
Antifungal Capacity of Microcapsules Containing Lactiplantibacillus plantarum TEP15 or Lactiplantibacillus pentosus TEJ4

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.1.1. Lactic Acid Bacteria
2.1.2. Phytopathogenic Fungi
2.2. Microencapsulation
2.3. Encapsulation Efficiency
2.4. Microcapsule Characterization
2.4.1. Moisture Content
2.4.2. Microcapsule Size
2.5. Storage for 28 Days
2.6. In Vitro Antifungal Activity of Microencapsulated Bacteria against Colletotrichum gloeosporioides, Fusarium sp., and Penicillium AT21H10
2.6.1. Evaluation against Mycelium
2.6.2. Evaluation against Spores
2.7. Data Analysis
3. Results and Discussion
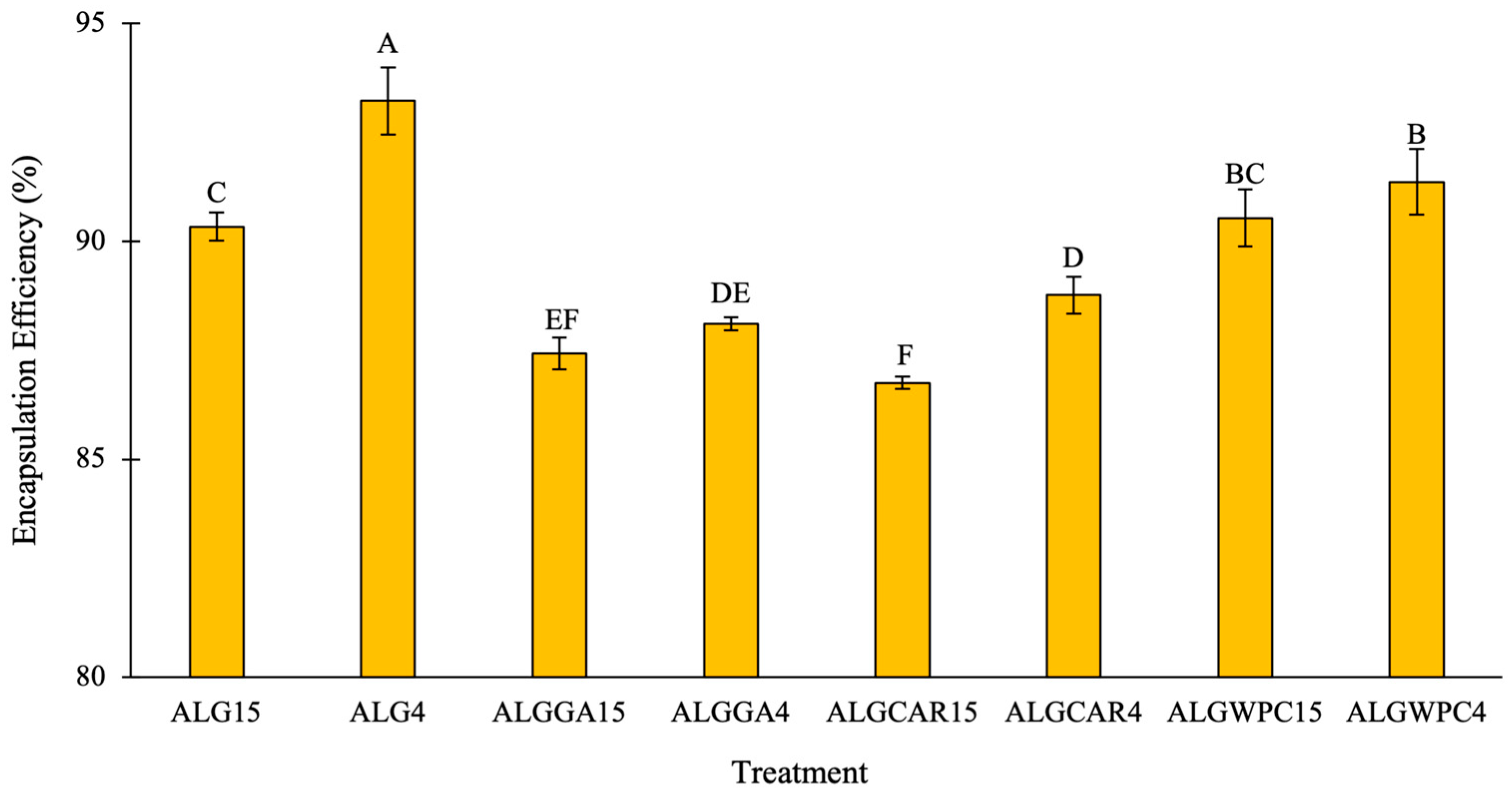
3.1. Encapsulation Efficiency
3.2. Moisture Content
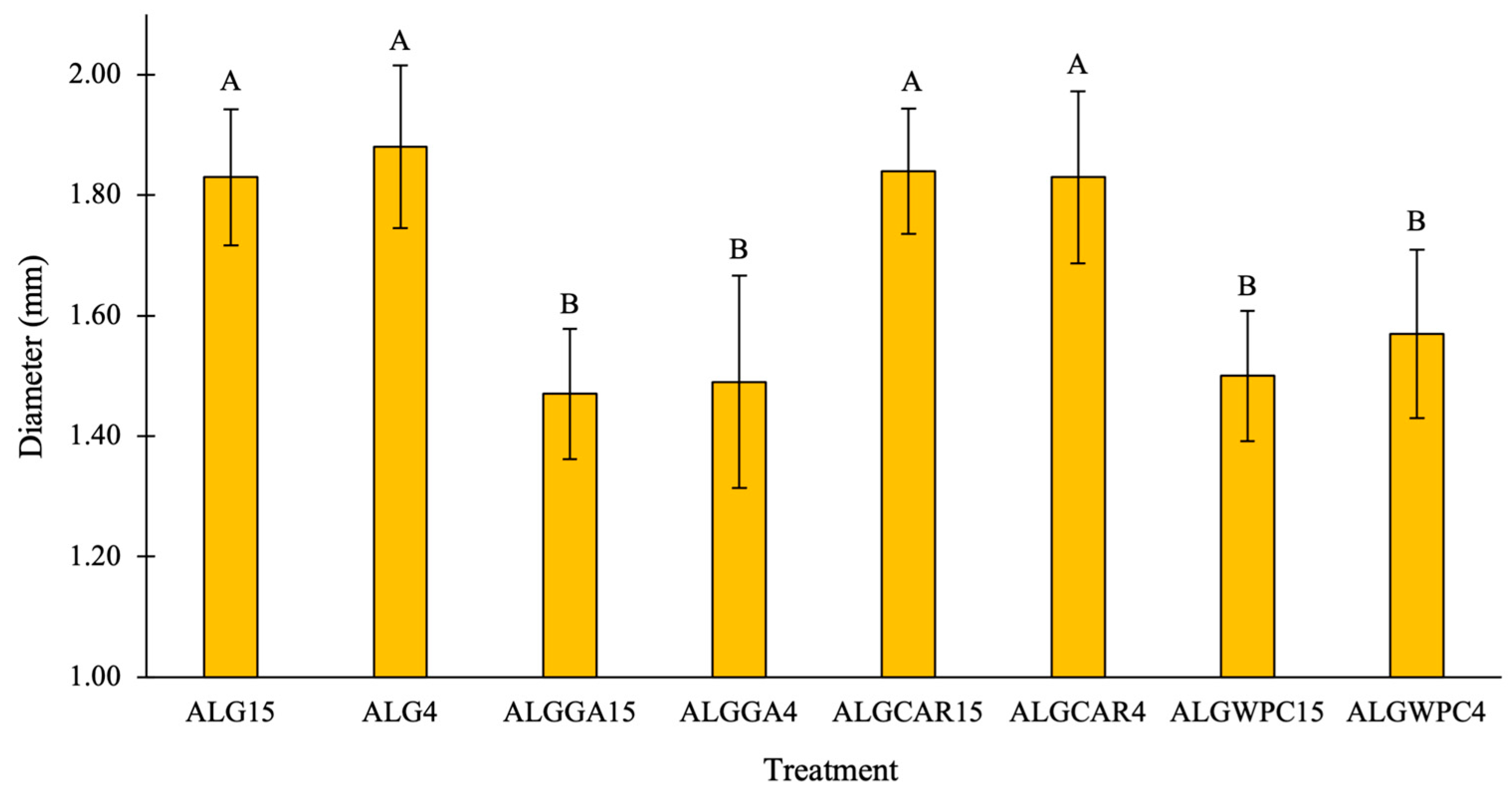
3.3. Microcapsule Size
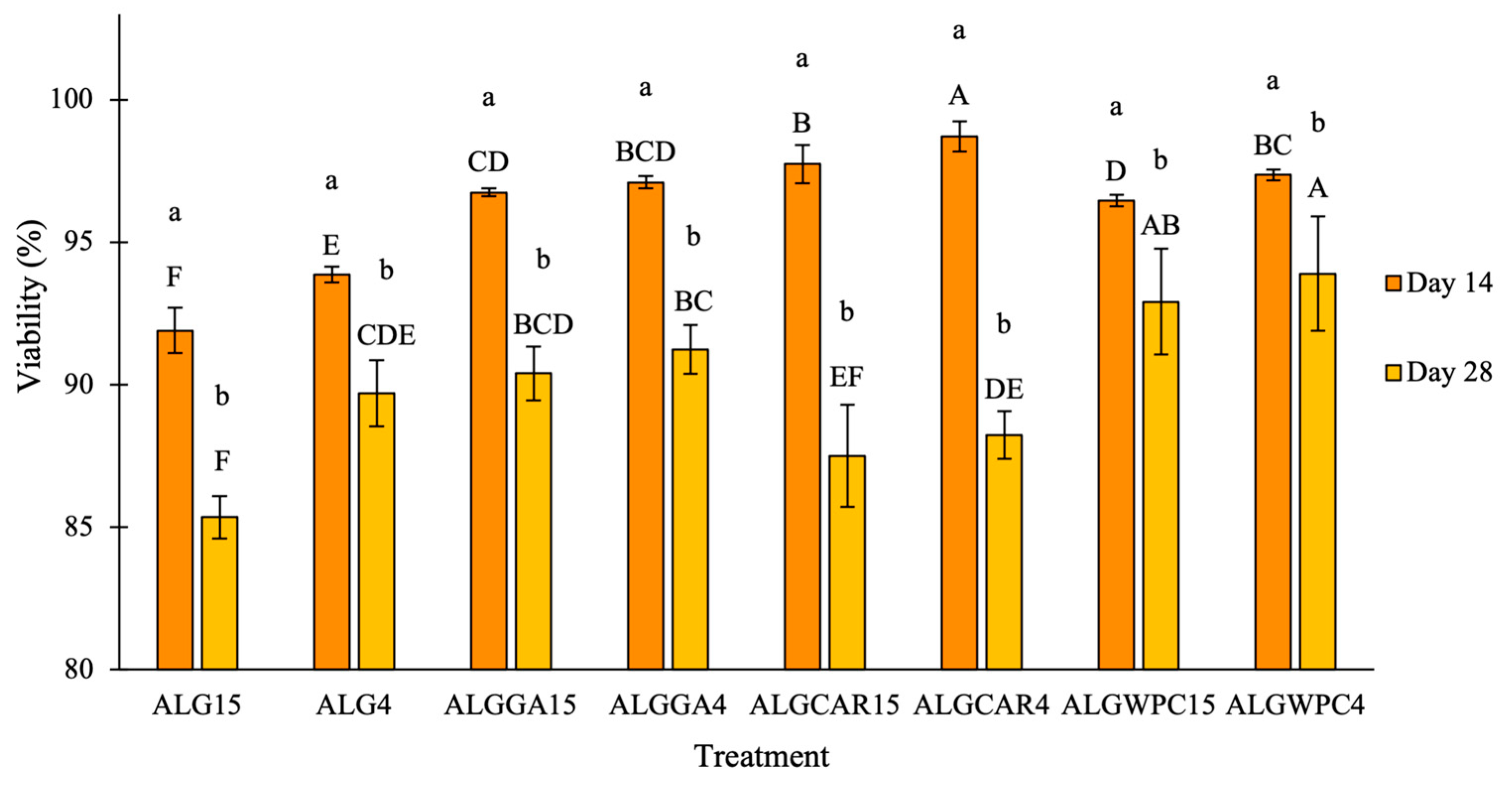
3.4. Survival Rate
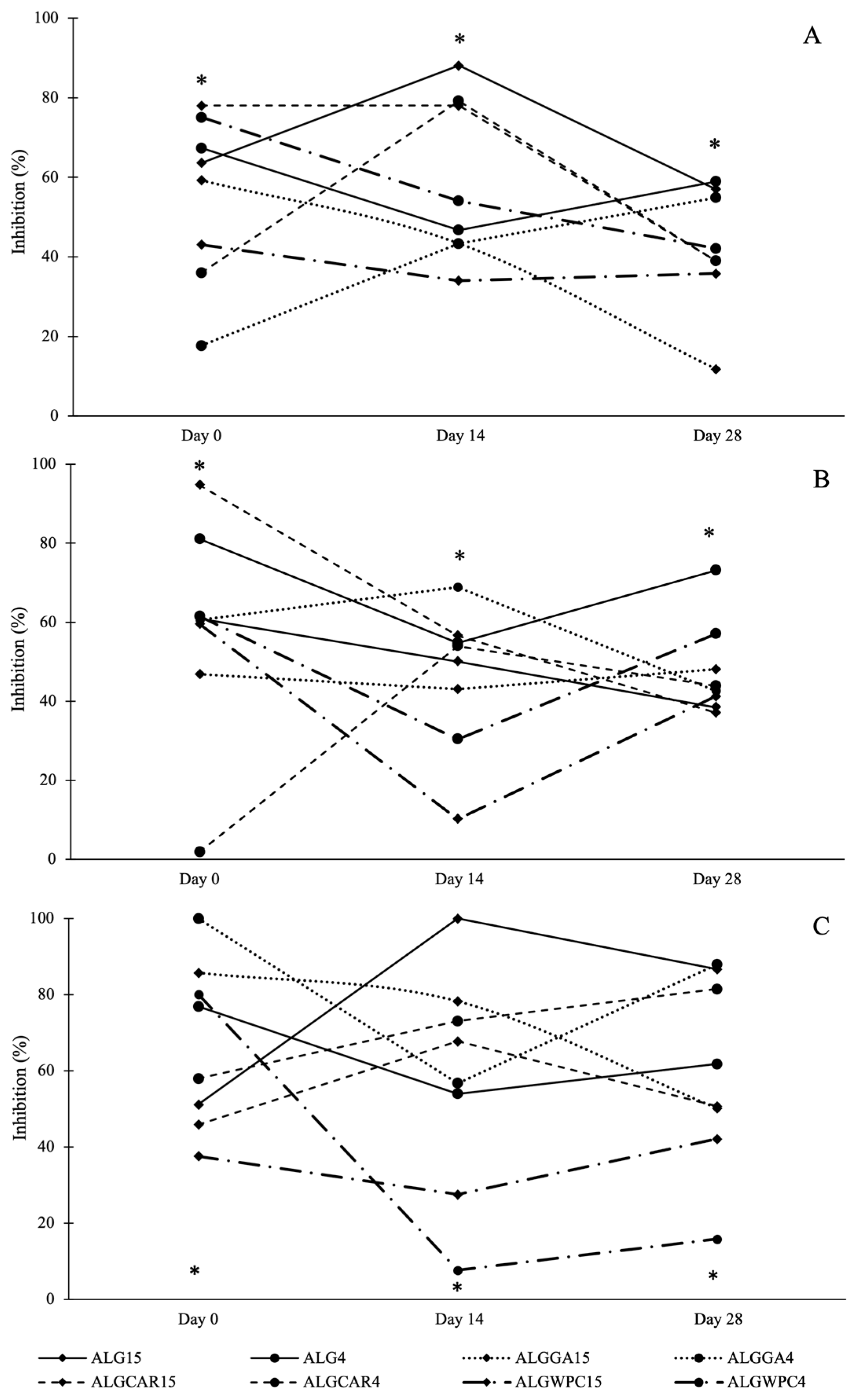
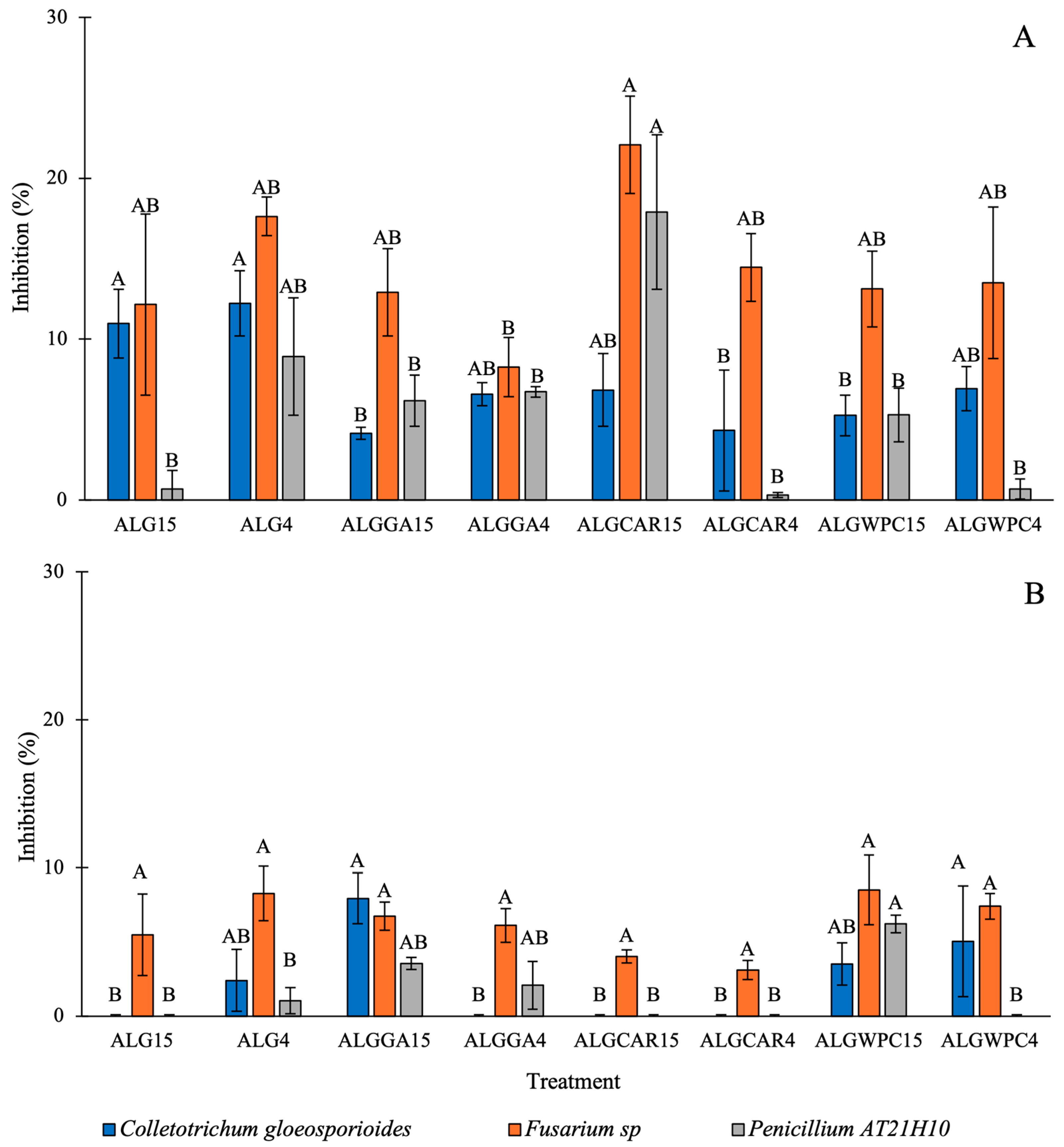
3.5. In Vitro Antifungal Activity against the Mycelium of Fungi
3.6. In Vitro Antifungal Activity against Spore Germination
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mohammadi, R.; Abbaszadeh, S.; Sharifzadeh, A.; Sepandi, M.; Taghdir, M.; Youseftabar, M.N.; Parastouei, K. In vitro activity of encapsulated lactic acid bacteria on aflatoxin production and growth of Aspergillus spp. Food Sci. Nutr. 2021, 9, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Dinu, S. Biocontrol of postharvest fungal diseases by microbial antagonists. Rom. J. Plant Prot. 2022, 15, 1–14. [Google Scholar] [CrossRef]
- Godana, E.A.; Yang, Q.; Zhang, X.; Zhao, L.; Wang, K.; Dhanasekaran, S.; Mehari, T.G.; Zhang, H. Biotechnological and biocontrol approaches for mitigating postharvest diseases caused by fungal pathogens and their mycotoxins in fruits: A review. J. Agric. Food Chem. 2023, 71, 17584–17596. [Google Scholar] [CrossRef] [PubMed]
- Ezzouggari, R.; Bahhou, J.; Taoussi, M.; Seddiqi, K.N.; Aberkani, K.; Barka, E.A.; Lahlali, R. Yeast warriors: Exploring the potential of yeasts for sustainable citrus post-harvest disease management. Agronomy 2024, 14, 288. [Google Scholar] [CrossRef]
- Hong, Y.; Ma, J.; Deng, L.; Yi, L.; Zeng, K. Screening and characterization of lactic acid bacteria with antifungal activity against Penicillium digitatum on citrus. Biol. Control 2019, 138, 104044. [Google Scholar]
- Gajbhiye, H.; Kapadnis, P. Antifungal activity producing lactic acid bacteria as biocontrol agents in plants. Biocontrol Sci. Technol. 2016, 26, 1451–1470. [Google Scholar] [CrossRef]
