
Structural Changes of the Interface Material of Scallop Adductor under Ultra-High Pressure

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Materials
2.1.1. Experiment Raw Materials
2.1.2. Sample Pretreatment and Experimental Equipment
2.2. Experiment Methods
2.2.1. Raman Spectrum Analysis
2.2.2. Scanning Electron Microscopy Analysis
2.2.3. Fourier-Transform Infrared Spectrum Analysis
3. Results
3.1. Impacts on the Adductor Muscle Structure at the Bonding Interface of Scallops Produced by Ultra-High Pressure
Effect on the Structure of Adductor Muscle Interface
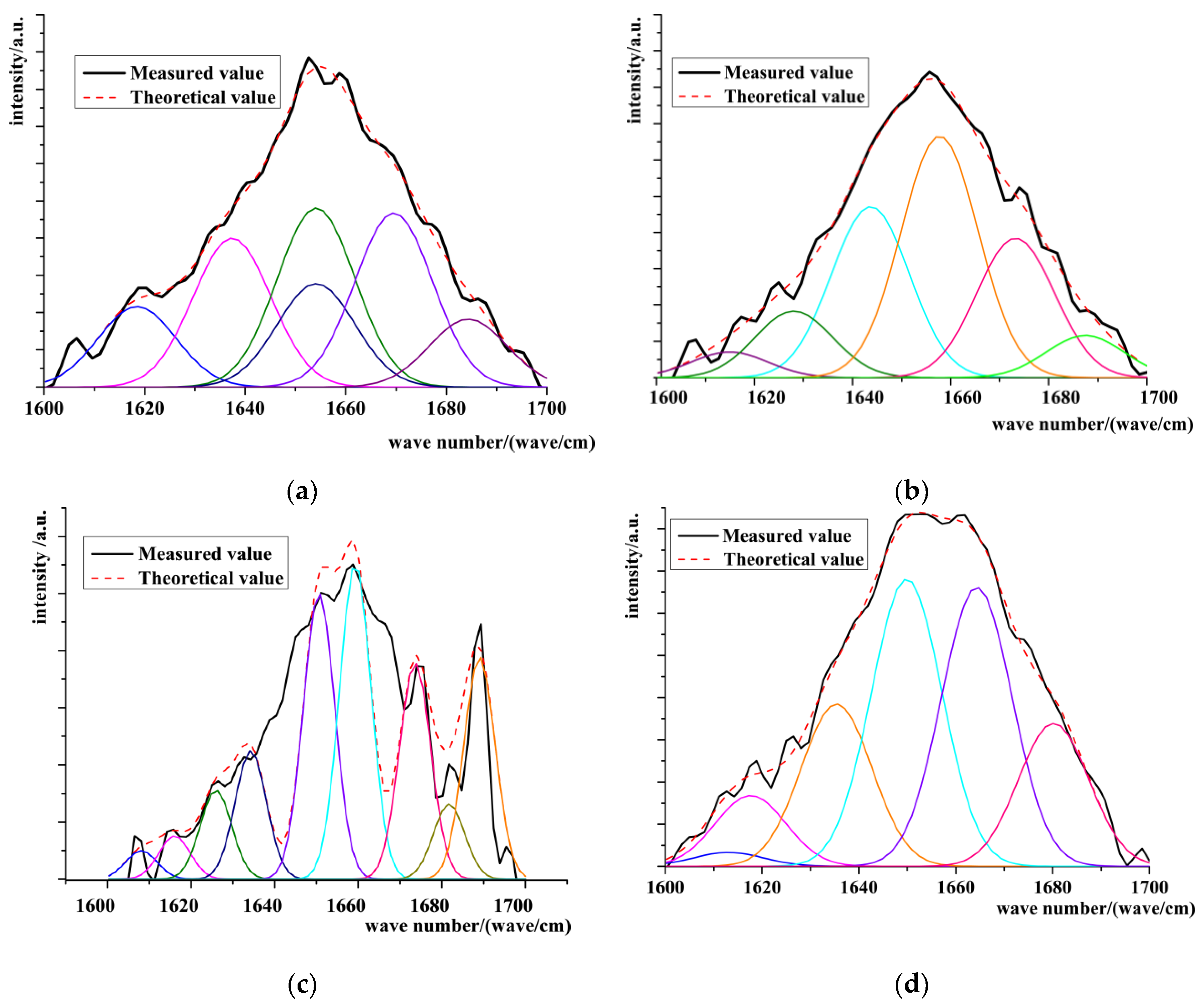
Raman Spectrum Analysis
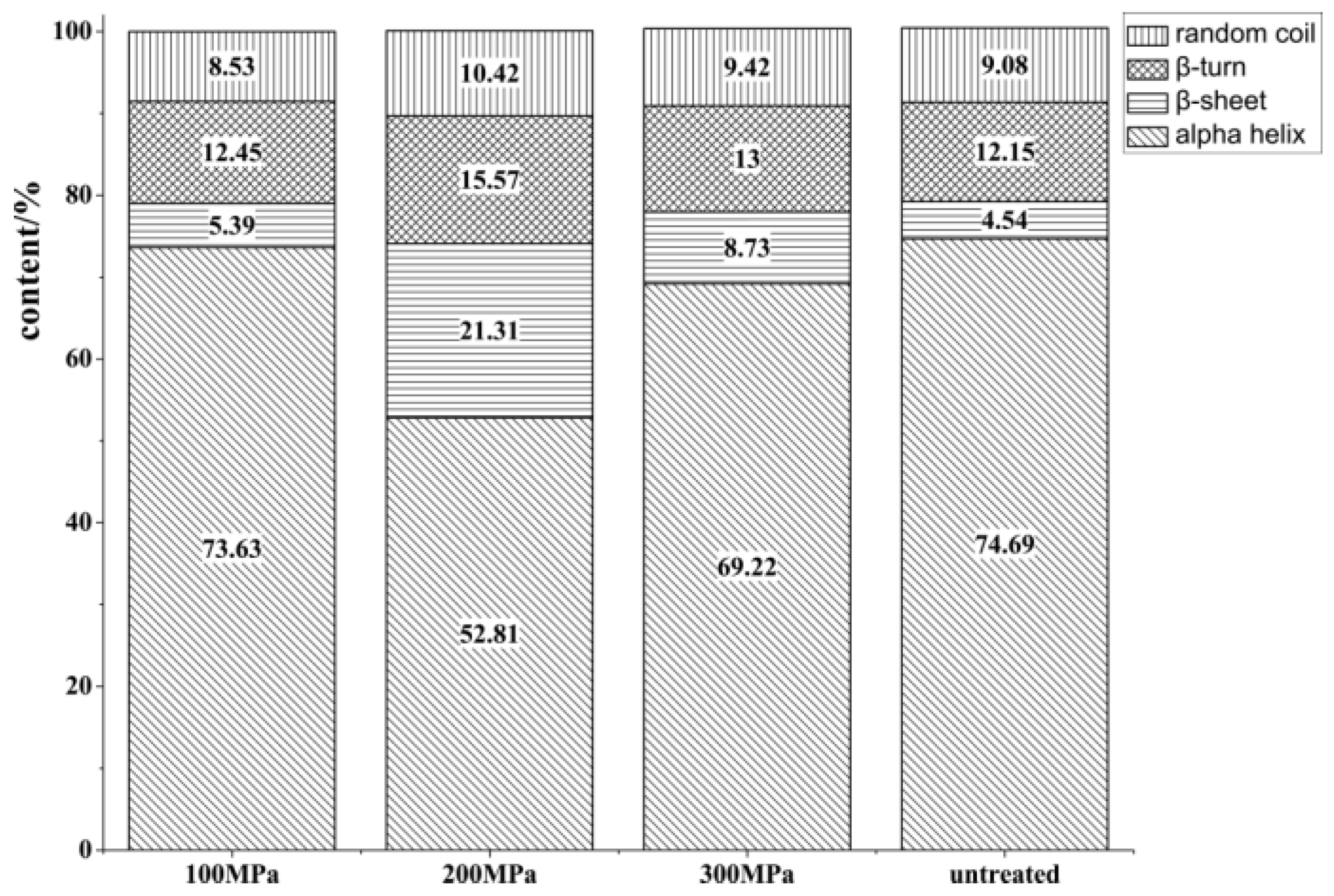
Changes in Secondary Structure
Scanning Electron Microscopy Analysis
3.2. Structural Changes of Organic–Inorganic Composites at the Interface of Scallop Shells under Ultra-High Pessure
3.2.1. Infrared Spectroscopic Analysis
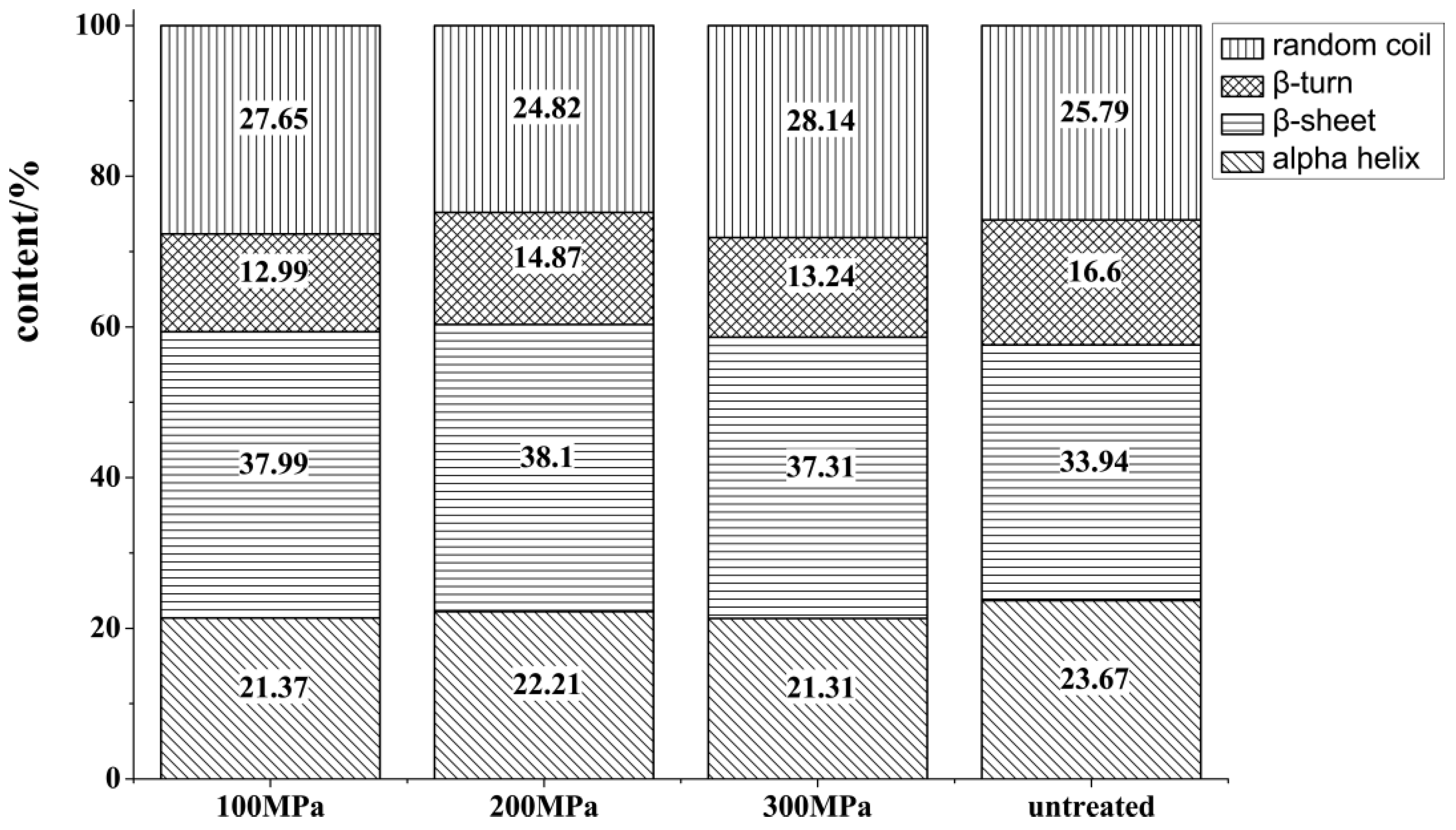
3.2.2. Secondary Structure Changes
3.2.3. Micro-Structure Analysis of the Inner Surface
4. Discussion and Conclusions
4.1. Contrasts to Previous Studies
4.2. Research Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.J.; Zhan, S.M.; Cao, P.L.; Liu, N.; Chen, X.H.; Wang, Y.J.; Wang, C.B. The polypeptide in Chlamys farreri can protect human dermal fibroblasts from ultraviolet B damage. Chin. J. Ocean Limn. 2005, 23, 357–362. [Google Scholar] [CrossRef]
- Fishery administration bureau of the ministry of agriculture and rural areas. In China Fishery Statistical Yearbook of 2020; China Agricultural Press: Beijing, China; pp. 1–50.
- Wang, Z.X.; Li, Y.S. Shellfish food and blood lipid. Foreign Med. Hyg. Vol. 1995, 6, 338–340. [Google Scholar]
- Li, W.Q.; Wang, J.; Sun, J.F.; Li, W.S. The nutrients analysis and evaluation of argopectens irradia. Acta Nutr. Sin. 2007, 33, 630–632. [Google Scholar]
