
Microbial Communities, Metabolites, Fermentation Quality and Aerobic Stability of Whole-Plant Corn Silage Collected from Family Farms in Desert Steppe of North China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
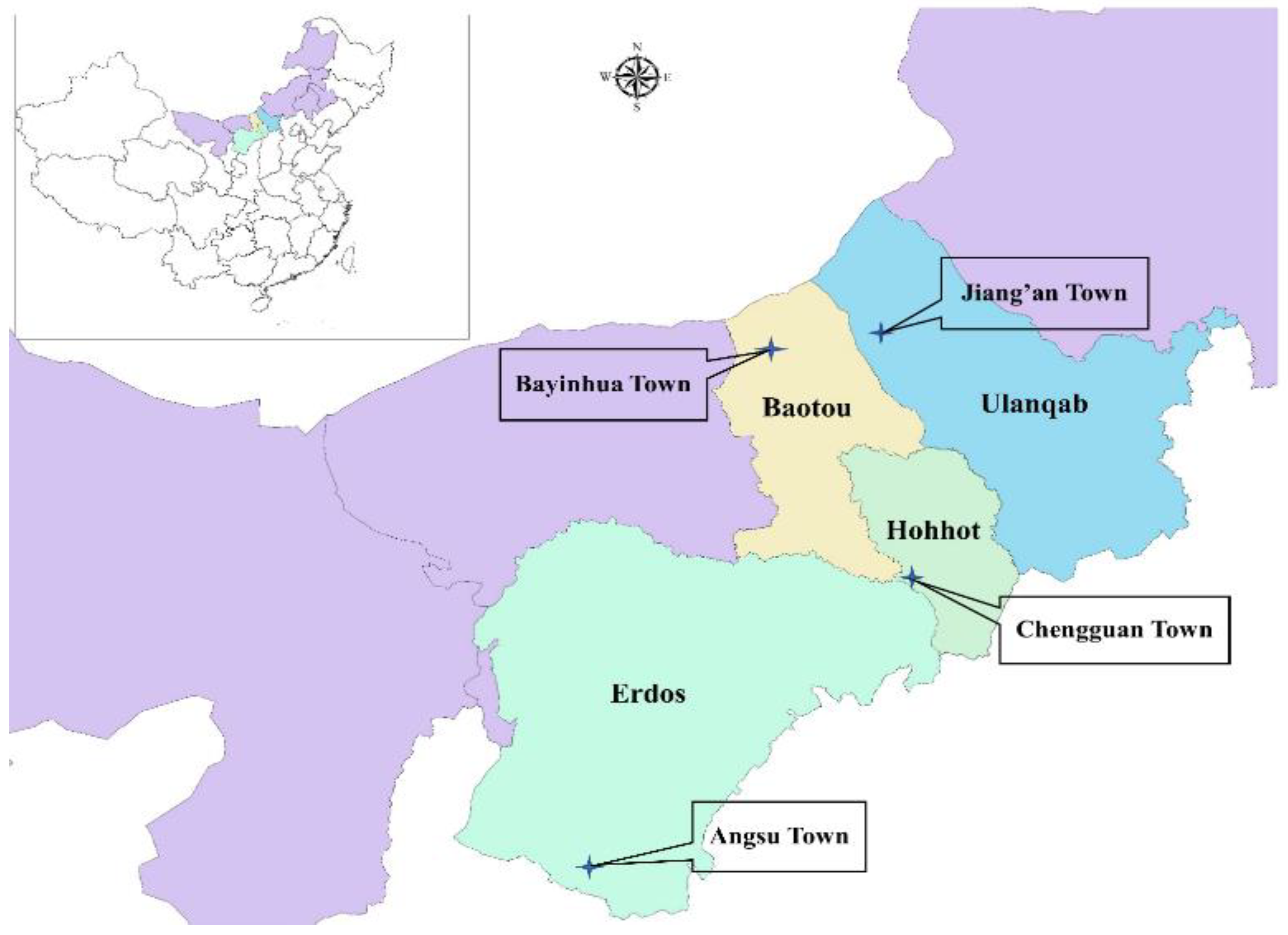
2.1. Sampling