- López-Seijas, J.; García-Fraga, B.; da Silva, A.F.; Sieiro, C. Wine lactic acid bacteria with antimicrobial activity as potential biocontrol agents against Fusarium oxysporum f. sp. lycopersici. Agronomy 2019, 10, 31. [Google Scholar] [CrossRef]
- Akhavan-Kharazian, Z.; Salehi-Jouzani, G.; Aghdasi, M.; Khorvash, M.; Zamani, M.; Mohammadzadeh, H. Biocontrol potential of Lactobacillus strains isolated from corn silages against some plant pathogenic fungi. Biol. Control 2017, 110, 33–43. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggia, F.; Dalanaj, N.; Prodi, A.; Nipoti, P.; Pisi, A.; Di Gioia, D. Microbial inoculants for the biocontrol of Fusarium spp. in durum wheat. BMC Microbiol. 2015, 15, 242. [Google Scholar] [CrossRef]
- López-Pérez, B. Antifungal Spectrum of Chitosan Films Added with Cell-Free Supernatants of Lactic Acid Bacteria. Bachelor’s Thesis, Universidad Autónoma de Chiapas, Tapachula, Mexico, 2022. [Google Scholar]
- Nasrollahzadeh, A.; Mokhtari, S.; Khomeiri, M.; Saris, P.E. Antifungal preservation of food by lactic acid bacteria. Foods 2022, 11, 395. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic acid bacteria as antifungal and anti-mycotoxigenic agents: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef]
- Shehata, M.; Badr, A.; El Sohaimy, S.; Asker, D.; Awad, T. Characterization of antifungal metabolites produced by novel lactic acid bacterium and their potential application as food biopreservatives. Ann. Agric. Sci. 2019, 64, 71–78. [Google Scholar] [CrossRef]
- Fathi, F.; Saberi, R.; Khodaygan, P. Survivability and controlled release of alginate-microencapsulated Pseudomonas fluorescens VUPF506 and their effects on biocontrol of Rhizoctonia solani on potato. Int. J. Biol. Macromol. 2021, 183, 627–634. [Google Scholar] [CrossRef]
- Daranas, N.; Roselló, G.; Cabrefiga, J.; Donati, I.; Francés, J.; Badosa, E.; Bonaterra, A. Biological control of bacterial plant diseases with Lactobacillus plantarum strains selected for their broad-spectrum activity. Ann. Appl. Biol. 2019, 174, 92–105. [Google Scholar] [CrossRef]
- Saberi, R.; Moradi, M.; Mohammadinejad, R.; Thakur, V. Biopolymers for biological control of plant pathogens: Advances in microencapsulation of beneficial microorganisms. Polymers 2021, 13, 1938. [Google Scholar] [CrossRef]
- Balla, A.; Silini, A.; Cherif-Silini, H.; Chenari Bouket, A.; Alenezi, F.N.; Belbahri, L. Recent advances in encapsulation techniques of plant growth-promoting microorganisms and their prospects in the sustainable agriculture. Appl. Sci. 2022, 12, 9020. [Google Scholar] [CrossRef]
