
Fructooligosaccharides: A Comprehensive Review on Their Microbial Source, Functional Benefits, Production Technology, and Market Prospects

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Functional Properties of Fructooligosaccharides
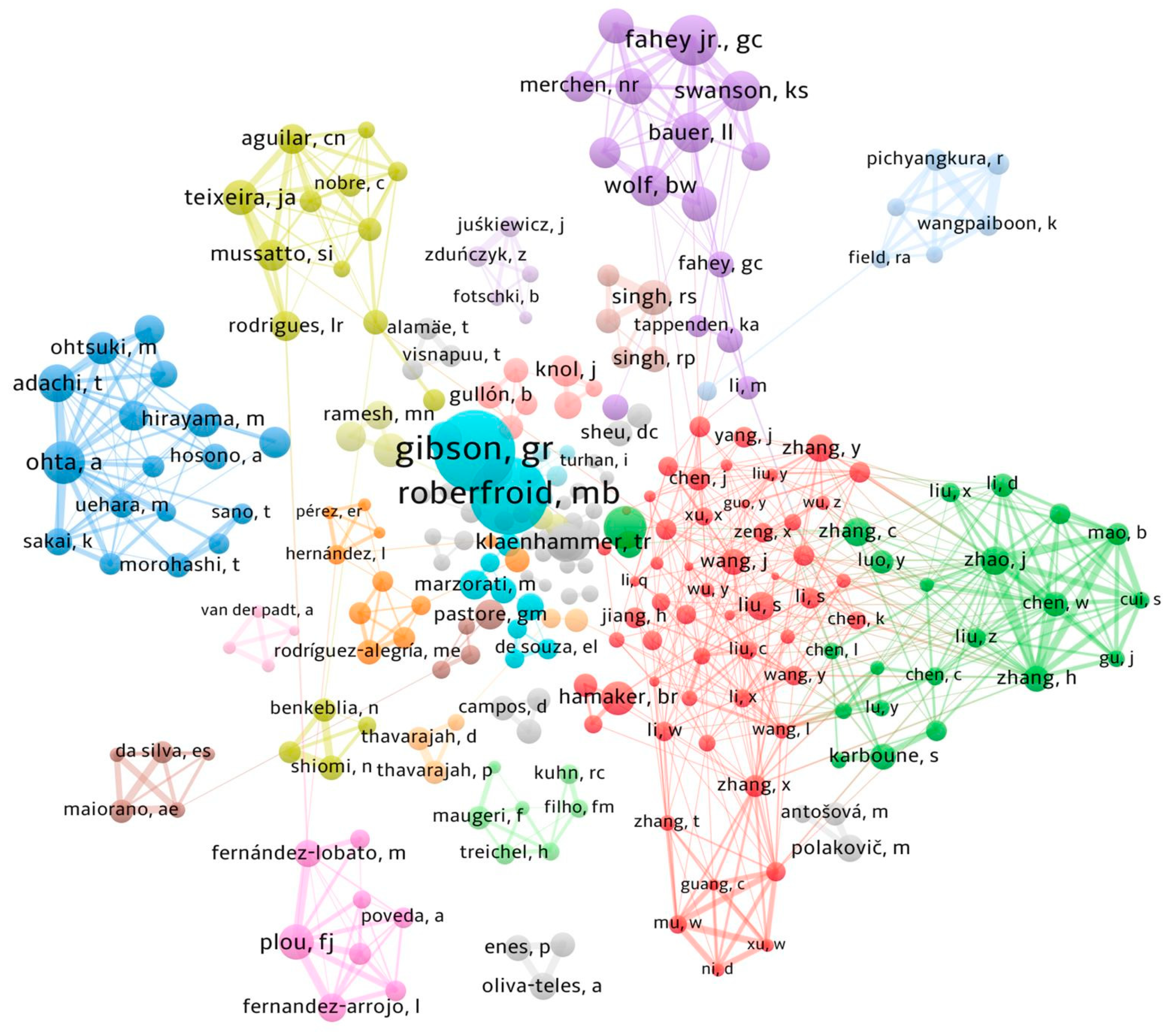
3. Bibliometric Analysis of FOS
4. Microorganisms Potentially Producing Enzymes with Fructosyltransferase Activity
5. Production of Fructooligosaccharides
5.1. Production of Fructooligosaccharides via Fermentation
5.1.1. Submerged Fermentation
5.1.2. Solid-State Fermentation
5.2. Production of Fructooligosaccharides Using Whole Cells and Soluble FTase Extracts
5.2.1. Soluble FTase
5.2.2. Whole Cells
6. Recovery and Purification of Fructooligosaccharides
6.1. Separation and Purification of Saccharides for FOS Production
6.1.1. FOS Purification by Adsorption
6.1.2. Membrane Technology and FOS Purification
7. Global Market and Techno-Economic Analysis of FOSs
7.1. FOS Global Market
7.2. Techno-Economic Analysis of FOS Production
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manicardi, T.; Baioni e Silva, G.; Longati, A.A.; Paiva, T.D.; Souza, J.P.M.; Pádua, T.F.; Furlan, F.F.; Giordano, R.L.C.; Giordano, R.C.; Milessi, T.S. Xylooligosaccharides: A bibliometric analysis and current advances of this bioactive food chemical as a potential product in biorefineries portfolios. Foods 2023, 12, 3007. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Xu, W.; Zhu, Y.; Pang, X.; Lv, J.; Mu, W. Insight into the effects and biotechnological production of kestoses, the smallest fructooligosaccharides. Crit. Rev. Biotechnol. 2020, 41, 34–46. [Google Scholar] [CrossRef]
- Bhadra, S.; Chettri, D.; Verma, A.K. Microbes in fructooligosaccharides production. Bioresour. Technol. Rep. 2022, 20, 101159. [Google Scholar] [CrossRef]
- Belmonte-Izquierdo, Y.; Salomé-Abarca, L.F.; González-Hernández, J.C.; López, M.G. Fructooligosaccharides (FOS) production by microorganisms with fructosyltransferase activity. Fermentation 2023, 9, 968. [Google Scholar] [CrossRef]
- Rahim, M.A.; Saeed, F.; Khalid, W.; Hussain, M.; Anjum, F.M. Functional and nutraceutical properties of fructo-oligosaccharides derivatives: A review. Int. J. Food Prop. 2021, 24, 1588–1602. [Google Scholar] [CrossRef]
- Witkowski, M.; Nemet, I.; Li, X.S.; Wilcox, J.; Ferrell, M.; Alamri, H.; Gupta, N.; Wang, Z.; Tang, W.H.W.; Hazen, S.L. Xylitol is prothrombotic and associated with cardiovascular risk. Eur. Heart J. 2024, 45, 2439–2452. [Google Scholar] [CrossRef]
- Rawat, H.K.; Nath, S.; Sharma, I.; Kango, N. Recent developments in the production of prebiotic fructooligosaccharides using fungal fructosyltransferases. Mycology 2024, 15, 564–584. [Google Scholar] [CrossRef]
- Sánchez-Martínez, M.J.; Soto-Jover, S.; Antolinos, V.; Martínez-Hernández, G.B.; López-Gómez, A. Manufacturing of short-chain fructooligosaccharides: From laboratory to industrial scale. Food Eng. Ver. 2020, 12, 149–172. [Google Scholar] [CrossRef]
- Pereira, R.S.; Vieira, A.C.; Leite, P.C.; Maestrelli, S.C.; Silva, E.S.; Maiorano, A.E.; Xavier, M.C.A.; Lopes, M.S.; De Paula, A.V.; Morales, S.A.V.; et al. Application of an agro-waste for the immobilization of microbial fructosyltransferase: A new alternative for fructooligosaccharide production. J. Braz. Chem. Soc. 2025, 36, e-20240172. [Google Scholar] [CrossRef]
- Guerra, L.; Romanini, D.; López, S.; Castelli, V.; Clementz, A. Upcycling of carrot discards into prebiotics (fructooligosaccharides) as high value food ingredients. Food Bioprod. Process. 2023, 138, 172–180. [Google Scholar] [CrossRef]
- Bis-Souza, C.V.; Pateiro, M.; Domínguez, R.; Penna, A.L.B.; Lorenzo, J.M.; Barretto, A.C.S. Impact of fructooligosaccharides and probiotic strains on the quality parameters of low-fat Spanish Salchichón. Meat. Sci. 2020, 159, 107936. [Google Scholar] [CrossRef]
- Silva, K.C.G.; Sato, A.C.K. Biopolymer gels containing fructooligosaccharides. Food Res. Int. 2017, 101, 88–95. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, O.; Flores-Gallegos, A.C.; Muñíz-Marquez, D.; Nobre, C.; Contreras-Esquivel, J.C.; Aguilar, C.N. Fructooligosaccharides production from agro-wastes as alternative low-cost source. Trends Food Sci. Technol. 2019, 91, 139–146. [Google Scholar] [CrossRef]
- Wang, S.; Pan, J.; Zhang, Z.; Yan, X. Investigation of dietary fructooligosaccharides from different production methods: Interpreting the impact of compositions on probiotic metabolism and growth. J. Funct. Foods 2020, 69, 103955. [Google Scholar] [CrossRef]
- Wong, W.-Y.; Chan, B.D.; Leung, T.-W.; Chen, M.; Tai, W.C.-S. Beneficial and anti-inflammatory effects of formulated prebiotics, probiotics, and symbiotic in normal and acute colitis mice. J. Funct. Foods 2022, 88, 104871. [Google Scholar] [CrossRef]
- Figueiredo, F.C.; Ranke, F.F.B.; Oliva-Neto, P. Evaluation of xylooligosaccharides and fructooligosaccharides on digestive enzymes hydrolysis and as a nutrient for different probiotics and Salmonella typhimurium. LWT 2020, 118, 108761. [Google Scholar] [CrossRef]
- Stojanović, S.; Ristović, M.; Stepanović, J.; Margetić, A.; Duduk, B.; Vujčić, Z.; Dojnov, B. Aspergillus welwitschiae inulinase enzyme cocktails obtained on agro-material inducers for the purpose of fructooligosaccharides production. Food Res. Int. 2022, 160, 111755. [Google Scholar] [CrossRef]
- Meng, Y.; Ma, Q.; Xu, X.; Feng, L.; Chen, Q.; Chen, Y.; Li, Z.; Liu, C.; Chen, K. Burdock fructooligosaccharide ameliorates the hypercholesterolemia and vascular inflammation in mice by regulating cholesterol homeostasis and anti-inflammatory properties. J. Funct. Foods 2023, 107, 105678. [Google Scholar] [CrossRef]
- Yuan, P.; Shao, T.; Han, J.; Liu, C.; Wang, G.; He, S.; Xu, S.; Nian, S.; Chen, K. Burdock fructooligosaccharide as an α-glucosidase inhibitor and its antidiabetic effect on high-fat diet and streptozotocin-induced diabetic mice. J. Funct. Foods 2021, 86, 104703. [Google Scholar] [CrossRef]
- Ojwach, J.; Kumar, A.; Mukaratirwa, S.; Mutanda, T. Fructooligosaccharides synthesized by fructosyltransferase from an indigenous coprophilous Aspergillus niger strain XOBP48 exhibits antioxidant activity. Bioact. Carbohydr. Diet. Fibre 2020, 24, 100238. [Google Scholar] [CrossRef]
- Bandyopadhyay, B.; Mitra, P.K.; Mandal, V.; Mandal, N.C. Novel fructooligosaccharides of Dioscorea alata L. tuber have prebiotic potentialities. Eur. Food Res. Technol. 2021, 247, 3099–3112. [Google Scholar] [CrossRef]
