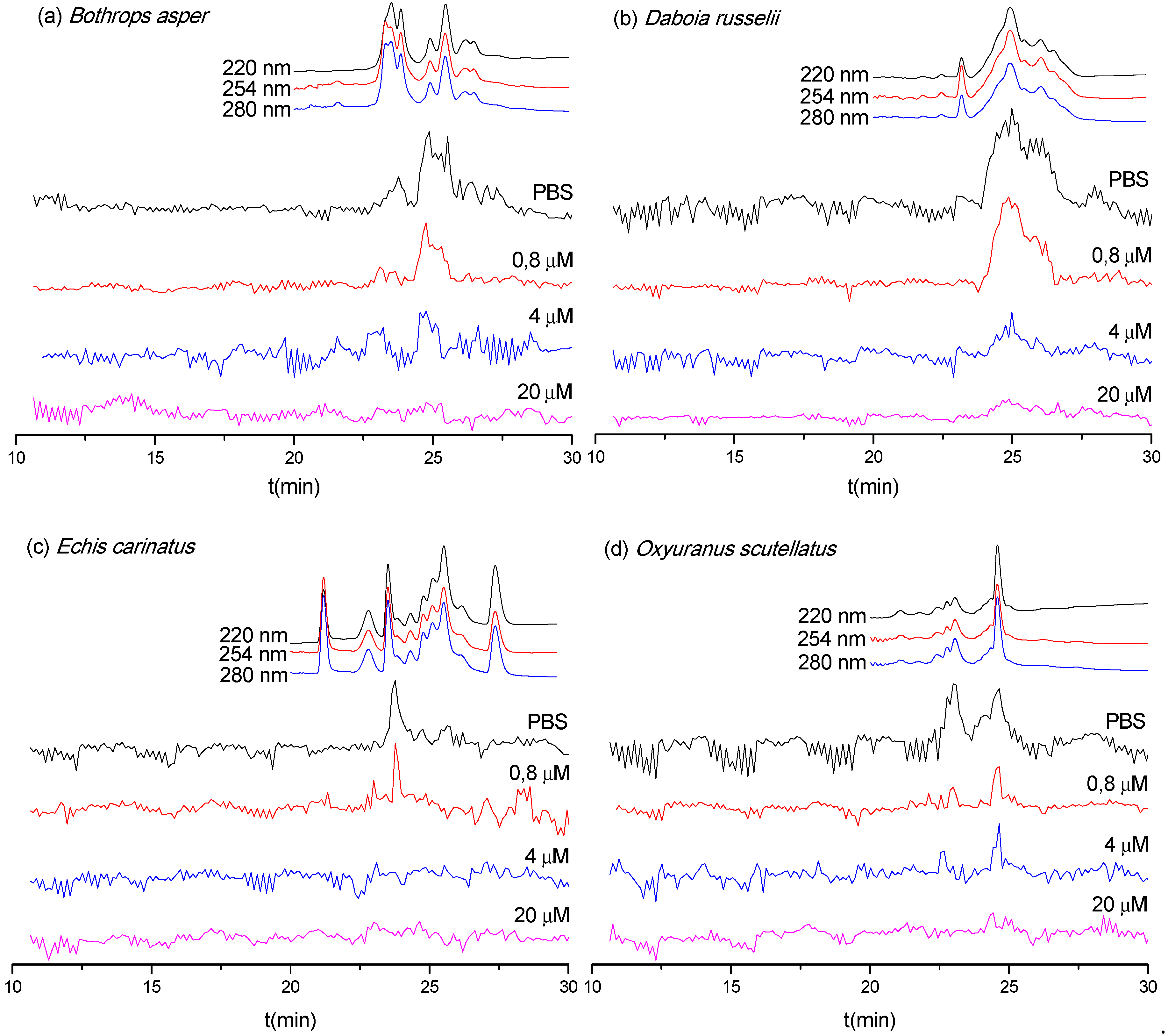
3.3. Neutralization Capabilities of Varespladib on the Enzymatic PLA2 Activity of Venom Toxins
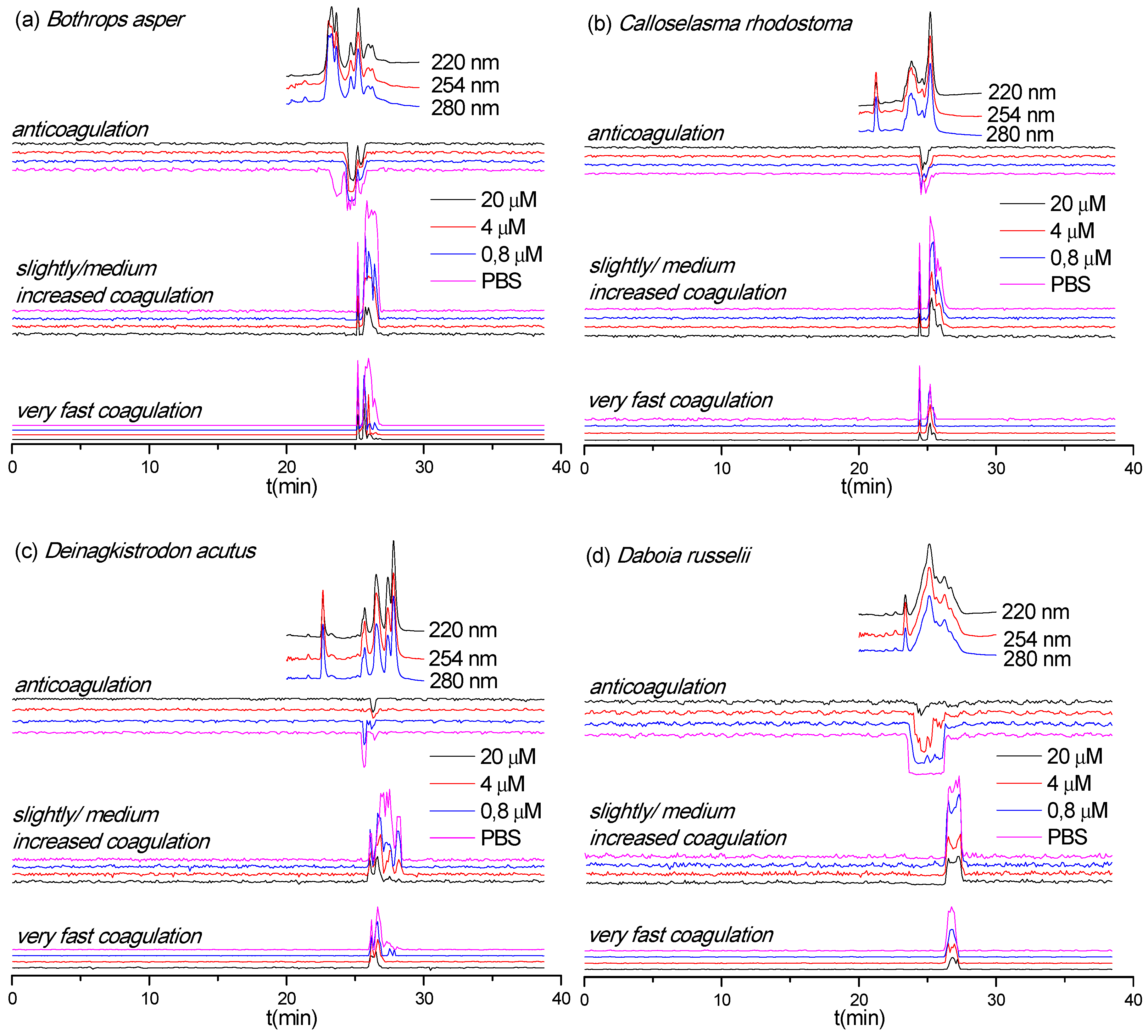
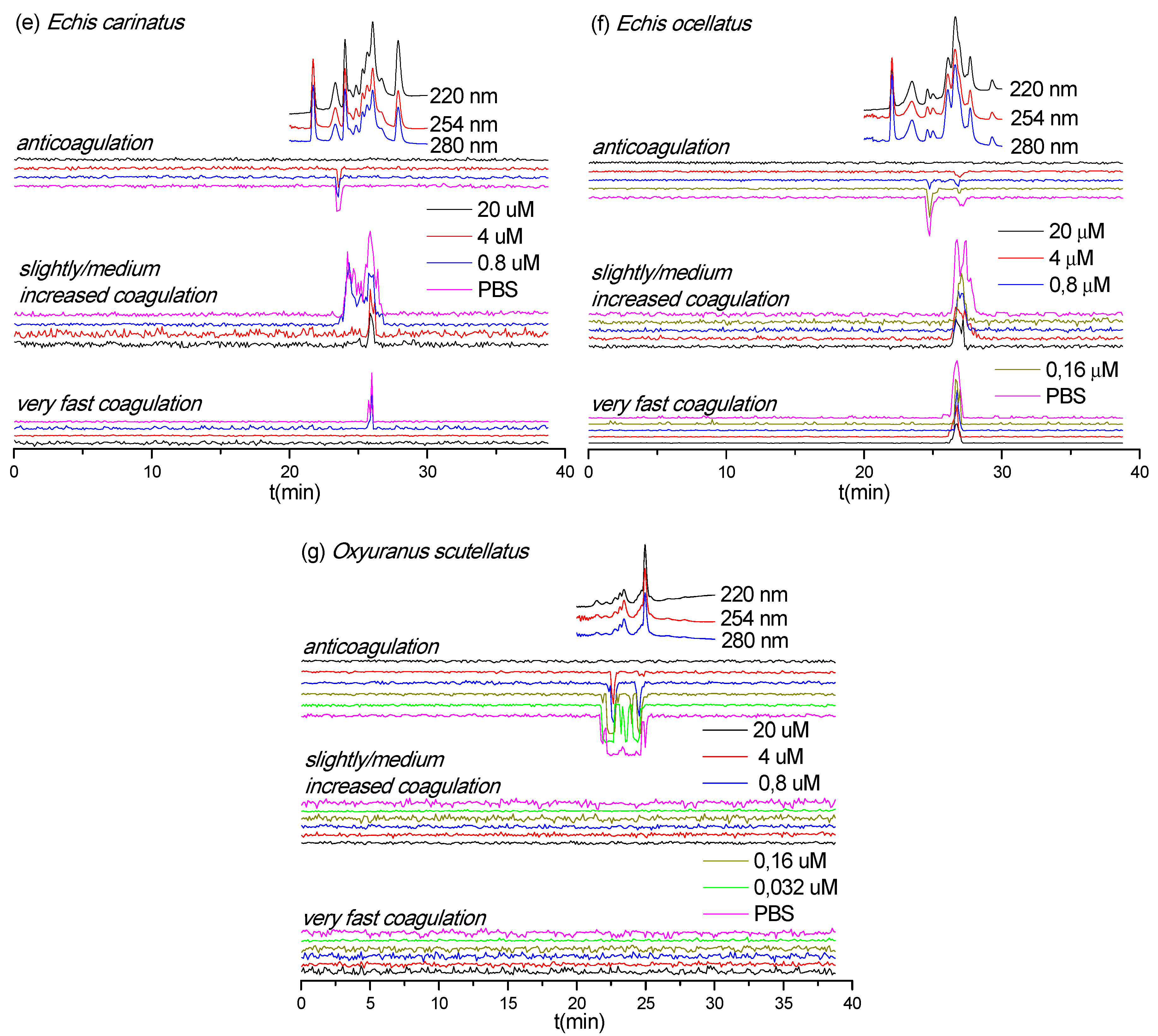
3.4. Neutralization Capabilities of Varespladib on Plasma Coagulation Activity of Venom Toxins
3.5. Identification of Venom Toxins Neutralized by Varespladib
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Price, J.A., III. A colorimetric assay for measuring phospholipase A2 degradation of phosphatidylcholine at physiological pH. J. Biochem. Biophys. Methods 2007, 70, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Dessen, A. Phospholipase A2 enzymes: Structural diversity in lipid messenger metabolism. Structure 2000, 8, R15–R22. [Google Scholar] [CrossRef]
- Hendrickson, H.S. Fluorescence-based assays of lipases, phospholipases, and other lipolytic enzymes. Anal. Biochem. 1994, 219, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tonello, F.; Rigoni, M. Cellular mechanisms of action of snake phospholipase A2 toxins. In Snake Venoms; Toxinology; Gopalakrishnakone, P., Inagaki, H., Vogel, C.W., Mukherjee, A., Rahmy, T., Eds.; Springer: Dordecht, The Netherlands, 2017; pp. 49–65. [Google Scholar]
- Costa, S.K.P.; Camargo, E.A.; Antunes, E. Inflammatory action of secretory phospholipases A2 from snake venoms. In Toxins and Drug Discovery; Toxinology; Gopalakrishnakone, P., Cruz, L., Luo, S., Eds.; Springer: Dordecht, The Netherlands, 2017; pp. 35–52. [Google Scholar]
- Cardoso, F.C.; Ferraz, C.R.; Arrahman, A.; Xie, C.; Casewell, N.R.; Lewis, R.J.; Kool, J. Multifunctional toxins in snake venoms and therapeutic implications: From pain to hemorrhage and necrosis. Front. Ecol. Evol. 2019, 7, 218–236. [Google Scholar]