- Frakolaki, G.; Giannou, V.; Kekos, D.; Tzia, C. A review of the microencapsulation techniques for the incorporation of probiotic bacteria in functional foods. Crit. Rev. Food Sci. Nutr. 2021, 61, 1515–1536. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cano, B.; Mendoza-Meneses, C.J.; García-Trejo, J.F.; Macías-Bobadilla, G.; Aguirre-Becerra, H.; Soto-Zarazúa, G.M.; Feregrino-Pérez, A.A. Review and perspectives of the use of alginate as a polymer matrix for microorganisms applied in agro-industry. Molecules 2022, 27, 4248. [Google Scholar] [CrossRef]
- Wang, X.; Gao, S.; Yun, S.; Zhang, M.; Peng, L.; Li, Y.; Zhou, Y. Microencapsulating alginate-based polymers for probiotics delivery systems and their application. Pharmaceuticals 2022, 15, 644. [Google Scholar] [CrossRef]
- Sujarit, K.; Pathom-aree, W.; Mori, M.; Dobashi, K.; Shiomi, K.; Lumyong, S. Streptomyces palmae CMU-AB204T, an antifungal producing-actinomycete, as a potential biocontrol agent to protect palm oil producing trees from basal stem rot disease fungus, Ganoderma boninense. Biol. Control 2020, 148, 104307. [Google Scholar] [CrossRef]
- Le, N.; Bach, L.G.; Nguyen, D.C.; Le, T.H.; Pham, K.H.; Nguyen, D.H.; Hoang-Thi, T.T. Evaluation of factors affecting antimicrobial activity of bacteriocin from Lactobacillus plantarum microencapsulated in alginate-gelatin capsules and its application on pork meat as a bio-preservative. Int. J. Environ. Res. Public Health 2019, 16, 1017. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Tinajero, S.; Osorio-Hernández, E.; Estrada-Drouaillet, B.; Silva-Espinosa, J.H.T.; Delgado-Martínez, R.; Espinoza-Ahumada, C.A.; Ríos-Velasco, C.; Torres-Castillo, J.A. In vitro biocontrol of Fusarium oxysporum with antagonistic microorganisms and In vivo effect on Solanum lycopersicum L. J. Environ. Sci. Health Part B 2022, 57, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, X.; Cheng, J.; Nie, X.; Yu, X.; Zhao, Y.; Wang, W. Microencapsulation of Bacillus subtilis B99-2 and its biocontrol efficiency against Rhizoctonia solani in tomato. Biol. Control 2015, 90, 34–41. [Google Scholar] [CrossRef]
- Barrios-Roblero, C.; Rosas-Quijano, R.; Salvador-Figueroa, M.; Gálvez-López, D.; Vázquez-Ovando, A. Antifungal lactic acid bacteria isolated from fermented beverages with activity against Colletotrichum gloeosporioides. Food Biosci. 2019, 29, 47–54. [Google Scholar] [CrossRef]
- Frakolaki, G.; Giannou, V.; Topakas, E.; Tzia, C. Effect of various encapsulating agents on the beads’ morphology and the viability of cells during BB-12 encapsulation through extrusion. J. Food Eng. 2021, 294, 110423. [Google Scholar] [CrossRef]