- Hajar-Azhari, S.; Rahim, M.H.A.; Razid Sarbini, S.; Muhialdin, B.J.; Olusegun, L.; Saari, N. Enzymatically synthesised fructooligosaccharides from sugarcane syrup modulate the composition and short-chain fatty acid production of the human intestinal microbiota. Food Res. Int. 2021, 149, 110677. [Google Scholar] [CrossRef]
- Kaewarsar, E.; Chaiyasut, C.; Lailerd, N.; Makhamrueang, N.; Peerajan, S.; Sirilun, S. Optimization of mixed inulin, fructooligosaccharides, and galactooligosaccharides as prebiotics for stimulation of probiotics growth and function. Foods 2023, 12, 1591. [Google Scholar] [CrossRef]
- Kariyawasam, K.M.G.M.M.; Lee, N.-K.; Paik, H.-D. Synbiotic yoghurt supplemented with novel probiotic Lactobacillus brevis KU200019 and fructooligosaccharides. Food Biosci. 2021, 39, 100835. [Google Scholar] [CrossRef]
- Dong, Q.; Lu, X.; Gao, B.; Liu, Y.; Aslam, M.Z.; Wang, X.; Li, Z. Lactiplantibacillus plantarum subsp. plantarum and fructooligosaccharides combination inhibits the growth, adhesion, invasion, and virulence of Listeria monocytogenes. Foods 2022, 11, 170. [Google Scholar] [CrossRef]
- Choi, G.-H.; Bock, H.-J.; Lee, N.-K.; Paik, H.-D. Soy Yogurt Using Lactobacillus plantarum 200655 and fructooligosaccharides: Neuroprotective effects against oxidative stress. J. Food Sci. Technol. 2022, 59, 4870–4879. [Google Scholar] [CrossRef]
- Toporovski, M.S.; De Morais, M.B.; Abuhab, A.; Crippa Júnior, M.A. Effect of polydextrose/fructooligosaccharide mixture on constipation symptoms in children aged 4 to 8 years. Nutrients 2021, 13, 1634. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Verma, N.K.; Thanabalu, T. Prebiotics in atopic dermatitis prevention and management. J. Funct. Food 2021, 78, 104352. [Google Scholar] [CrossRef]
- Wu, H.; Chiou, J. Potential benefits of probiotics and prebiotics for coronary heart disease and stroke. Nutrients 2021, 13, 2878. [Google Scholar] [CrossRef]
- Hidaka, H.; Eida, T.; Adachi, T.; Saitoh, Y. Industrial production of fructooligosaccharides and its application for human and animals. J. Agric. Chem. Soc. Jpn. 1987, 61, 915–923. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Ohtsuki, M.; Baba, S.; Adachi, T.; Sakata, T.; Sakaguchi, E. Calcium and magnesium absorption from the colon and rectum are increased in rats fed fructooligosaccharides. J. Nutr. 1995, 125, 2417–2424. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Sakai, K.; Takasaki, M.; Uehara, M.; Adlercreutz, H.; Morohashi, T.; Ishimi, Y.A. Combination of dietary fructooligosaccharides and isoflavone conjugates increases femoral bone mineral density and equol production in ovariectomized mice. J. Nutr. 2002, 132, 2048–2054. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, H.; Xue, S.; Ge, N.; Sun, Y.; Chi, Z.; Liu, G.; Chi, Z. Efficient conversion of cane molasses into fructooligosaccharides by a glucose derepression mutant of Aureobasidium melanogenum with high β-fructofuranosidase activity. J. Agric. Food Chem. 2019, 67, 13665–13672. [Google Scholar] [CrossRef]
- Ning, Y.; Wang, J.; Chen, J.; Yang, N.; Jin, Z.; Xu, X. Production of neo-fructooligosaccharides using free-whole-cell biotransformation by Xanthophyllomyces dendrorhous. Bioresour. Technol. 2010, 101, 7472–7478. [Google Scholar] [CrossRef]
- Gu, J.; Mao, B.; Cui, S.; Liu, X.; Zhang, H.; Zhao, J.; Chen, W. Metagenomic insights into the effects of fructooligosaccharides (FOS) on the composition of luminal and mucosal microbiota in C57BL/6J mice, especially the bifidobacterium composition. Nutrients 2019, 11, 2431. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Fahey, G.C., Jr.; Wolf, B.W. Selected indigestible oligosaccharides affect large bowel mass, cecal and fecal short-chain fatty acids, pH and microflora in rats. J. Nutr. 1997, 127, 130–136. [Google Scholar] [CrossRef]
- Li, M.; Monaco, M.H.; Wang, M.; Comstock, S.S.; Kuhlenschmidt, T.B.; Fahey Jr, G.C.; Miller, M.J.; Kuhlenschmidt, M.S.; Donovan, S.M. Human milk oligosaccharides shorten rotavirus-induced diarrhea and modulate piglet mucosal immunity and colonic microbiota. ISME J. 2014, 8, 1609–1620. [Google Scholar] [CrossRef]
- Nobre, C.; Simões, L.S.; Gonçalves, D.A.; Berni, P.; Teixeira, J.A. Fructooligosaccharides production and the health benefits of prebiotics. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 109–138. [Google Scholar] [CrossRef]
- Santos-Moriano, P.; Fernandez-Arrojo, L.; Mengibar, M.; Belmonte-Reche, E.; Peñalver, P.; Acosta, F.N.; Ballesteros, A.O.; Morales, J.C.; Kidibule, P.; Fernandez-Lobato, M.; et al. Enzymatic production of fully deacetylated chitooligosaccharides and their neuroprotective and anti-inflammatory properties. Biocatal. Biotransform. 2017, 36, 57–67. [Google Scholar] [CrossRef]
- Ghazi, I.; Fernandez-Arrojo, L.; Garcia-Arellano, H.; Ferrer, M.; Ballesteros, A.; Plou, F.J. Purification and kinetic characterization of a fructosyltransferase from Aspergillus aculeatus. J. Biotechnol. 2007, 128, 204–211. [Google Scholar] [CrossRef]
- Jayalakshmi, J.; Sadiq, M.A.; Sivakumar, V. Microbial enzymatic production of fructooligosaccharides from sucrose in agricultural harvest. Asian J. Microbiol. Biotechnol. Environ. Sci. 2021, 23, 84–88. [Google Scholar]
- Dhevagi, P.; Ramya, A.; Priyatharshini, S.; Geetha Thanuja, A.K.; Ambreetha, S.; Nivetha, A. Industrially Im-portant Fungal Enzymes: Productions and Applications. In Recent Trends in Mycological Research. Fungal Biology; Yadav, A.N., Ed.; Springer: Cham, Switzerland, 2021; pp. 263–309. [Google Scholar] [CrossRef]
- Faria, L.L.; Morales, S.A.V.; Prado, J.P.Z.; Dias, G.S.; De Almeida, A.F.; Xavier, M.D.C.A.; Silva, E.S.; Maiorano, A.E.; Perna, R.F. Biochemical characterization of extracellular fructosyltransferase from Aspergillus oryzae IPT-301 immobilized on silica gel for the production of fructooligosaccharides. Biotechnol. Lett. 2021, 43, 43–59. [Google Scholar] [CrossRef]
- Lammens, W.; Le Roy, K.; Schroeven, L.; Van Laere, A.; Rabijns, A.; Van den Ende, W. Structural insights into glycoside hy-drolase family 32 and 68 enzymes: Functional implications. J. Exp. Bot. 2009, 60, 727–740. [Google Scholar] [CrossRef]
- Almeida, M.N.; Guimarães, V.M.; Falkoski, D.L.; De Camargo, B.R.; Fontes-Sant’ana, G.C.; Maitan-Alfenas, G.P.; De Rezende, S.T. Purification and characterization of an invertase and a transfructosylase from Aspergillus terreus. J. Food Biochem. 2018, 42, e12551. [Google Scholar] [CrossRef]