- Panfoli, I.; Calzia, D.; Ravera, S.; Morelli, A. Inhibition of hemorragic snake venom components: Old and new approaches. Toxins 2010, 2, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Gutiérrez, J.M. Phospholipases A2 from viperidae snake venoms: How do they induce skeletal muscle damage? Acta Chim. Slov. 2011, 58, 647–659. [Google Scholar]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) appears to be a potent, broad-spectrum, inhibitor of snake venom phospholipase A2 and a possible pre-referral treatment for envenomation. Toxins 2016, 8, 248. [Google Scholar] [CrossRef]
- Williams, D.J.; Faiz, M.A.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PLoS Negl. Trop. Dis. 2019, 13, 7059–7080. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 1–21. [Google Scholar] [CrossRef]
- Ainsworth, S.; Slagboom, J.; Alomran, N.; Pla, D.; Alhamdi, Y.; King, S.I.; Bolton, F.M.; Gutiérrez, J.M.; Vonk, F.J.; Toh, C.-H.; et al. The paraspecific neutralisation of snake venom induced coagulopathy by antivenoms. Commun. Biol. 2018, 1, 34. [Google Scholar] [CrossRef]
- Bulfone, T.C.; Samuel, S.P.; Bickler, P.E.; Lewin, M.R. Developing small molecule therapeutics for the Initial and adjunctive treatment of snakebite. J. Trop. Med. 2018, 2018, 4320175. [Google Scholar] [CrossRef] [PubMed]
- Resiere, D.; Gutiérrez, J.M.; Névière, R.; Cabié, A.; Hossein, M.; Kallel, H. Antibiotic therapy for snakebite envenoming. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Albulescu, L.-O.; Hale, M.S.; Ainsworth, S.; Alsolaiss, J.; Crittenden, E.; Calvete, J.J.; Evans, C.; Wilkinson, M.C.; Harrison, R.A.; Kool, J.; et al. Preclinical validation of a repurposed metal chelator as an early-intervention therapeutic for hemotoxic snakebite. Sci. Transl. Med. 2020, 12, eaay8314. [Google Scholar] [CrossRef] [PubMed]
- Bittenbinder, M.A.; Zdenek, C.N.; Op den Brouw, B.; Youngman, N.J.; Dobson, J.S.; Naude, A.; Vonk, F.J.; Fry, B.G. Coagulotoxic cobras: Clinical implications of strong anticoagulant actions of African spitting Naja venoms that are not neutralised by antivenom but are by LY315920 (Varespladib). Toxins 2018, 10, 516. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Kastelein, J.J.; Schwartz, G.G.; Bash, D.; Rosenson, R.S.; Cavender, M.A.; Brennan, D.M.; Koenig, W.; Jukema, J.W.; Nambi, V.; et al. Varespladib and cardiovascular events in patients with an acute coronary syndrome: The VISTA-16 randomized clinical trial. JAMA 2014, 311, 252–262. [Google Scholar] [CrossRef]