- Wang, L. On the nutritional value, biological activity and culture of scallop. J. Mudanjiang Univ. 2007, 16, 92–94. [Google Scholar]
- Pham, H.; Shihoko, K.; Shin-Ichi, Y.; Ken-Ichi, O.; Koji, Y. Dietary effect of EPA rich and DHA rich fish oils on the immune function of rats. Biol. Sci. 2009, 16, 135–140. [Google Scholar] [CrossRef]
- Huang, X.C.; Liu, H.H.; Su, X.R. Study on the content of fatty acids in gonads of seven species of economic shellfish. Fish. Sci 2005, 8, 20–22. [Google Scholar] [CrossRef]
- Wang, X.J.; Liu, X.S.; Sun, K.; Yang, L.H.; Yao, M.Y.; Jiang, F.F. Extraction and analysis of nutritional components of chlamys farreri. Food Sci. 2007, 28, 271–275. [Google Scholar] [CrossRef]
- Susan, D.I.; Arin, K.; Mark, A.F.; Megan, L.; Kevin, S. Gray meat in the atlantic sea scallop, Placopecten magellanicus, and the identification of a known pathogenic scallop apicomplexan. J. Invert. Pathol. 2016, 141, 66–75. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Zheng, Y.; Liu, H.H.; Yan, L.; Zheng, Y.; Liu, H.; Zhang, Q.; Song, Y.; Tian, Y.Y.; Liu, J.R. Activity and characteristics of superoxide dismutase in adductor muscle of live dry—Storaged yesso scallop Patinopecten yessoensis. J. Dalian Ocean Univ. 2018, 33, 651–657. [Google Scholar] [CrossRef]
- Thomas, W.E. Understanding the counterrntuitive phenomenon of catch bonds. Curr. Nanosci. 2007, 3, 63–77. [Google Scholar] [CrossRef]
- Weiss, I.M.; Lüke, F.; Eichner, N.; Guth, C.; Clausen-Schaumann, H. On the function of chitin synthase extracellular domains in biomineralization. J. Srruct. Biol. 2013, 183, 216–225. [Google Scholar] [CrossRef] [PubMed]
- He, J.B.; Wen, S.L. Interface features of shell and closed-shell-muscle. J. Inorg. Mater. 1998, 13, 127–128. [Google Scholar] [CrossRef]
- Liao, Z.; Sun, Q.; Jiang, Y.T. Molecular composition and mechanism of muscle-shell attachment of shellfish. J. Zhejiang Ocean Univ. (Nat. Sci.) 2018, 37, 313–319. [Google Scholar]