- Hirabayashi, K.; Kondo, N.; Toyota, H.; Hayashi, S. Production of the functional trisaccharide 1-kestose from cane sugar molasses using Aspergillus japonicus β-fructofuranosidase. Curr. Microbiol. 2017, 74, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Ganaie, M.A.; Lateef, A.; Gupta, U.S. Enzymatic trends of fructooligosaccharides production by microorganisms. Appl. Biochem. Biotechnol. 2014, 172, 2143–2159. [Google Scholar] [CrossRef]
- Dias, G.S.; Santos, E.D.; Xavier, M.C.A.; Almeida, A.F.; Silva, E.S.; Maiorano, A.E.; Perna, R.F.; Morales, S.A.V. Study on the transfructosylation activity of Aspergillus oryzae IPT-301 cells in a packed bed reactor aiming at fructooligosaccharide production. Chem. Technol. Biotechnol. 2022, 97, 2904–2911. [Google Scholar] [CrossRef]
- Castro, C.C.; Nobre, C.; Duprez, M.E.; Weireld, G.; Hantson, A.L. Screening and selection of potential carriers to immobilize Aureobasidium pullulans cells for fructo-oligosaccharides production. Biochem. Eng. J. 2017, 118, 82–90. [Google Scholar] [CrossRef]
- Antošová, M.; Polakovič, M. Fructosyltransferases: The enzymes catalyzing the production of fructooligosaccharides. Chem. Pap. 2001, 55, 350–358. [Google Scholar]
- Fernandez, R.C.; Ottoni, C.A.; Da Silva, E.S.; Matsubara, R.M.; Carter, J.M.; Magossi, L.R.; Wada, M.A.; Rodrigues, M.F.A.; Maresma, B.G.; Maiorano, A.E. Screening of beta-fructofuranosidase-producing microorganisms and effect of pH and temperature on enzymatic rate. Appl. Microbiol. Biotechnol. 2007, 75, 87–93. [Google Scholar] [CrossRef]
- Michel, M.R.; Rodríguez-Jasso, R.M.; Aguilar, C.N.; Gonzalez-Herrera, S.M.; Flores-Gallegos, A.C.; Rodríguez-Herrera, R. Fructosyltransferase sources, production, and applications for prebiotics production. In Probiotics and Prebiotics in Human Nutrition and Health; IntechOpen: London, UK, 2016; p. 394. [Google Scholar] [CrossRef]
- Mohan, C.; Carvajal-Millan, E.; Ravishankar, C. Current trends in the biotechnological production of fructooligosaccharides. In Research Methodology in Food Sciences; Apple Academic Press: London, UK, 2018; pp. 199–220. [Google Scholar]
- Zhang, J.; Wang, L.; Luan, C.; Liu, G.; Liu, J.; Zhong, Y. Establishment of a rapid and effective plate chromogenic assay for screening of Aspergillus species with high -fructofuranosidase activity for fructooligosaccharides production. J. Microbiol. Methods. 2019, 166, 105740. [Google Scholar] [CrossRef]
- Aung, T.; Jiang, H.; Liu, G.; Chi, Z.; Hu, Z.; Chi, Z. Overproduction of β-fructofuranosidase1 with a high FOS synthesis activity for efficient biosynthesis of fructooligosaccharides. Int. J. Biol. Macromol. 2019, 30, 988–996. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, J.; Wang, L.; Liu, H.; Li, N.; Zhou, H.; Ning, Z.; Zhang, W.; Wang, L.; Huang, F.; et al. Continuous production of fructooligosaccharides by recycling of the thermal-stable β-fructofuranosidase produced by Aspergillus niger. Biotechnol. Lett. 2021, 43, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Choukade, R.; Kango, N. Characterization of a mycelial fructosyltransferase from Aspergillus tamarii NKRC 1229 for efficient synthesis of fructooligosaccharides. Food Chem. 2019, 286, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Batista, J.M.S.; Brandão-Costa, R.M.P.; Cunha, M.N.C.; Rodrigues, H.O.S.; Porto, A.L.F. Purification and biochemical characterization of an extracellular fructosyltransferase-rich extract produced by Aspergillus tamarii Kita UCP1279. Biocatal. Agric. Biotechnol. 2020, 26, 101647. [Google Scholar] [CrossRef]
- Tódero, L.M.; Rechia, C.G.V.; Guimarães, L.H.S. Production of short-chain fructooligosaccharides (scFOS) using extra-cellular β-D-fructofuranosidase produced by Aspergillus thermomutatus. J. Food Biochem. 2019, 43, e12937. [Google Scholar] [CrossRef]
- Maiorano, A.E.; Silva, E.S.; Perna, R.F.; Ottoni, C.A.; Piccoli, R.A.M.; Fernandez, R.C.; Maresma, B.G.; Rodrigues, M.F.A. Effect of agitation speed and aeration rate on fructosyltransferase production of Aspergillus oryzae IPT-301 in stirred tank bioreactor. Biotechnol. Lett. 2020, 42, 2619–2629. [Google Scholar] [CrossRef]
- Araújo, V.P.B.; Araújo, T.K.; Sousa, K.M.N.; Albuquerque, W.W.C.; Nascimento, A.K.C.D.; Cardoso, K.B.B.; Nascimento, T.P.; Batista, J.M.D.S.; Cavalcanti, M.T.H.; Porto, A.L.F.; et al. A novel β-fructofuranosidase produced by Penicillium citreonigrum URM 4459: Purification and biochemical features. Prep. Biochem. Biotechnol. 2023, 53, 906–913. [Google Scholar] [CrossRef]
- Xie, Y.; Zhou, H.; Liu, C.; Zhang, J.; Li, N.; Zhao, Z.; Sun, G.; Zhong, Y. A molasses habitat-derived fungus Aspergillus tubingensis XG21 with high β-fructofuranosidase activity and its potential use for fructooligosaccharides production. AMB Expr. 2017, 7, 128. [Google Scholar] [CrossRef]
- Han, S.; Pan, L.; Zeng, W.; Yang, L.; Yang, D.; Chen, G.; Liang, Z. Improved production of fructooligosaccharides (FOS) using a mutant strain of Aspergillus oryzae S719 overexpressing β-fructofuranosidase (FTase) genes. LWT 2021, 146, 111346. [Google Scholar] [CrossRef]
- Ojwach, J.; Kumar, A.; Mukaratirwa, S.; Mutanda, T. Purification and biochemical characterization of an extracellular fructo-syltransferase enzyme from Aspergillus niger sp. XOBP48: Implication in fructooligosaccharide production. 3 Biotech 2020, 10, 459. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.S.; Hassanpour, M.; Mussatto, S.I.; Harrison, M.D.; Speight, R.E.; O’Hara, I.M.; Zhang, Z. Transformation of sugarcane molasses into fructooligosaccharides with enhanced prebiotic activity using whole-cell biocatalysts from Aureobasidium pullulans FRR 5284 and an invertase-deficient Saccharomyces cerevisiae 1403-7A. Bioresour. Bioprocess. 2021, 8, 64. [Google Scholar] [CrossRef]
- de Andrades, D.; Arfelli, V.C.; Oriente, A.; Henn, C.; Pereira, V.A.A.C.; Simao, R.C.G.; Silva, J.L.C.; Kadowaki, M.K. Improved production of Î2-galactosidase and Î2-fructofuranosidase by fungi using alternative carbon sources. Sci. Res. Essays 2015, 10, 236–242. [Google Scholar] [CrossRef]
- Muñiz-Márquez, D.B.; Contreras, J.C.; Rodríguez, R.; Mussatto, S.I.; Teixeira, J.A.; Aguilar, C.N. Enhancement of fructosyltransferase and fructooligosaccharides production by A. oryzae DIA-MF in solid-state fermentation using aguamiel as culture médium. Bioresour. Technol. 2016, 213, 276–282. [Google Scholar] [CrossRef]