- Shaposhnik, Z.; Wang, X.; Trias, J.; Fraser, H.; Lusis, A.J. The synergistic inhibition of atherogenesis in apoE−/− mice between pravastatin and the sPLA2 inhibitor varespladib (A-002). J. Lipid Res. 2009, 50, 623–629. [Google Scholar] [CrossRef]
- De Luca, D.; Minucci, A.; Piastra, M.; Cogo, P.E.; Vendittelli, F.; Marzano, L.; Gentile, L.; Giardina, B.; Conti, G.; Capoluongo, E.D. Ex vivo effect of varespladib on secretory phospholipase A2 alveolar activity in infants with ARDS. PLoS ONE 2012, 7, e47066. [Google Scholar] [CrossRef]
- Salvador, G.H.; Gomes, A.A.; Bryan-Quirós, W.; Fernández, J.; Lewin, M.R.; Gutiérrez, J.M.; Lomonte, B.; Fontes, M.R. Structural basis for phospholipase A2-like toxin inhibition by the synthetic compound Varespladib (LY315920). Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Cavender, M.A.; Kastelein, J.J.; Schwartz, G.; Waters, D.D.; Rosenson, R.S.; Bash, D.; Hislop, C. Inhibition of secretory phospholipase A2 in patients with acute coronary syndromes: Rationale and design of the vascular inflammation suppression to treat acute coronary syndrome for 16 weeks (VISTA-16) trial. Cardiovasc. Drugs Ther. 2012, 26, 71–75. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Elliott, M.; Stasiv, Y.; Hislop, C.; PLASMA II Investigators. Randomized trial of an inhibitor of secretory phospholipase A2 on atherogenic lipoprotein subclasses in statin-treated patients with coronary heart disease. Eur. Heart J. 2011, 32, 999–1005. [Google Scholar] [CrossRef]
- Adis, R.; Profile, D. Varespladib. Am. J. Cardiovasc. Drugs 2011, 11, 137–143. [Google Scholar]
- Nicholls, S. Varespladib trial terminated, increased MI risk. Reactions 2014, 1484, 1–18. [Google Scholar]