- Sandoval-Mosqueda, I.; Llorente-Bousquets, A.; Montiel-Sosa, J.F.; Corona, L.; Guadarrama-Álvarez, Z. Encapsulation of Lactobacillus plantarum ATCC 8014 and Pediococcus acidilactici ATCC 8042 in a freeze-dried alginate-gum arabic system and its in vitro testing under gastrointestinal conditions. J. Microencapsul. 2019, 36, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Batalha, L.S.; Gontijo, M.T.P.; de Carvalho-Teixeira, A.V.N.; Boggione, D.M.G.; Lopez, M.E.S.; Eller, M.R.; Mendonça, R.C.S. Encapsulation in alginate-polymers improves stability and allows controlled release of the UFV-AREG1 bacteriophage. Food Res. Int. 2021, 139, 109947. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Saeed, M.; Ahmad, T.; Yamin, I.; Khan, W.A.; Iqbal, M.W.; Riaz, T. Characterization of biopolymeric encapsulation system for improved survival of Lactobacillus brevis. J. Food Meas. Charact. 2022, 16, 2292–2299. [Google Scholar] [CrossRef]
- Praepanitchai, O.A.; Noomhorm, A.; Anal, A.K. Survival and behavior of encapsulated probiotics (Lactobacillus plantarum) in calcium alginate soy protein isolate-based hydrogel beads in different processing conditions (pH and temperature) and in pasteurized mango juice. Food Microbiol. 2019, 2019, 9768152. [Google Scholar] [CrossRef]
- Parsana, Y.; Yadav, M.; Kumar, S. Microencapsulation in the chitosan-coated alginate-inulin matrix of Limosilactobacillus reuteri SW23 and Lactobacillus salivarius RBL50 and their characterization. Carbohydr. Polym. Technol. Appl. 2023, 5, 100285. [Google Scholar] [CrossRef]
- Castro-Rosas, J.; Gómez-Aldapa, C.A.; Chávez-Urbiola, E.A.; Hernández-Bautista, M.; Rodríguez-Marín, M.L.; Cabrera-Canales, Z.E.; Falfán-Cortés, R.N. Characterisation, storage viabilit, and application of microspheres with Lactobacillus paracasei obtained by the extrusion technique. Int. J. Food Sci. Technol. 2021, 56, 1809–1817. [Google Scholar] [CrossRef]
- Petraitytė, S.; Šipailienė, A. Enhancing encapsulation efficiency of alginate capsules containing lactic acid bacteria by using different divalent cross-linkers sources. LWT 2019, 110, 307–315. [Google Scholar] [CrossRef]
- Avila-Reyes, S.V.; Garcia-Suarez, F.J.; Jiménez, M.T.; San Martín-Gonzalez, M.F.; Bello-Perez, L.A. Protection of L. rhamnosus by spray-drying using two prebiotics colloids to enhance the viability. Carbohydr. Polym. 2014, 102, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Michel-Aceves, A.C.; Hernández-Morales, J.; Toledo-Aguilar, R.; Sabino López, J.E.; Romero-Rosales, T. Antagonistic capacity of native Trichoderma spp. against Phytophthora parasitica and Fusarium oxysporum isolated from roselle crops. Rev. Fitotec. Mex. 2019, 42, 235–241. [Google Scholar]