- Michel, M.R.; Gallegos, A.C.F.; Villarreal-Morales, S.L.; Aguilar-Zárate, P.; Aguilar, C.N.; Riutort, M.; Rodríguez-Herrera, R. Fructosyltransferase production by Aspergillus oryzae BM-DIA using solid-state fermentation and the properties of its nucleotide and protein sequences. Folia Microbiol. 2021, 66, 469–481. [Google Scholar] [CrossRef]
- de Oliveira, R.L.; da Silva, W.B.; Couto, K.S.; Porto, T.S. Sequential cultivation method for β-fructofuranosidase production from Aspergillus tamarii URM4634, evaluation of their biochemical and kinetic/thermodynamic characteristics, and application on sucrose hydrolysis. 3 Biotech 2024, 14, 186–197. [Google Scholar] [CrossRef]
- Moreira, L.A.; Oliveira, A.H.C.; Guimarães, L.H.S. Production and characterization of an extracellular Mn2+ activated β-D-fructofuranosidase from Aspergillus labruscus ITAL 28.255 with transfructosylating activity. J. App. Biol. Biotechnol. 2024, 12, 236–243. [Google Scholar] [CrossRef]
- Liang, X.; Li, C.; Cao, W.; Cao, W.; Shen, F.; Wan, Y. Fermentative production of fructo-oligosaccharides using Aureobasidium pullulans: Effect of dissolved oxygen concentration and fermentation mode. Molecules 2021, 26, 3867. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.L.; Bernardino, M.I.S.; Silva, T.B.S.; Converti, A.; Porto, C.S.; Porto, T.S. Extraction and purification of Aspergillus tamarii β-fructofuranosidase with transfructosylating activity using aqueous biphasic systems (PEG/phosphate) and magnetic field. Prep. Biochem. Biotechnol. 2021, 52, 478–486. [Google Scholar] [CrossRef]
- de Oliveira, R.L.; da Silva, M.F.; Converti, A.; Porto, T.S. Production of β-fructofuranosidase with transfructosylating activity by Aspergillus tamarii URM4634 solid-state fermentation on agroindustrial by-products. Int. J. Biol. Macromol. 2020, 144, 343–350. [Google Scholar] [CrossRef]
- Dinarvand, M.; Rezaee, M.; Foroughi, M. Optimizing culture conditions for production of intra and extracellular in-ulinase and invertase from Aspergillus niger ATCC 20611 by response surface methodology (RSM). Braz. J. Microbiol. 2017, 48, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, C.; Xie, Y.; Li, N.; Ning, Z.; Du, N.; Huang, X.; Zhong, Y. Enhancing fructooligosaccharides production by genetic improvement of the industrial fungus Aspergillus niger ATCC 20611. J. Biotechnol. 2017, 249, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ademakinwa, A.N.; Ayinla, Z.A.; Agboola, F.K. Strain improvement and statistical optimization as a combined strategy for improving fructosyltransferase production by Aureobasidium pullulans NAC8. J. Genet. Eng. Biotechnol. 2017, 15, 345–358. [Google Scholar] [CrossRef]
- Coetzee, G.; Smith, J.J.; Görgens, J.F. Influence of codon optimization, promoter, and strain selection on the heterologous production of a β-fructofuranosidase from Aspergillus fijiensis ATCC 20611 in Pichia pastoris. Folia Microbiol. 2022, 67, 339–350. [Google Scholar] [CrossRef]
- Guilarte, B.; Gutarra, C.B.; Cuervo-Fernández, R.; Silva, E.S.; Maiorano, A.E.; Rodrigues, M.F.A. Mutagenesis of Aspergillus oryzae IPT-301 to improve the production of β-fructofuranosidase. Braz. J. Microbiol. 2010, 41, 186–195. [Google Scholar] [CrossRef]
- Mao, S.; Liu, Y.; Yang, J.; Ma, X.; Zeng, F.; Zhang, Z.; Wang, S.; Han, H.; Qin, H.M.; Lu, F. Cloning, expression and characterization of a novel fructosyltransferase from Aspergillus niger and its application in the synthesis of fructooligosaccharides. RSC Adv. 2019, 9, 23856–23863. [Google Scholar] [CrossRef]
- Yang, H.L.; Wang, Y.; Zhang, L.; Shen, W. Heterologous expression and enzymatic characterization of fructosyl-transferase from Aspergillus niger in Pichia pastoris. New Biotechnol. 2016, 33, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Torres, F.A.G.; Moraes, L.M.P. Proteínas recombinantes produzidas em leveduras. Biotecnol. Ciênc. Desenvol. 2002, 29, 20–22. [Google Scholar]
- Amorim, C.; Silvério, S.C.; Rodrigues, L.R. One-step process for producing prebiotic arabino-xylooligosaccharides from Brewer’s spent grain employing Trichoderma species. Food Chem. 2019, 270, 86–94. [Google Scholar] [CrossRef]
- Mitchell, D.A.; Krieger, N. Looking Through a New Lens: Expressing the Ping Pong Bi Bi equation in terms of specificity constants. Biochem. Eng. J. 2022, 178, 108276. [Google Scholar] [CrossRef]
- Vega, R.; Zuniga-Hansen, M.E. A new mechanism and kinetic model for the enzymatic synthesis of short-chain fructooligosaccharides from sucrose. Biochem. Eng. J. 2014, 82, 158–165. [Google Scholar] [CrossRef]
- Flores-Maltos, D.E.; Mussatto, S.I.; Contreras-Esquivel, J.C.; Rodriguez-Herrera, R.; Teixeira, J.A.; Aguilar, C.N. Biotechnological production and application of fructooligosaccharides. Crit. Rev. Biotechnol. 2014, 36, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Kampen, W.H. Nutritional Requirements in Fermentation Processes. In Fermentation and Biochemical Engineering Handbook, 3rd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 37–57. [Google Scholar] [CrossRef]
- Nobre, C.; Gonçalves, D.A.; Teixeira, J.A.; Rodrigues, L.R. One-step co-culture fermentation strategy to produce high-content fructooligosaccharides. Carbohydr. Polym. 2018, 201, 31–38. [Google Scholar] [CrossRef]
- Perna, R.; Cunha, J.; Goncalves, M.; Basso, R.; Silva, E.S.; Maiorano, A.E. Microbial fructosyltransferase: Production by submerged fermentation and evaluation of pH and temperature effects on transfructosylation and hydrolytic enzymatic activities. Int. J. Eng. Res. Sci. 2018, 4, 43–50. [Google Scholar] [CrossRef]
- Pessoni, R.A.B.; Simões, K.; Braga, M.R.; Figueiredo-Ribeiro, R.D.C.L. Effects of substrate composition on growth and fructo-oligosaccharide production by Gliocladium virens. Dyn. Biochem. Process Biotech. Mol. Biol. 2009, 3, 96–101. [Google Scholar]
- Xu, D.B.; Madrid, C.P.; Röhr, M.; Kubicek, C.P. The influence of type and concentration of the carbon source on production of citric acid by Aspergillus niger. Appl. Microbiol. Biotechnol. 1989, 30, 553–558. [Google Scholar] [CrossRef]
- Okuyama, M.; Serizawa, R.; Tanuma, M.; Kikuchi, A.; Sadahiro, J.; Tagami, T.; Lang, W.; Kimura, A. Molecular insight into regioselectivity of transfructosylation catalyzed by GH68 levansucrase and β-fructofuranosidase. J. Biol. Chem. 2021, 296, 100398. [Google Scholar] [CrossRef]