- Lewin, M.R.; Gutiérrez, J.M.; Samuel, S.P.; Herrera, M.; Bryan-Quirós, W.; Lomonte, B.; Bickler, P.E.; Bulfone, T.C.; Williams, D.J. Delayed oral LY333013 rescues mice from highly neurotoxic, lethal doses of Papuan Taipan (Oxyuranus scutellatus) venom. Toxins 2018, 10, 380. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, J.; Zhang, D.; Xiao, H.; Xiong, S.; Huang, C. Exploration of the inhibitory potential of varespladib for snakebite envenomation. Molecules 2018, 23, 391. [Google Scholar] [CrossRef] [PubMed]
- Bryan-Quirós, W.; Fernández, J.; Gutiérrez, J.M.; Lewin, M.R.; Lomonte, B. Neutralizing properties of LY315920 toward snake venom group I and II myotoxic phospholipases A2. Toxicon 2019, 157, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Lewin, M.R.; Williams, D.; Lomonte, B. Varespladib (LY315920) and methyl varespladib (LY333013) abrogate or delay lethality induced by presynaptically acting neurotoxic snake venoms. Toxins 2020, 12, 131. [Google Scholar] [CrossRef]
- Lewin, M.; Bulfone, T.; Samuel, S.; Gilliam, L. LY333013: A Candidate first-in-class, broad spectrum, oral field antidote to snakebite with curative potential. Ann. Emerg. Med. 2018, 72, S125. [Google Scholar] [CrossRef]
- Still, K.B.M.; Slagboom, J.; Kidwai, S.; Xie, C.; Zhao, Y.; Eisses, B.; Jiang, Z.; Vonk, F.J.; Somsen, G.W.; Casewell, N.R.; et al. Development of high-throughput screening assays for profiling snake venom phospholipase A2 activity after chromatographic fractionation. Toxicon 2020, 184, 28–38. [Google Scholar] [CrossRef]
- Still, K.B.M.; Nandlal, R.S.; Slagboom, J.; Somsen, G.W.; Casewell, N.R.; Kool, J. Multipurpose HTS coagulation analysis: Assay development and assessment of coagulopathic snake venoms. Toxins 2017, 9, 382. [Google Scholar] [CrossRef]
- Xie, C.; Slagboom, J.; Albulescu, L.-O.; Bruyneel, B.; Still, K.B.M.; Vonk, F.J.; Somsen, G.W.; Casewell, N.R.; Kool, J. Antivenom neutralization of coagulopathic snake venom toxins assessed by bioactivity profiling using nanofractionation analytics. Toxins 2020, 12, 53. [Google Scholar] [CrossRef]