- Zhao, L.P.; Xu, H.Z.; Chen, D.; Bao, L.F.; Fan, M.H.; Liao, Z. Microstructure and spectral analysis of Mytilus coruscus shell. J. Zhejiang Univ. (Sci. Ed.) 2015, 42, 339–346. [Google Scholar] [CrossRef]
- Chen, Q.H.; Wen, L.H.; Chen, X.; Fang, X.B.; Ling, J.G.; Xuan, X.T. Effect of ultra-high pressure assisted shelling on biochemical characteristics and structure of myofifibrillar protein in Patinopecten Yessoensis. Food Sci. 2021, 42, 102–107. [Google Scholar] [CrossRef]
- Benjakul, S.; Visessanguan, W.C.; Thongkaew, M.T. Comparative study on physicochemical changes of muscle proteins from some tropical fish during frozen storage. Food Res. Int. 2003, 36, 787–795. [Google Scholar] [CrossRef]
- Zhang, P.X.; Zhou, X.F.; Fang, Y. Raman spectra of vegetables and fruits at two excitation wavelengths. J. Light Scatt. 2004, 16, 136–140. [Google Scholar] [CrossRef]
- Michalczyk, M.; Surowka, K. Changes in protein fractions of rainbow trout (Oncorhynchus mykiss) gravads during production and storage. Food Chem. 2007, 104, 1006–1013. [Google Scholar] [CrossRef]
- An, X.L.; Li, Q.Z.; Liu, H.P. FTIR study of the interaction between bovine serum albumins and cetyltrimethyl ammonium bromide. J. Southwest Norm. Univ. 2005, 30, 699–702. [Google Scholar]
- Gao, W.H.; Ye, R.S.; Pan, T.T.; Zeng, X.A. Analysis of structural changes of surimi proteins during frozen storage by Raman spectroscopy. Food Sci. 2018, 39, 71–77. [Google Scholar] [CrossRef]
- Chen, H.Y.; Han, M.Y. Raman spectroscopic study of the effects of microbial transglutaminase on heat-induced gelation of pork myofibrillar proteins and its relationship with textural characteristics. Food Res. Int. 2011, 44, 1514–1520. [Google Scholar] [CrossRef]