- Ko, H.; Bae, J.H.; Sung, B.H.; Kim, M.J.; Park, H.J.; Sohn, J.H. Microbial production of medium chain fructooligosac-charides by recombinant yeast secreting bacterial inulosucrase. Enzyme Microb. Technol. 2019, 130, 109364. [Google Scholar] [CrossRef]
- Ademakinwa, A.N.; Ayinla, Z.A.; Omitogun, O.G.; Agboola, F.K. Preparation, characterization and optimization of cross-linked fructosyltransferase aggregates for the production of prebiotic fructooligosaccharides. BioTechnologia 2018, 99, 417–434. [Google Scholar] [CrossRef]
- Choukade, R.; Kango, N. Production, properties, and applications of fructosyltransferase: A current appraisal. Crit. Rev. Biotechnol. 2021, 41, 1178–1193. [Google Scholar] [CrossRef]
- Nobre, C.; Alves Filho, E.G.; Fernandes, F.A.N.; Brito, E.S.; Rodrigues, S.; Teixeira, J.A.; Rodrigues, L.R. Production of fructooligosaccharides by Aspergillus ibericus and their chemical characterization. LWT 2018, 89, 58–64. [Google Scholar] [CrossRef]
- Nobre, C.; Do Nascimento, A.K.C.; Silva, S.P.; Coelho, E.; Coimbra, M.A.; Cavalcanti, M.T.H.; Teixeira, J.A.; Porto, A.L.F. Process development for the production of prebiotic fructooligosaccharides by Penicillium citreonigrum. Bioresour. Technol. 2019, 282, 464–474. [Google Scholar] [CrossRef]
- Reis, F.S.; Ćirić, A.; Stojković, D.; Barros, L.; Ljaljević-Grbić, M.; Soković, M.; Ferreira, I.C.F.R. Effects of different culture conditions on biological potential and metabolites production in three Penicillium isolates. Drug Dev. Ind. Pharm. 2015, 41, 253–262. [Google Scholar] [CrossRef]
- Barros, R.G.C.; Pereira, U.C.; Andrade, J.K.S.; Barbosa, P.F.; Vasconcelos, S.V.; Nogueira, J.P.; Rajan, M.; Narain, N. Intracellular and extracellular enzyme patterns during biosynthesis of short chain fructooligosaccharides from Aspergillus spp. strains: Profile, biological structure correlation and chemometric characterizations. Biores. Technol. Rep. 2020, 11, 100546. [Google Scholar] [CrossRef]
- Martin, L.J.; Akhavan, B.; Bilek, M.M.M. Electric Fields control the orientation of peptides irreversibly immobilized on radical-functionalized surfaces. Nat. Commun. 2018, 9, 357. [Google Scholar] [CrossRef] [PubMed]
- Kronstad, J.W.; Caza, M. Shared and distinct mechanisms of iron acquisition by bacterial and fungal pathogens of humans. front. cell. Infect. Microbiol. 2013, 4, 80. [Google Scholar] [CrossRef]
- Stautz, J.; Hellmich, Y.; Fuss, M.F.; Silberberg, J.M.; Devlin, J.R.; Stockbridge, R.B.; Hänelt, I. Molecular mechanisms for bacterial potassium homeostasis. J. Mol. Biol. 2021, 433, 166968. [Google Scholar] [CrossRef]
- Udeh, O. Role of magnesium ions on yeast performance during very high gravity fermentation. J. Brewing Distilling 2013, 4, 19–45. [Google Scholar] [CrossRef]
- Nascimento, G.C.; Batista, R.D.; do Amaral Santos, C.C.A.; da Silva, E.M.; de Paula, F.C.; Mendes, D.B.; de Oliveira, D.P.; Almeida, A.F. β-fructofuranosidase and β-d-fructosyltransferase from new Aspergillus carbonarius PC-4 strain isolated from canned peach syrup: Effect of carbon and nitrogen sources on enzyme production. Sci. World J. 2019, 2019, 6956202. [Google Scholar] [CrossRef]
- Muñiz-Márquez, D.B.; Teixeira, J.A.; Mussatto, S.I.; Contreras-Esquivel, J.C.; Rodríguez-Herrera, R.; Aguilar, C.N. Fructo-oligosaccharides (FOS) production by fungal submerged culture using aguamiel as a low-cost by-product. LWT 2019, 102, 75–79. [Google Scholar] [CrossRef]
- Nobre, C.; Castro, C.C.; Hantson, A.L.; Teixeira, J.A.; De Weireld, G.; Rodrigues, L.R. Strategies for the production of high-content fructooligosaccharides through the removal of small saccharides by co-culture or successive fermen-tation with yeast. Carbohydr. Polym. 2016, 136, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Germec, M.; Turhan, I. Effect of pH control and aeration on inulinase production from sugarbeet molasses in a bench-scale bioreactor. Biomass Convers. Biorefin. 2023, 13, 4727–4739. [Google Scholar] [CrossRef]
- Palma, M.B.; Milagres, A.M.F.; Prata, A.M.R.; De Mancilha, I.M. Influence of aeration and agitation rate on the xylanase activity from Penicillium janthinellum. Process Biochem. 1996, 31, 141–145. [Google Scholar] [CrossRef]
- Singh, R.S.; Singh, T.; Pandey, A. Production of fungal endoinulinase in a stirred tank reactor and fructooligosaccharides preparation by crude endoinulinase. Bioresour. Technol. Rep. 2021, 15, 100743. [Google Scholar] [CrossRef]
- Alvarado-Obando, M.; Contreras, N.; León, D.; Botero, L.; Beltran, L.; Díaz, D.; Rodríguez-López, A.; Reyes, L.H.; Alméciga-Díaz, C.J.; Sánchez, O.F. Engineering a heterologously expressed fructosyltransferase from Aspergillus oryzae N74 in Komagataella phaffii (Pichia pastoris) for kestose production. New Biotechnol. 2022, 69, 18–27. [Google Scholar] [CrossRef]
- Maumela, P.; Rose, S.; Van Rensburg, E.; Chimphango, A.F.A.; Görgens, J.F. Bioprocess optimisation for high cell density endoinulinase production from recombinant Aspergillus niger. Appl. Biochem. Biotechnol. 2021, 193, 3271–3286. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.S.; Lee, J.H.; Kim, J.M.; Park, S.W.; Kim, S.W. Effects of morphology and rheology on neo-fructosyltransferase production by Penicillium citrinum. Biotechnol. Bioprocess Eng. 2006, 11, 100–104. [Google Scholar] [CrossRef]
- Ul Haq, I.; Nawaz, A.; Rehman, A. Optimization of inoculum volume, fermentation medium and aeration rate for the production of glucose oxidase by UV mutant strain of Aspergillus niger AN-14. Pak. J. Bot. 2015, 47, 329–332. [Google Scholar]
- Veiter, L.; Rajamanickam, V.; Herwig, C. The filamentous fungal pellet—Relationship between morphology and productivity. Appl. Microbiol. Biotechnol. 2018, 102, 2997–3006. [Google Scholar] [CrossRef]
- Hölker, U.; Höfer, M.; Lenz, J. Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Appl. Microbiol. Biotechnol. 2004, 64, 175–186. [Google Scholar] [CrossRef]
- Singh, R.S.; Chauhan, K.; Kaur, K.; Pandey, A. Statistical optimization of solid-state fermentation for the production of fungal inulinase from apple pomace. Bioresour. Technol. Rep. 2020, 9, 100364. [Google Scholar] [CrossRef]
- De La Rosa, O.; Múñiz-Márquez, D.B.; Contreras-Esquivel, J.C.; Wong-Paz, J.E.; Rodríguez-Herrera, R.; Aguilar, C.N. Im-proving the fructooligosaccharides production by solid-state fermentation. Biocatal. Agric. Biotechnol. 2020, 27, 101704. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Ballesteros, L.F.; Martins, S.; Maltos, D.A.F.; Aguilar, C.N.; Teixeira, J.A. Maximization of fructooligosaccharides and β-fructofuranosidase production by Aspergillus japonicus under solid-state fermentation conditions. Food Bioprocess Technol. 2013, 6, 2128–2134. [Google Scholar] [CrossRef]