- Slagboom, J.; Mladić, M.; Xie, C.; Kazandjian, T.D.; Vonk, F.; Somsen, G.W.; Casewell, N.R.; Kool, J. High throughput screening and identification of coagulopathic snake venom proteins and peptides using nanofractionation and proteomics approaches. PLoS Negl. Trop. Dis. 2020, 14, e0007802. [Google Scholar] [CrossRef]
- Kerns, R.T.; Kini, R.M.; Stefansson, S.; Evans, H.J. Targeting of venom phospholipases: The strongly anticoagulant phospholipase A2 from Naja nigricollis venom binds to coagulation factor Xa to inhibit the prothrombinase complex. Arch. Biochem. Biophys. 1999, 369, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Mounier, C.M.; Bon, C.; Kini, R.M. Anticoagulant venom and mammalian secreted phospholipases A2: Protein-versus phospholipid-dependent mechanism of action. Pathophysiol. Haemost. Thromb. 2001, 31, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M.; Evans, H.J. A model to explain the pharmacological effects of snake venom phospholipases A2. Toxicon 1989, 27, 613–635. [Google Scholar] [CrossRef]
- Zdenek, C.N.; Youngman, N.J.; Hay, C.; Dobson, J.; Dunstan, N.; Allen, L.; Milanovic, L.; Fry, B.G. Anticoagulant activity of black snake (Elapidae: Pseudechis) venoms: Mechanisms, potency, and antivenom efficacy. Toxicol. Lett. 2020, 330, 176–184. [Google Scholar] [CrossRef]
- Youngman, N.J.; Walker, A.; Naude, A.; Coster, K.; Sundman, E.; Fry, B.G. Varespladib (LY315920) neutralises phospholipase A2 mediated prothrombinase-inhibition induced by Bitis snake venoms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 236, 108818. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Peaks Retention Time (min) | Mascot Results Matching the Exact Mass | m/z Values from MS Data | Exact Mass from MS Data | Exact Mass Calculated from Mascot Data | Toxin Class | Varespladib Concentration Required for Full Inhibition |
|---|---|---|---|---|---|---|---|
| B. asper | 23.2–24.1 | PA2H2_BOTAS | 1373.368810+ | 13,714.5646 | 13,714.56817 | PLA2 | 0.8 μM |
| 24.3–25.8 | PA2HA_BOTAS | 1266.598511+ | 13,912.4649 | 13,896.51308 | PLA2 | 20 μM | |