- Bryant, R.N.; Pasteris, J.D.; Fike, D.A. Express: Variability in the raman spectrum of unpolished growth and fracture surfaces of pyrite due to laser heating and crystal orientation. Appl. Spectrosc. 2018, 72, 37. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.M.; Cambero, M.I.; Ordóñez, J.A.; Hoza, L.D.L.; Carmona, P. Raman spectroscopy study of the structural effect of microbial transglutaminase on meat systems and its relationship with textural characteristics. Food Chem. 2008, 109, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.Z.; Zhao, Q.Z.; Zhao, M.M.; Bao, Y.; Chun, C.; Jiao, Y.R. Structural evaluation of myofibrillar proteins during processing of Cantonese sausage by Raman spectroscopy. J. Agric. Food Chem. 2011, 59, 11070–11077. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Z.H.; Wang, Y.M.; Yong, X.; Chang, H.X. Effects of konjac glucomannan on heat-induced changes of physicochemical and structural properties of surimi gels. Food Res. Int. 2016, 83, 152–161. [Google Scholar] [CrossRef]
- Zhang, D.K.; Zhang, H.E.; Zhu, Y.J.; Yang, J.P.; Lei, Y.S.; Yang, H.; Lou, Y.J. Effect of high hydrostatic pressure treatment on myofifibrillar protein structure of cultured large yellow croaker. Food Sci. 2019, 40, 61–67. [Google Scholar] [CrossRef]
- Yang, H.; Lu, S.C.; Zhang, H.E.; Liu, L.J.; Qi, X.Y. Effects of high hydrostatic pressure processing on the flavor and quality of cultured Yellow croaker (Pseudosciaena crocea). Food Sci. 2014, 35, 244–249. [Google Scholar] [CrossRef]
- Sano, T.; Ohno, T.; Otsuka-Fuchino, H.; Matsumoto, J.J.; Tsuchiya, T. Carp natural actomyosin: Thermal denaturation mechanism. J. Food Sci. 1994, 59, 1002–1008. [Google Scholar] [CrossRef]
- Li, L.K.; Spector, A. Circular dichroism of beta-poly-L-lysine. J. Am. Chem. Soc. 1969, 91, 220–222. [Google Scholar] [CrossRef]
- Herrero, M.A. Raman spectroscopy a promising technique for quality assessment of meat and fish: A review. Food Chem. 2008, 107, 1642–1651. [Google Scholar] [CrossRef]