- Singhania, R.R.; Sukumaran, R.K.; Patel, A.K.; Larroche, C.; Pandey, A. Advancement and comparative profiles in the production technologies using solid-state and submerged fermentation for microbial cellulases. Enzyme Microb. Technol. 2010, 46, 541–549. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef]
- Robledo, A.; Aguilera-Carbó, A.; Rodriguez, R.; Martinez, J.L.; Garza, Y.; Aguilar, C.N. Ellagic acid production by Aspergillus niger in solid state fermentation of pomegranate residues. J. Ind. Microbiol. Biotechnol. 2008, 35, 507–513. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Teixeira, J.A. Increase in the fructooligosaccharides yield and productivity by solid-state fermentation with Aspergillus japonicus using agro-industrial residues as support and nutrient source. Biochem. Eng. J. 2010, 53, 154–157. [Google Scholar] [CrossRef]
- Ojwach, J.; Adetunji, A.I.; Mutanda, T.; Mukaratirwa, S. Oligosaccharides production from coprophilous fungi: An emerging functional food with potential health-promoting properties. Biotechnol. Rep. 2022, 33, e00702. [Google Scholar] [CrossRef]
- Bhargav, S.; Panda, B.P.; Ali, M.; Javed, S. Solid-state fermentation: An overview. Chem. Biochem. Eng. Q. 2008, 22, 49–70. [Google Scholar]
- Ojwach, J.; Kumar, A.; Mutanda, T.; Mukaratirwa, S. Fructosyltransferase and inulinase production by indigenous co-prophilous fungi for the biocatalytic conversion of sucrose and inulin into oligosaccharides. Biocat. Agric. Biotechnol. 2020, 30, 101867. [Google Scholar] [CrossRef]
- Charoenwongpaiboon, T.; Punnatin, P.; Klaewkla, M.; Pramoj Na Ayutthaya, P.; Wangpaiboon, K.; Chunsrivirot, S.; Field, R.A.; Pichyangkura, R. Conserved calcium-binding residues at the Ca-I site involved in fructooligosaccharide synthesis by Lactobacillus reuteri 121 inulosucrase. ACS Omega 2020, 5, 28001–28011. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Zhang, S.; Huang, Z.; Xu, W.; Zhang, W.; Mu, W. Directionally modulating the product chain length of an inu-losucrase by semi-rational engineering for efficient production of 1-kestose. Enzyme Microb. Technol. 2022, 160, 110085. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Gu, S.; Teng, K.; Ren, J.; Liu, G.; Zhong, J. Characterization of a novel fructosyltransferase from Lactobacillus crispatus, InuCA, that attaches to the cell surface by electrostatic interaction. Appl. Environ. Microbiol. 2022, 88, e02399-21. [Google Scholar] [CrossRef]
- Cunha, J.S.; Ottoni, C.A.; Morales, S.A.V.; Silva, E.S.; Maiorano, A.E.; Perna, R.F. Synthesis and characterization of fructosyltransferase from Aspergillus oryzae IPT-301 for high fructooligosaccharides production. Braz. J. Chem. Eng. 2019, 36, 657–668. [Google Scholar] [CrossRef]
- Khatun, M.S.; Harrison, M.D.; Speight, R.E.; O’Hara, I.M.; Zhang, Z. Efficient production of fructo-oligosaccharides from sucrose and molasses by a novel Aureobasidium pullulan strain. Biochem. Eng. J. 2020, 163, 107747. [Google Scholar] [CrossRef]
- Han, S.; Ye, T.; Leng, S.; Pan, L.; Zeng, W.; Chen, G.; Liang, Z. Purification and biochemical characteristics of a novel fructosyltransferase with a high fos transfructosylation activity from Aspergillus oryzae S719. Protein Exp. Purif. 2020, 167, 105549. [Google Scholar] [CrossRef] [PubMed]
- Picazo, B.; Flores-Gallegos, A.C.; Ilina, A.; Rodríguez-Jasso, R.M.; Aguilar, C.N. Production of an enzymatic extract from Aspergillus oryzae DIA-MF to improve the fructooligosaccharides profile of aguamiel. Front. Nutr. 2019, 6, 15. [Google Scholar] [CrossRef]
- Virgen-Ortíz, J.J.; Ibarra-Junquera, V.; Escalante-Minakata, P.; Centeno-Leija, S.; Serrano-Posada, H.; De Jesús Ornelas-Paz, J.; Pérez-Martínez, J.D.; Osuna-Castro, J.A. Identification and functional characterization of a fructooligosacchari-des-forming enzyme from Aspergillus aculeatus. Appl. Biochem. Biotechnol. 2016, 179, 497–513. [Google Scholar] [CrossRef]
- De Pereira, J.C.; Giese, E.C.; Moretti, M.M.d.S.; Gomes, A.C. dos S.; Perrone, O.M.; Boscolo, M.; da Silva, R.; Gomes, E.; Martins, D.A.B. Effect of metal ions, chemical agents and organic compounds on lignocellulolytic enzymes activities. Enzyme Inhib. Act. 2017, 29, 139–164. [Google Scholar] [CrossRef]
- Wei, T.; Huang, S.; Zang, J.; Jia, C.; Mao, D. Cloning, expression and characterization of a novel fructosyltransferase from Aspergillus oryzae ZZ-01 for the synthesis of sucrose 6-acetate. Catalysts 2016, 6, 67. [Google Scholar] [CrossRef]
- Charoenwongpaiboon, T.; Klaewkla, M.; Chunsrivirot, S.; Wangpaiboon, K.; Pichyangkura, R.; Field, R.A.; Prousoontorn, M.H. Rational re-design of Lactobacillus reuteri 121 inulosucrase for product chain length control. RSC Adv. 2019, 9, 14957–14965. [Google Scholar] [CrossRef]
- Garcia, R.L.; Dias, G.S.; Morales, S.A.V.; Xavier, M.C.A.; Silva, E.S.; Maiorano, A.E.; Tardioli, P.W.; Perna, R.F. Glu-taraldehyde-crosslinked cells from Aspergillus oryzae IPT-301 for high transfructosylation activity: Optimization of the immobilization variables, characterization and operational stability. Braz. J. Chem. Eng. 2021, 38, 273–285. [Google Scholar] [CrossRef]
- Choukade, R.; Kango, N. Purification of Aspergillus tamarii mycelial fructosyltransferase (m-FTase), optimized fos production, and evaluation of its anticancer potential. J. Food Sci. 2022, 87, 3294–3306. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, J.; Carle, R.; Kammerer, D.R. Adsorption and ion exchange: Basic principles and their application in food processing. J. Agric. Food Chem. 2010, 59, 22–42. [Google Scholar] [CrossRef]
- Nobre, C.; Teixeira, J.A.; Rodrigues, L.R. New Trends and technological challenges in the industrial production and purification of fructo-oligosaccharides. Crit. Rev. Food Sci. Nutr. 2013, 55, 1444–1455. [Google Scholar] [CrossRef]
- Sundari, C.S.; Balasubramanian, D. Hydrophobic surfaces in saccharide chains. Prog. Biophys. Mol. Biol. 1997, 67, 183–216. [Google Scholar] [CrossRef]
- Kuhn, R.C.; Filho, F.M. Purification of fructooligosaccharides in an activated charcoal fixed bed column. New Biotechnol. 2010, 27, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, R.C.; Mazutti, M.A.; Albertini, L.B.; Filho, F.M. evaluation of fructooligosaccharides separation using a fixed-bed column packed with activated charcoal. New Biotechnol. 2014, 31, 237–241. [Google Scholar] [CrossRef]