| 24.3–25.8 | PA2H3_BOTAS | 1378.369710+ | 13,765.5812 | 13,765.58896 | PLA2 | 20 μM | |
| 24.3–25.8 | PA2B3_BOTAS | 1164.881112+ | 13,957.5333 | 13,957.48720 | PLA2 | 20 μM | |
| 24.3–25.8 | PA2A2_BOTAS | - | - | 14,194 | PLA2 | 20 μM | |
| D. russelii | 23.9–27.4 | PA2B8_DABRR | 1511.69629+ | 13,587.2248 | 13,587.2027 | PLA2 | 20 μM |
| 23.9–27.4 | PA2B5_DABRR | 13,587 | PLA2 | 20 μM | |||
| 23.9–27.4 | PA2B3_DABRR | - | - | 13,687 | PLA2 | 20 μM | |
| E. carinatus | |||||||
| 23.3–24.4 | PA2A1_ECHCA | - | - | 16,310 | PLA2 | 0.8 μM | |
| O. scutellatus | 22.6–25.1 | PA2TA_OXYSC | - | - | 13,829 | PLA2 | 20 μM |
| 22.6–25.1 | PA2TB_OXYSC | - | - | 16,008 | PLA2 | 20 μM | |
| 23.6–25.1 | PA21_OXYSC | - | - | 16,898 | PLA2 | 20 μM | |
| 23.6–25.1 | PA2TC_OXYSC | - | - | 13,313 | PLA2 | 20 μM |
| Species | Peak Retention Time (min) | Peak Activity | Mascot Results Matching the Exact Mass | m/z Values from MS Data | Exact Mass from MS Data | Exact Mass Calculated from Mascot Data | Toxin Class | Varespladib Concentration Needed for Full Inhibition |
|---|---|---|---|---|---|---|---|---|
| B. asper | 23.1–24.2 | Anticoagulation | PA2H2_BOTAS | 1373.368810+ | 13,714.5646 | 13,714.56817 | PLA2 | 0.8 μM |
| 24.2–25.2 | Anticoagulation | PA2HA_BOTAS | 1266.598511+ | 13,912.4649 | 13,896.51308 | PLA2 | 20 μM | |
| 24.2–25.2 | Anticoagulation | PA2H3_BOTAS | 1378.369710+ | 13,765.5812 | 13,765.58896 | PLA2 | 20 μM | |
| 25.2–25.8 | Anticoagulation | PA2B3_BOTAS | 1164.881112+ | 13,957.5333 | 13,957.48720 | PLA2 | 20 μM | |
| 25.2–25.8 | Anticoagulation | PA2A2_BOTAS | – | - | 14,194 | PLA2 | 20 μM | |
| 25.2–25.8 | Anticoagulation | VM2_BOTAS | - | - | 53,564 | SVMP | NOI | |
| 25.0–26.8 | Procoagulation | VSPL_BOTAS | - | - | 28,019 | SVSP | PN | |
| 25.0–26.8 | Procoagulation | VM1B1_BOTAS | - | - | 45,936 | SVMP | PN | |
| 25.4–26.8 | Procoagulation | SLA_BOTAS | - | 7084 | CTL | PN | ||
| C. rhodostoma | 24.3–25.5 | Anticoagulation | PA2BD_CALRH | 1244.110311+ | 13,665.0848 | 13,665.0237 | PLA2 | NOI |
| 24.3–25.5 | Anticoagulation | PA2AB_CALRH | - | - | 14,352 | PLA2 | NOI | |