- Dauphin, Y. Infrared spectra and element al composition in recent biogenic calcites: Relationships between the υ4 band wave length and Sr and Mg concentrations. Appl. Spectrom. 1999, 53, 184–190. [Google Scholar] [CrossRef]
- Baldauf, N.; Rndriguez, R.L.; YousefLuis, A.E.; Rodriguez, S.E. Effect of selective growth media on the differentiation of Salmonella enterica serovars by Fouriertransform mid-infrared spectroscopy. J. Microbiol. Meth. 2007, 68, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Gang, L.; Ji, L.; Ke, S.; Wang, S.; Chen, L.; Liu, Y.; Huang, Q. Composition, secondary structure, and self-assembly of oat protein isolate. J. Agric. Food Chem. 2009, 57, 4522–4558. [Google Scholar] [CrossRef]
- Bao, L.F.; Gao, P.; Zhao, L.P. Microstructural characteristics and FTIR analysis of the shell from green mussel (Perna canaliculus). J. Zhejiang Ocean Univ. (Nat. Sci.) 2014, 33, 347–353. [Google Scholar] [CrossRef]
- He, P.; Chen, J.X.; Su, M.; Han, J.; Cheng, K. Analysis of chemical composition and structure characteristics of shells. Cecsc. J. 2015, 66, 450–454. [Google Scholar] [CrossRef]
- Liang, Y.; Zhao, J.; Wang, L. The fourier transform infrared spectroscopy(FT-IR) and thermal analysis of the mollusc shell. J. Min. Pet. Sci. 2007, 27, 12–16. [Google Scholar] [CrossRef]
- Shi, Y.G.; Guo, Q.Q.; Yang, X.W.; Liu, X.F.; Wang, S.J.; Zhu, H.; Zhang, N. FTIR analysis on the structure of soybean—Casein protein complex. Chin. J. Food 2018, 18, 225–231. [Google Scholar] [CrossRef]
- Ma, Y.F.; Qiao, L.; Feng, Q.L. Research progress on biomineralization mechanism of freshwater pearl. J. Inorg. Mater. 2012, 8, 109–116. [Google Scholar] [CrossRef]
- Xie, C.F.; Qiu, T.Q.; Lu, H.Q.; Yang, L.S. Morphology variation of calciumcarbonate crystal irradiated by ultrasonic. J. South China Univ. Technol. (Nat. Sci. Ed.) 2007, 35, 62–66. [Google Scholar]
- Lin, J.Y.; Ma, K.Y.; Bai, Z.Y.; Li, J.L. Molecular cloning and characterization of perlucin from the freshwater pearl mussel, Hyriposis cumingii. Gene 2013, 526, 210–216. [Google Scholar] [CrossRef]