- Nobre, C.; Teixeira, J.A.; Rodrigues, L.R. Fructo-oligosaccharides purification from a fermentative broth using an activated charcoal column. New Biotechnol. 2012, 29, 395–401. [Google Scholar] [CrossRef]
- Goulding, R.W. Liquid chromatography of sugars and related polyhydric alcohols on cation exchangers. J. Chromatogr. A 1975, 103, 229–239. [Google Scholar] [CrossRef]
- Gramblička, M.; Polakovič, M. Adsorption equilibria of glucose, fructose, sucrose, and fructooligosaccharides on cation exchange resins. J. Chem. Eng. Data 2007, 52, 345–350. [Google Scholar] [CrossRef]
- Angyal, S.J. Complexes of metal cations with carbohydrates in solution. Adv. Carbohydr. Chem. Biochem. 1989, 47, 1–43. [Google Scholar] [CrossRef]
- Nobre, C.; Suvarov, P.; De Weireld, G. Evaluation of commercial resins for fructo-oligosaccharide separation. New Biotechnol. 2014, 31, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Liu, S.; Chai, Y.; Wu, G.; Guan, N.; Li, L. Zeolites for separation: Fundamental and application. J. Energy Chem. 2022, 71, 288–303. [Google Scholar] [CrossRef]
- Sherman, J.P.; Chao, C.C. Carbohydrate separations using zeolite molecular sieves. Stud. Surf. Sci. Catal. 1986, 28, 1025–1032. [Google Scholar] [CrossRef]
- Kuhn, R.C.; Mazutti, M.A.; Filho, F.M. Kinetic and mass transfer effects for adsorption of glucose, fructose, sucrose and fructooligosaccharides into x zeolite. LWT Food Sci. Technol. 2012, 48, 127–133. [Google Scholar] [CrossRef]
- Kuhn, R.C.; Mazutti, M.A.; Filho, F.M. Separation and purification of fructooligosaccharides on a zeolite fixed-bed column. J. Sep. Sci. 2014, 37, 927–933. [Google Scholar] [CrossRef]
- Piazzi Fuhr, A.C.F.; Vieira, Y.; Kuhn, R.C.; Salau, N.P.G. Selective adsorption processes for fructooligosaccharides separation by activated carbon and zeolites through machine learning. Chem. Eng. Res. Des. 2023, 190, 379–394. [Google Scholar] [CrossRef]
- Strathmann, H. Membrane separation processes. J. Membr. Sci. 1981, 9, 121–189. [Google Scholar] [CrossRef]
- Verbeke, R.; Nulens, I.; Thijs, M.; Lenaerts, M.; Bastin, M.; Van Goethem, C.; Koeckelberghs, G.; Vankelecom, I.F.J. Solutes in solvent resistant and solvent tolerant nanofiltration: How molecular interactions impact membrane rejection. J. Membr. Sci. 2023, 677, 121595. [Google Scholar] [CrossRef]
- Pinelo, M.; Jonsson, G.; Meyer, A.S. Membrane technology for purification of enzymatically produced oligosaccharides: Molecular and operational features affecting performance. Sep. Purif. Technol. 2009, 70, 1–11. [Google Scholar] [CrossRef]
- Miyajima, K.; Machida, K.; Nakagaki, M. Hydrophobic indexes for various monosaccharides. Bull. Chem. Soc. Jpn. 1985, 58, 2595–2599. [Google Scholar] [CrossRef]
- Wen, J.; Chen, Y.; Yan, Q.; Jiang, L.; Chen, X.; Fan, Y. Experiment on and modeling of purification of fructooligosaccharides using ceramic nanofiltration membranes. Sep. Purif. Technol. 2023, 323, 124508. [Google Scholar] [CrossRef]
- Rizki, Z.; Janssen, A.E.M.; Claassen, G.D.H.; Boom, R.M.; Van Der Padt, A. Multi-criteria design of membrane cascades: Selection of configurations and process parameters. Sep. Purif. Technol. 2020, 237, 116349. [Google Scholar] [CrossRef]
- Cao, W.; Deng, T.; Cao, W.; Shen, F.; Wan, Y. From sucrose to fructo-oligosaccharides: Production and purification of fructo-oligosaccharides by an integrated enzymatic catalysis and membrane separation process. Sep. Purif. Technol. 2022, 288, 120678. [Google Scholar] [CrossRef]
- Wen, J.; Han, Q.; Qiu, M.; Jiang, L.; Chen, X.; Fan, Y. Membrane technologies for the separation and purification of functional oligosaccharides: A review. Sep. Purif. Technol. 2024, 346, 127463. [Google Scholar] [CrossRef]
- Fructooligosaccharides—Global Market Report. Research and Markets. Available online: https://Www.Researchandmarkets.Com/Reports/5989824/Fructooligosaccharides-Global-Market-Report (accessed on 1 October 2024).
- Credence Research. Fructooligosaccharides (FOR) Market. Available online: https://www.credenceresearch.com/report/fructooligosaccharides-fos-market (accessed on 1 October 2024).
- Fructo-Oligosaccharides (FOS) Market—Global Industry Analysis and Forecast (2024–2030) by Type, Application and Region. Available online: https://www.maximizemarketresearch.com/market-report/fructo-oligosaccharides-fos-market/123240/ (accessed on 1 October 2024).
- Consumer Shift Towards Natural FOS Supplements Boosting Market for Functional Foods. Available online: https://www.pristinemarketinsights.com/fructo-oligosaccharides-market-report (accessed on 1 October 2024).
- FOS Prebiotic Powder, 1 Kg, Packet. Available online: https://www.indiamart.com/proddetail/fos-prebiotic-powder-2853167683455.html?mTd=1 (accessed on 1 October 2024).
- BSTBIO 95% Food Grade Fructooligosaccharides Powder Sweetener FOS Dried Food Additives in Kosher Certified Bag. Available online: https://www.alibaba.com/product-detail/BSTBIO-95-Food-Grade-Fructooligosaccharides-Powder_10000021279783.html (accessed on 1 October 2024).
- Fructooligosaccharides from Chicory. Available online: https://www.sigmaaldrich.com/BR/en/product/sigma/f8052 (accessed on 1 October 2024).
- Saporepuro FOS Oligofructose Powder 250 g. Available online: https://www.amazon.com.be/-/en/Saporepuro-FOS-oligofructose-powder-250/dp/B0CNWC6LNM (accessed on 1 October 2024).
- Amanful, B.; Dogbe, E.S.; Bosman, C.E.; Görgens, J.F. Stochastic techno-economic analysis for the co-production of alternative sweeteners in sugarcane biorefineries. Food Bioprod. Process. 2024, 143, 9–20. [Google Scholar] [CrossRef]
- Dogbe, E.S.; Mandegari, M.; Görgens, J.F. Revitalizing the sugarcane industry by adding value to a-molasses in biorefineries. Biofuels Bioprod. Bioref. 2020, 14, 1089–1104. [Google Scholar] [CrossRef]
- Bedzo, O.K.K.; Mandegari, M.; Görgens, J.F. Comparison of immobilized and free enzyme systems in industrial production of short-chain fructooligosaccharides from sucrose using a techno-economic approach. Biofuels Bioprod. Bioref. 2019, 13, 1274–1288. [Google Scholar] [CrossRef]
- Fructooligosaccharide, 1 kg. Available online: https://www.carlroth.com/pl/en/oligosaccharids/fructooligosaccharide/p/31ye.4? (accessed on 27 March 2025).
- Fructooligosaccharides Price. Available online: https://www.made-in-china.com/products-search/hot-china-products/Fructooligosaccharides_Price.html (accessed on 27 March 2025).