| 24.3–25.5 | Anticoagulation | VSPF1_CALRH | - | - | 26,570 | SVSP | NOI | |
| 24.3–25.5 | Anticoagulation | SLEA_CALRH | - | - | 15,962 | CTL | NOI | |
| 24.3–25.5 | Anticoagulation | SLEB_CALRH | - | - | 15,190 | CTL | NOI | |
| 24.3–26.6 | Procoagulation | VSPF2_CALRH | - | - | 29,145 | SVSP | PN | |
| 24.9–26.6 | Procoagulation | SLYA_CALRH | - | - | 15,796 | CTL | PN | |
| 24.9–26.6 | Procoagulation | SLYB_CALRH | - | - | 16,770 | CTL | PN | |
| D. acutus | 25.4–25.9 | Anticoagulation | PA2A_DEIAC | - | - | 14,820 | PLA2 | 4 μM |
| 25.4–25.9 | Anticoagulation | SL_DEIAC | - | - | 18,332 | CTL | 4 μM | |
| 26.0–27.2 | Procoagulation | VSP1_DEIAC | - | - | 29,480 | SVSP | PN | |
| 26.0–27.2 | Procoagulation | VSPA_DEIAC | - | - | 26,132 | SVSP | PN | |
| 26.4–27.8 | Procoagulation | SLCB_DEIAC | - | - | 17,133 | CTL | PN | |
| 26.4–27.8 | Procoagulation | VM1AC_DEIAC | - | - | 47,690 | SVMP | PN | |
| 26.4–27.8 | Procoagulation | VM11_DEIAC | - | - | 47,845 | SVMP | PN | |
| 26.4–27.8 | Procoagulation | VM1H5_DEIAC | - | - | 46,518 | SVMP | PN | |
| 26.4–27.8 | Procoagulation | VM3AK_DEIAC | - | - | 69,752 | SVMP | PN | |
| 27.8–28.4 | Procoagulation | VM3A2_DEIAC | - | 27,151 | SVMP | 20 μM | ||
| 27.8–28.4 | Procoagulation | VM3AH_DEIAC | - | 70,721 | SVMP | 20 μM | ||
| D. russelii | 23.4–26.4 | Anticoagulation | PA2B8_DABRR | 1511.69629+ | 13,587.2248 | 13,587.2027 | PLA2 | 20 μM |
| 23.4–26.4 | Anticoagulation | PA2B5_DABRR | – | 13,587 | PLA2 | 20 μM | ||
| 23.4–26.4 | Anticoagulation | PA2B3_DABRR | – | 13,687 | PLA2 | 20 μM | ||
| 26.2–27.6 | Procoagulation | – | - | - | - | - | ||
| E. carinatus | 23.3–23.8 | Anticoagulation | PA2A1_ECHCA | - | - | 16,310 | PLA2 | 0.8 μM |
| 23.8–26.9 | Procoagulation | - | - | - | - | - | ||
| E. ocellatus | 24.4–25.1 | Anticoagulation | PA2A5_ECHOC | 1541.47189+ | 13,856.1382 | 13,856.0665 | PLA2 | 4 μM |
| 26.3–28.2 | Procoagulation | VM3E2_ECHOC | - | - | 69,426 | SVMP | PN | |
| 26.3–28.2 | Procoagulation | VM3E6_ECHOC | - | - | 57,658 | SVMP | PN | |