- Evans, J.S. “Tuning in” to mollusk shell nacre-and prismatic–associated protein terminal sequence. Implications for bio mineralization and the construction of high performance inorganic-organic composites. Chem. Rev. 2008, 108, 4455–4462. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, X.L.; Wang, R.L.; Ma, Y.J.; Yang, J. Influence of storage microenvironment to the secondary structure in wheat proteins. Cereals Oils 2014, 27, 36–38. [Google Scholar]
- Gao, L.L.; Wang, Z.Y.; Rao, W.L.; Cao, L.C.; Zhang, D.Q. Molecular interaction analysis between collagen and chitosan blend film based on infrared spectroscopy. Trans. Chin. Soc. Agric. Eng. 2018, 34, 285–291. [Google Scholar] [CrossRef]
- Shi, L.; Xiong, G.Q.; Yin, T.; Ding, A.Z.; Li, X.; Wu, W.J.; Qiao, Y.; Liao, L.; Jiao, C.H.; Wang, L. Effects of ultra-high pressure treatment on the protein denaturation and water properties of red swamp crayfish (Procambarus clarkia). LWT 2020, 133, 110124. [Google Scholar] [CrossRef]
- Stupp, S.I.; Braun, P.V. Molecular manipulation of microstructures: Biomaterials, ceramics, and semiconductors. Nature 1996, 381, 56–58. [Google Scholar] [CrossRef]
- Ansboro, S.; Greiser, U.; Barry, F.; Murphy, M. Strategies for improved targeting of therapeutic cells: Implications for tissue repair. Eur. Cells Mater. 2012, 23, 310–318. [Google Scholar] [CrossRef]
- Surewicz, W.K.; Mantsch, H.H. New insight into protein secondary structure in water from second-derivative amide Ⅰ infrared spectra. Biochim. Bilphys. Acta 1988, 952, 115–130. [Google Scholar] [CrossRef]
- Zhu, C.; Chen, L.H.; Zeng, X.Y.; Sun, Y.; Jiao, D.X.; Liu, M.H.; Zheng, M.Z.; Liu, J.S.; Liu, H.M. Effects of ultra high pressure-magnetic field treatment on protein properties and quality characteristics of stored shrimp (Litopenaeus vannamei). LWT 2022, 170, 114070. [Google Scholar] [CrossRef]