- Szopa, D.; Mielczarek, M.; Skrzypczak, D.; Izydorczyk, G.; Mikula, K.; Chojnacka, K.; Witek-Krowiak, A. Encapsulation efficiency and survival of plant growth-promoting microorganisms in an alginate-based matrix—A systematic review and protocol for a practical approach. Ind. Crops Prod. 2022, 181, 114846. [Google Scholar] [CrossRef]
- Li, P.; Müller, M.; Chang, M.W.; Frettlöh, M.; Schönherr, H. Encapsulation of autoinducer sensing reporter bacteria in reinforced alginate-based microbeads. ACS Appl. Mater. Interfaces 2017, 9, 22321–22331. [Google Scholar] [CrossRef] [PubMed]
- Bagdat, E.S.; Akman, P.K.; Kutlu, G.; Tornuk, F. Optimization of spray-drying process parameters for microencapsulation of three probiotic lactic acid bacteria selected by their high viability rate in sucrose and fructose levels and high temperatures. Syst. Microbiol. Biomanuf. 2023, 4, 687–698. [Google Scholar] [CrossRef]
- Ferrando, V.; Quiberoni, A.; Reinhemer, J.; Suárez, V. Resistance of functional Lactobacillus plantarum strains against food stress conditions. Food Microbiol. 2015, 48, 63–71. [Google Scholar] [CrossRef]
- Nezamdoost-Sani, N.; Khaledabad, M.A.; Amiri, S.; Khaneghah, A.M. Alginate and derivatives hydrogels in encapsulation of probiotic bacteria: An updated review. Food Biosci. 2023, 52, 102433. [Google Scholar] [CrossRef]
- Afzaal, M.; Khan, A.U.; Saeed, F.; Arshad, M.S.; Khan, M.A.; Saeed, M.; Anjum, F.M. Survival and stability of free and encapsulated probiotic bacteria under simulated gastrointestinal conditions and in ice cream. Food Sci. Nutr. 2020, 8, 1649–1656. [Google Scholar] [CrossRef]
- Goy, R.C.; Britto, D.D.; Assis, O.B. A review of the antimicrobial activity of chitosan. Polímeros 2019, 19, 241–247. [Google Scholar] [CrossRef]
- Yasmin, I.; Saeed, M.; Pasha, I.; Zia, M.A. Development of whey protein concentrate-pectin-alginate based delivery system to improve survival of B. longum BL-05 in simulated gastrointestinal conditions. Probiotics Antimicrob. Proteins 2019, 11, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Rajam, R.; Karthik, P.; Parthasarathi, S.; Joseph, G.S.; Anandharamakrishnan, C. Effect of whey protein–alginate wall systems on survival of microencapsulated Lactobacillus plantarum in simulated gastrointestinal conditions. J. Funct. Foods 2012, 4, 891–898. [Google Scholar] [CrossRef]