| 26.3–28.2 | Procoagulation | SL1_ECHOC | - | - | 16,601 | CTL | PN | |
| 26.3–28.2 | Procoagulation | SL124_ECHOC | - | - | 16,882 | CTL | PN | |
| O. scutellatus | 21.7–25.2 | Anticoagulation | PA2TA_OXYSC | - | - | 13,829 | PLA2 | 20 μM |
| 21.7–25.2 | Anticoagulation | PA2TB_OXYSC | - | - | 16,008 | PLA2 | 20 μM | |
| 21.7–25.2 | Anticoagulation | PA21_OXYSC | - | - | 16,898 | PLA2 | 20 μM | |
| 21.7–25.2 | Anticoagulation | PA2TC_OXYSC | - | - | 13,313 | PLA2 | 20 μM | |
| 21.7–25.2 | Anticoagulation | VKT_OXYSC | - | 9711 | kunitz | 20 μM | ||
| 21.7–25.2 | Anticoagulation | VKT3_OXYSC | - | - | 9029 | kunitz | 20 μM |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, C.; Albulescu, L.-O.; Still, K.B.M.; Slagboom, J.; Zhao, Y.; Jiang, Z.; Somsen, G.W.; Vonk, F.J.; Casewell, N.R.; Kool, J. Varespladib Inhibits the Phospholipase A2 and Coagulopathic Activities of Venom Components from Hemotoxic Snakes. Biomedicines 2020, 8, 165. https://doi.org/10.3390/biomedicines8060165
Xie C, Albulescu L-O, Still KBM, Slagboom J, Zhao Y, Jiang Z, Somsen GW, Vonk FJ, Casewell NR, Kool J. Varespladib Inhibits the Phospholipase A2 and Coagulopathic Activities of Venom Components from Hemotoxic Snakes. Biomedicines. 2020; 8(6):165. https://doi.org/10.3390/biomedicines8060165
Chicago/Turabian StyleXie, Chunfang, Laura-Oana Albulescu, Kristina B. M. Still, Julien Slagboom, Yumei Zhao, Zhengjin Jiang, Govert W. Somsen, Freek J. Vonk, Nicholas R. Casewell, and Jeroen Kool. 2020. "Varespladib Inhibits the Phospholipase A2 and Coagulopathic Activities of Venom Components from Hemotoxic Snakes" Biomedicines 8, no. 6: 165. https://doi.org/10.3390/biomedicines8060165
APA StyleXie, C., Albulescu, L.-O., Still, K. B. M., Slagboom, J., Zhao, Y., Jiang, Z., Somsen, G. W., Vonk, F. J., Casewell, N. R., & Kool, J. (2020). Varespladib Inhibits the Phospholipase A2 and Coagulopathic Activities of Venom Components from Hemotoxic Snakes. Biomedicines, 8(6), 165. https://doi.org/10.3390/biomedicines8060165