- Li, Y.; Pandelaere, M. The purity premium effect: The asymmetrical value change around pure products. Psychol. Mark. 2024, 41, 328–343. [Google Scholar] [CrossRef]
- Ordóñez, L.D. The effect of correlation between price and quality on consumer choice. Organ. Behav. Hum. Decis. Process 1998, 75, 258–273. [Google Scholar] [CrossRef] [PubMed]
- Gruska, R.M.; Baryga, A.; Kunicka-Styczyńska, A.; Brzeziński, S.; Rosicka-Kaczmarek, J.; Miśkiewicz, K.; Sumińska, T. Fresh and stored sugar beet roots as a source of various types of mono- and oligosaccharides. Molecules 2022, 27, 5125. [Google Scholar] [CrossRef] [PubMed]
- Ureta, M.M.; Romano, N.; Kakisu, E.; Gómez-Zavaglia, A. Synthesis of fructo-oligosaccharides using grape must and sucrose as raw materials. Food Res. Int. 2019, 123, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Klaver, M.; Petersen, A.M.; Görgens, J.F. Economic comparison of decentralized versus centralized processing of sugarcane to fructooligosaccharides and ethanol. Biofuels Bioprod. Bioref. 2023, 17, 1566–1578. [Google Scholar] [CrossRef]
- Vacharanukrauh, T.; Plubwungklam, S.; Pichyangkura, R.; Soottitantawat, A. Techno-economic comparison of different reactor types used in the manufacture of fructooligosaccharides from sucrose. Asia-Pac. J. Chem. Eng. 2023, 18, e2936. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Microorganism | Carbon Source | Enzymatic Activity | Yield/Conversion | References |
|---|---|---|---|---|
| Aureobasidium melanogenum (mutant D28) | Sugarcane molasses | 2100.0 ± 61 U mL−1 | 0.58 g FOS g−1 molasses | [55] |
| Aureobasidium melanogenum 33 | Sucrose | 577.7 U mL−1 | 0.66 g FOSg−1 sucrose | [56] |
| Aspergillus niger ATCC 20611 (FV1-11CCFV1-7) | Sucrose | 413.0 U g−1 | [57] | |
| 410.0 U g−1 | ||||
| Aspergillus niger ATCC 20611 | Sucrose | 310.0 U g−1 | ||
| Aspergillus tamarii NKRC 1229 | Sucrose | 21.58 U g−1 intra 14.45 U g−1 mycelial | 55.0% | [58] |
| Aspergillus tamarii Kita UCP 1229 | Sucrose | 1629.03 U gds−1 | [59] | |
| Aspergillus thermomutatus | Sucrose | 6.5 U mg−1 protein—extra 0.5 U mg−1 protein—intra | 62.5% | [60] |
| Aspergillus oryzae IPT 301 | Sucrose | 2100.0 U g−1 | [61] | |
| Penicillium citreonigrum URM 4459 | Sucrose | 22.05 U mg−1 370.67 µmol min−1 | [62] | |
| Aspergillus tubingensis XG21 | Sugarcane molasses | 558.3 U g−1 | 56.9% | [63] |
| Aspergillus oryzae S719 | Wheat bran | 155.4 U mL−1 | 586.0 ± 4.7 FOS L−1 | [64] |
| Aspergillus niger sp. XOBP48 | Sucrose | 1.219.17 U mg−1 protein | [65] | |
| Aureobasidium pullulans FRR 5284 | Sugarcane molasses | 23.6 U mL−1 (intra and extra) | [66] | |
| Chrysonilia sitophila | Soybean meal Quinoa meal | 3.98 U mL−1 (extra) 27.91 U mL−1 (intra) | [67] | |
| Gliocladium virens | Orange peel Quinoa meal | 0.32 U mL−1 (extra) 2.06 U mL−1 (intra) | [67] | |
| Aspergillus oryzae DIA-MF | Aguamiel | 1.347 U mL−1 | 9.06 g L−1 | [68] |
| Aspergillus niger NRRL3 | Carrot juice Carrot bagasse | 90.82 U mL−1 244 ± 5.00 U mgprot | [10] | |
| Penicillium citrinum | Banana peel Cane molasses | 6.9 ± 0.15 U mL−1 min−1 7.3 ± 0.29 U mL−1 min−1 | [42] | |
| Aspergillus oryzae DIA-MF Aspergillus flavus | Aguamiel | 1.59 U mL−1 0.27 U mL−1 | [69] | |
| Aspergillus welwitschiae FAW1 | Artichoke/wheat bran/peptone | 6.3 U mL−1 | [17] | |
| Aspergillus tamarii URM4634 | Soybean bran and sucrose | 26.10 U mL−1 | [70] | |
| Aspergillus labruscus ITAL 28.255 | Rye flour and sucrose | 1.81 U mL−1 (extra) 1.76 U mL−1 (intra) | 150 mg mL−1 | [71] |
| Aureobasidium pullulans ipe-3 KY618121 | Sucrose | 269.6 U/(g/L biomass) | 123.2 g L−1 | [72] |
| Aspergillus tamarii URM4634 | Sucrose | 50.90 U mL−1 | [73] | |
| A. tamarii URM4634 | Soy bran | 66.93 U mL−1 | [74] |
| Value | Observation/Information | Reference |
|---|---|---|
| USD 3.0/kg to USD 4.0/kg | Standard Purity FOS (50–70% Purity) Applications: Used in general food and beverage products as fiber or low-calorie sweetener where ultrapure is not critical. Characteristics: Contains a blend of FOS with other sugars such as glucose and sucrose. | [165] |
| USD 4.5/kg to USD 6.0/kg | High Purity FOS (70–90% Purity) Applications: Dietary supplements, functional foods, and products requiring higher concentrations of prebiotic activity. Characteristics: Enhanced prebiotic benefits due to reduced levels of monosaccharides and disaccharides. | [165] |
| USD 6.5/kg to USD 8.0/kg | Ultra-High Purity FOS (Above 90% Purity) Applications: Essential for infant formulas, pharmaceutical products, and specialty nutraceuticals where maximum efficacy and minimum contamination are critical. Characteristics: High concentration of FOS with minimal (or no) presence of other sugars, ensuring optimal prebiotic functionality. | [165] |
| USD 10.90/kg | Brand Tiens, Packet size of 1 kg. | [166] |
| USD 15/kg | Sample price of the supplier Kap (xiamen) Bio-Tech Co., Ltd. | [167] |
| USD 44/kg | FOS powder, brand SaporePuro, packet size of 250 g. | [168] |
| USD 2.61/kg | Selling price considered for FOS powder, 90% purity. | [169] |
| USD 3.20/kg | Selling price considered for FOS powder, 95% purity. | [170] |
| USD 0.47/kg | Selling price considered for 75% Brix FOS syrup. | [170] |
| USD 5.00/kg | Selling price considered for FOS powder, 95% purity. | [171] |
| USD 5.00/kg | Selling price considered for FOS powder, 97% purity. | [172] |
| USD 1057.00/kg a | Selling price considered for FOS ≥ 97% purity. | [173] |
| Marketplace value | There are several suppliers, each offering a different specified product. | [174] |
| USD 3.50/kg to USD 30.00/kg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, G.S.; Vieira, A.C.; Baioni e Silva, G.; Simões, N.F.; Milessi, T.S.; Saraiva, L.S.; Xavier, M.d.C.A.; Longati, A.A.; Rodrigues, M.F.A.; Fernandes, S.; et al. Fructooligosaccharides: A Comprehensive Review on Their Microbial Source, Functional Benefits, Production Technology, and Market Prospects. Processes 2025, 13, 1252. https://doi.org/10.3390/pr13041252
Dias GS, Vieira AC, Baioni e Silva G, Simões NF, Milessi TS, Saraiva LS, Xavier MdCA, Longati AA, Rodrigues MFA, Fernandes S, et al. Fructooligosaccharides: A Comprehensive Review on Their Microbial Source, Functional Benefits, Production Technology, and Market Prospects. Processes. 2025; 13(4):1252. https://doi.org/10.3390/pr13041252
Chicago/Turabian StyleDias, Giancarlo Souza, Ana Carolina Vieira, Gabriel Baioni e Silva, Nicole Favero Simões, Thais S. Milessi, Larissa Santos Saraiva, Michelle da Cunha Abreu Xavier, Andreza Aparecida Longati, Maria Filomena Andrade Rodrigues, Sergio Fernandes, and et al. 2025. "Fructooligosaccharides: A Comprehensive Review on Their Microbial Source, Functional Benefits, Production Technology, and Market Prospects" Processes 13, no. 4: 1252. https://doi.org/10.3390/pr13041252
APA StyleDias, G. S., Vieira, A. C., Baioni e Silva, G., Simões, N. F., Milessi, T. S., Saraiva, L. S., Xavier, M. d. C. A., Longati, A. A., Rodrigues, M. F. A., Fernandes, S., Silva, E. S. d., Maiorano, A. E., Morales, S. A. V., Basso, R. C., & Perna, R. F. (2025). Fructooligosaccharides: A Comprehensive Review on Their Microbial Source, Functional Benefits, Production Technology, and Market Prospects. Processes, 13(4), 1252. https://doi.org/10.3390/pr13041252