- Poddar, D.; Das, S.; Jones, G.; Palmer, J.; Jameson, G.B.; Haverkamp, R.G.; Singh, H. Stability of probiotic Lactobacillus paracasei during storage as affected by the drying method. Int. Dairy J. 2014, 39, 1–7. [Google Scholar] [CrossRef]
- Liu, H.; Cui, S.W.; Chen, M.; Li, Y.; Liang, R.; Xu, F.; Zhong, F. Protective approaches and mechanisms of microencapsulation to the survival of probiotic bacteria during processing, storage and gastrointestinal digestion: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2863–2878. [Google Scholar] [CrossRef] [PubMed]
- Rizo-Vázquez, F.; Vázquez-Ovando, A.; Mejía-Reyes, D.; Gálvez-López, D.; Rosas-Quijano, R. Use of lactulose as prebiotic and chitosan coating for improvement the viability of Lactobacillus sp. FM4. C1. 2 microencapsulate with alginate. Processes 2024, 12, 133. [Google Scholar] [CrossRef]
- Poletto, G.; Raddatz, G.C.; Cichoski, A.J.; Zepka, L.Q.; Lopes, E.J.; Barin, J.S.; de Menezes, C.R. Study of viability and storage stability of Lactobacillus acidophillus when encapsulated with the prebiotics rice bran, inulin and Hi-maize. Food Hydrocoll. 2019, 95, 238–244. [Google Scholar] [CrossRef]
- Gul, O.; Dervisoglu, M. Application of multicriteria decision technique to determine optimum sodium alginate concentration for microencapsulation of Lactobacillus casei Shirota by extrusion and emulsification. J. Food Process Eng. 2017, 40, e12481. [Google Scholar] [CrossRef]
- Altamirano-Ríos, A.V.; Guadarrama-Lezama, A.Y.; Arroyo-Maya, I.J.; Hernández-Álvarez, A.J.; Orozco-Villafuerte, J. Effect of encapsulation methods and materials on the survival and viability of Lactobacillus acidophilus: A review. Int. J. Food Sci. Technol. 2022, 57, 4027–4040. [Google Scholar] [CrossRef]
- Fareez, I.M.; Lim, S.M.; Mishra, R.K.; Ramasamy, K. Chitosan coated alginate–xanthan gum bead enhanced pH and thermotolerance of Lactobacillus plantarum LAB12. Int. J. Biol. Macromol. 2015, 72, 1419–1428. [Google Scholar] [CrossRef]
- Luca, L.; Oroian, M. Influence of different prebiotics on viability of Lactobacillus casei, Lactobacillus plantarum and Lactobacillus rhamnosus encapsulated in alginate microcapsules. Foods 2021, 10, 710. [Google Scholar] [CrossRef]
- Ta, L.P.; Bujna, E.; Antal, O.; Ladányi, M.; Juhász, R.; Szécsi, A.; Nguyen, Q.D. Effects of various polysaccharides (alginate, carrageenan, gums, chitosan) and their combination with prebiotic saccharides (resistant starch, lactosucrose, lactulose) on the encapsulation of probiotic bacteria Lactobacillus casei 01 strain. Int. J. Biol. Macromol. 2021, 183, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, Z.; Bai, L.; Deng, J.; Zhou, Q. Biomaterial-based encapsulated probiotics for biomedical applications: Current status and future perspectives. Mater. Design 2021, 210, 110018. [Google Scholar] [CrossRef]