- Choi, S.M.; Ma, C.Y. Structural characterization of globulin from common buckwheat (Fagopyrum esculentum Moench) using circular dichroism and Raman spectroscopy. Food Chem. 2007, 102, 150–160. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | The Wavelength of Characteristic Peak/Wave/cm | Functional Group | Corresponding Structure |
|---|---|---|---|
| 1 | 3435 | N-H | Stretching vibration |
| 2 | 2927 | -CH3- | Symmetric stretching vibration of anti -CH3- |
| 3 | 2519 | -SH- | Stretching vibration of -SH- |
| 4 | 2355 | O=C=O | Antisymmetric stretching vibration |
| 5 | 1789 | C=O | Stretching vibration of C=O |
| 6 | 1473 | O=C=O | Antisymmetric stretching vibration of CO32− |
| 7 | 1074 | O=C=O | Symmetric stretching vibration of CO32− |
| 8 | 864 | O=C=O | Out-of-plane bending vibration of CO32− |
| 9 | 707 | O=C=O | In-plane bending vibration of CO32− |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Chang, J.; Zhang, Y.; Li, D.; Xia, N.; Wang, J.; Sun, Z. Structural Changes of the Interface Material of Scallop Adductor under Ultra-High Pressure. Processes 2023, 11, 521. https://doi.org/10.3390/pr11020521
Gong X, Chang J, Zhang Y, Li D, Xia N, Wang J, Sun Z. Structural Changes of the Interface Material of Scallop Adductor under Ultra-High Pressure. Processes. 2023; 11(2):521. https://doi.org/10.3390/pr11020521
Chicago/Turabian StyleGong, Xue, Jiang Chang, Yinglei Zhang, Danting Li, Ning Xia, Jing Wang, and Zhihui Sun. 2023. "Structural Changes of the Interface Material of Scallop Adductor under Ultra-High Pressure" Processes 11, no. 2: 521. https://doi.org/10.3390/pr11020521
APA StyleGong, X., Chang, J., Zhang, Y., Li, D., Xia, N., Wang, J., & Sun, Z. (2023). Structural Changes of the Interface Material of Scallop Adductor under Ultra-High Pressure. Processes, 11(2), 521. https://doi.org/10.3390/pr11020521