- Razavi, S.; Janfaza, S.; Tasnim, N.; Gibson, D.L.; Hoorfar, M. Microencapsulating polymers for probiotics delivery systems: Preparation, characterization, and applications. Food Hydrocoll. 2021, 120, 106882. [Google Scholar] [CrossRef]
- Saberi, R.R.; Skorik, Y.A.; Thakur, V.K.; Moradi, P.M.; Tamanadar, E.; Noghabi, S.S. Encapsulation of plant biocontrol bacteria with alginate as a main polymer material. Int. J. Mol. Sci. 2021, 22, 11165. [Google Scholar] [CrossRef]
- Frent, O.D.; Vicas, L.G.; Duteanu, N.; Morgovan, C.M.; Jurca, T.; Pallag, A.; Marian, E. Sodium alginate—Natural microencapsulation material of polymeric microparticles. Int. J. Mol. Sci. 2022, 23, 12108. [Google Scholar] [CrossRef]
- Chen, M.J.; Chen, K.N. Applications of probiotic encapsulation in dairy products. In Encapsulation and Controlled Release Technologies in Food Systems; Lakkis, J.A., Ed.; Balckwell Publishing: Oxford, UK, 2007; pp. 83–112. [Google Scholar]
- Riseh, R.S.; Vazvani, M.G.; Hassanisaadi, M.; Thakur, V.K.; Kennedy, J.F. Use of whey protein as a natural polymer for the encapsulation of plant biocontrol bacteria: A review. Int. J. Biol. Macromol. 2023, 234, 123708. [Google Scholar]
- El-Salam, M.H.; El-Shibiny, S. Preparation and properties of milk proteins-based encapsulated probiotics: A review. Dairy Sci Technol. 2015, 95, 393–412. [Google Scholar] [CrossRef]
- Estevinho, B.N.; Rocha, F. Application of biopolymers in microencapsulation processes. In Biopolymers for Food Design; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: New York, NY, USA, 2018; pp. 191–222. [Google Scholar]
- Linares-Morales, J.R.; Gutiérrez-Méndez, N.; Rivera-Chavira, B.E.; Pérez-Vega, S.B.; Nevárez-Moorillón, G.V. Biocontrol processes in fruits and fresh produce, the use of lactic acid bacteria as a sustainable option. Front. Sustain. Food Syst. 2018, 2, 50. [Google Scholar] [CrossRef]
- Gasperini, L.; Mano, J.F.; Reis, R.L. Natural polymers for the microencapsulation of cells. J. R. Soc. Interface 2014, 11, 20140817. [Google Scholar] [CrossRef]
- Bianchini, A. Lactic acid bacteria as antifungal agents. In Advances in Fermented Foods and Beverages; Holzapfel, W., Ed.; Woodhead Publishing: Cambridge, UK, 2015; pp. 333–353. [Google Scholar]
- Dijksterhuis, J. Fungal spores: Highly variable and stress-resistant vehicles for distribution and spoilage. Food Microbiol. 2019, 81, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Sephton-Clark, P.C.; Voelz, K. Spore germination of pathogenic filamentous fungi. Adv. Appl. Microbiol. 2018, 102, 117–157. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Code | Strain | Formulations |
|---|---|---|
| ALG15 | L. plantarum TEP15 | Sodium alginate (2% w/v) |
| ALGWPC15 | L. plantarum TEP15 | Sodium alginate (0.75% w/v) + whey protein (1.5% w/v) |
| ALGCAR15 | L. plantarum TEP15 | Sodium alginate (2% w/v) + κ-carrageenan (0.05% w/v) |
| ALGGA15 | L. plantarum TEP15 | Sodium alginate (2% w/v) + gum arabic (1% w/v) |
| ALG4 | L. pentosus TEJ4 | Sodium alginate (2% w/v) |
| ALGWPC4 | L. pentosus TEJ4 | Sodium alginate (0.75% w/v) + whey protein (1.5% w/v) |
| ALGWPC4 | L. pentosus TEJ4 | Sodium alginate (2% w/v) + κ-carrageenan (0.05% w/v) |
| ALGGA4 | L. pentosus TEJ4 | Sodium alginate (2% w/v) + gum arabic (1% w/v) |
| Treatment | Day 0 | Day 14 | Day 28 | |||
|---|---|---|---|---|---|---|
| ALG15 | 94.57 ± 1.21 | ab B | 95.87 ± 1.50 | a AB | 97.24 ± 0.31 | a A |
| ALG4 | 93.71 ± 1.31 | b B | 94.45 ± 0.75 | b B | 95.96 ± 0.54 | d A |
| ALGGA15 | 95.31 ± 1.34 | ab A | 96.67 ± 0.34 | a A | 96.88 ± 0.18 | abc A |
| ALGGA4 | 95.77 ± 1.10 | a A | 96.11 ± 0.80 | a A | 96.93 ± 0.18 | ab A |
| ALGCAR15 | 94.80 ± 0.12 | ab C | 95.84 ± 0.66 | a B | 97.10 ± 0.18 | a A |
| ALGCAR4 | 95.21 ± 0.35 | ab B | 95.97 ± 0.13 | a A | 96.26 ± 0.25 | cd A |
| ALGWPC15 | 95.13 ± 0.20 | ab C | 95.80 ± 0.60 | ab B | 96.42 ± 0.20 | bcd A |
| ALGWPC4 | 94.59 ± 0.41 | ab B | 95.43 ± 0.45 | ab A | 95.81 ± 0.27 | d A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Ruiz, Y.; Mejía-Reyes, D.; Rosas-Quijano, R.; Gálvez-López, D.; Salvador-Figueroa, M.; Vázquez-Ovando, A. Antifungal Capacity of Microcapsules Containing Lactiplantibacillus plantarum TEP15 or Lactiplantibacillus pentosus TEJ4. Processes 2024, 12, 763. https://doi.org/10.3390/pr12040763
Pérez-Ruiz Y, Mejía-Reyes D, Rosas-Quijano R, Gálvez-López D, Salvador-Figueroa M, Vázquez-Ovando A. Antifungal Capacity of Microcapsules Containing Lactiplantibacillus plantarum TEP15 or Lactiplantibacillus pentosus TEJ4. Processes. 2024; 12(4):763. https://doi.org/10.3390/pr12040763
Chicago/Turabian StylePérez-Ruiz, Yeni, David Mejía-Reyes, Raymundo Rosas-Quijano, Didiana Gálvez-López, Miguel Salvador-Figueroa, and Alfredo Vázquez-Ovando. 2024. "Antifungal Capacity of Microcapsules Containing Lactiplantibacillus plantarum TEP15 or Lactiplantibacillus pentosus TEJ4" Processes 12, no. 4: 763. https://doi.org/10.3390/pr12040763