2.2. Analyses
2.3. Statistical Analyses
3. Results
3.1. Characteristics of Whole-Plant Corns before Ensiling
3.2. Microbial Counts and Diversity
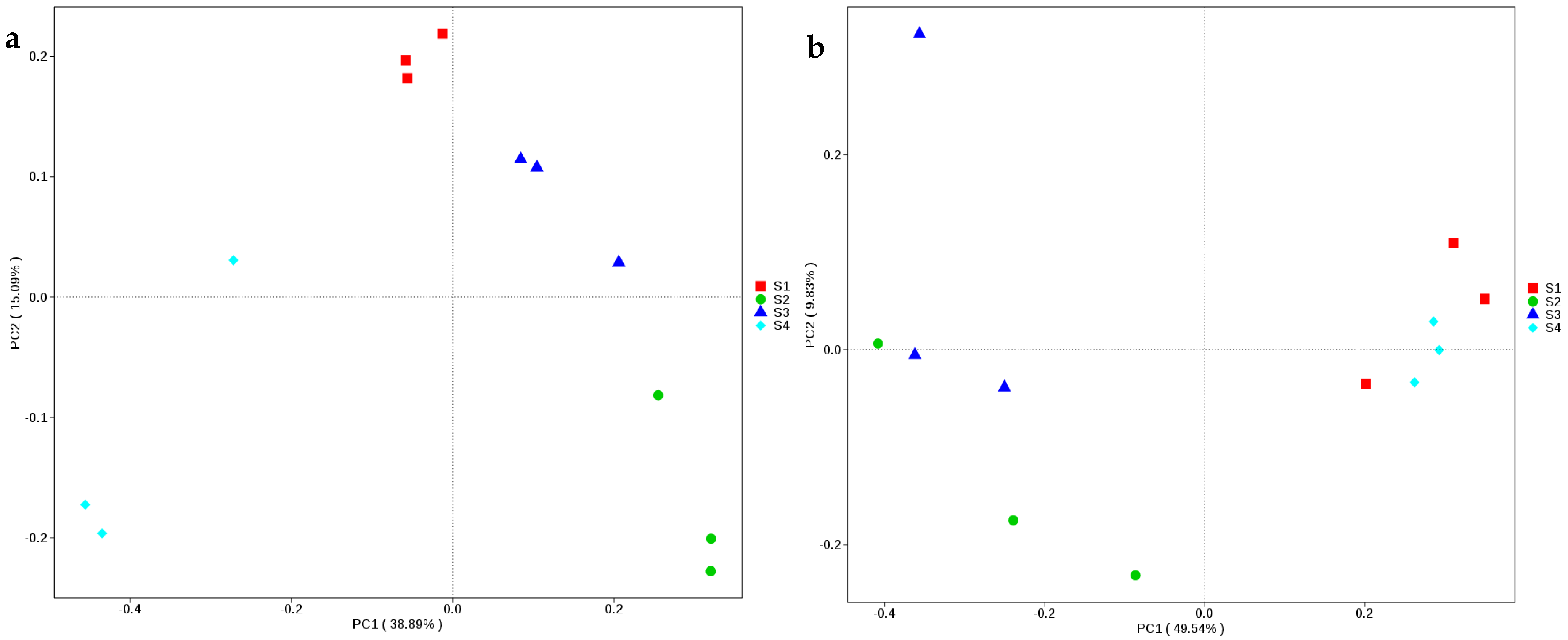
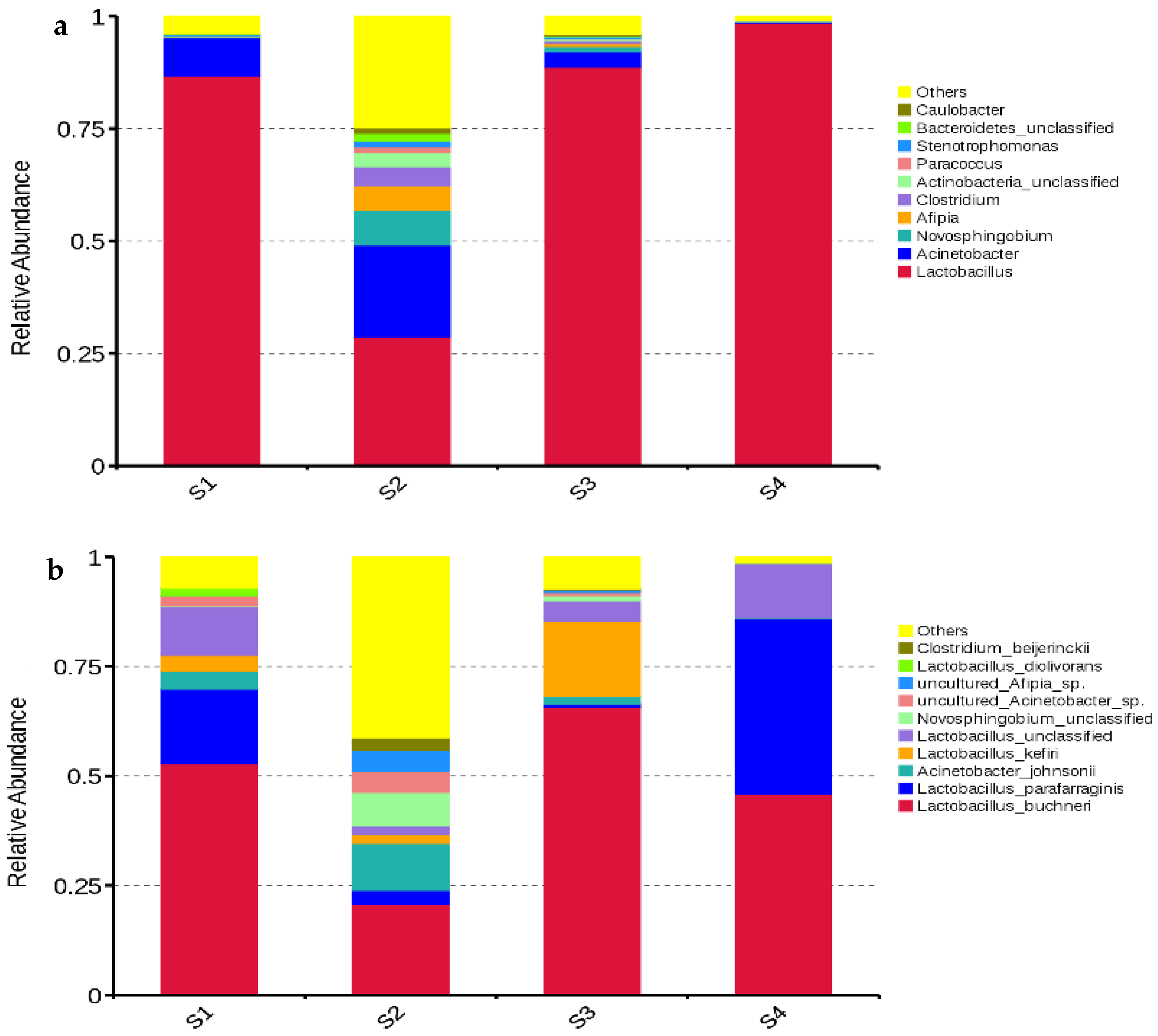
3.3. Bacterial Community
3.4. Fungal Community
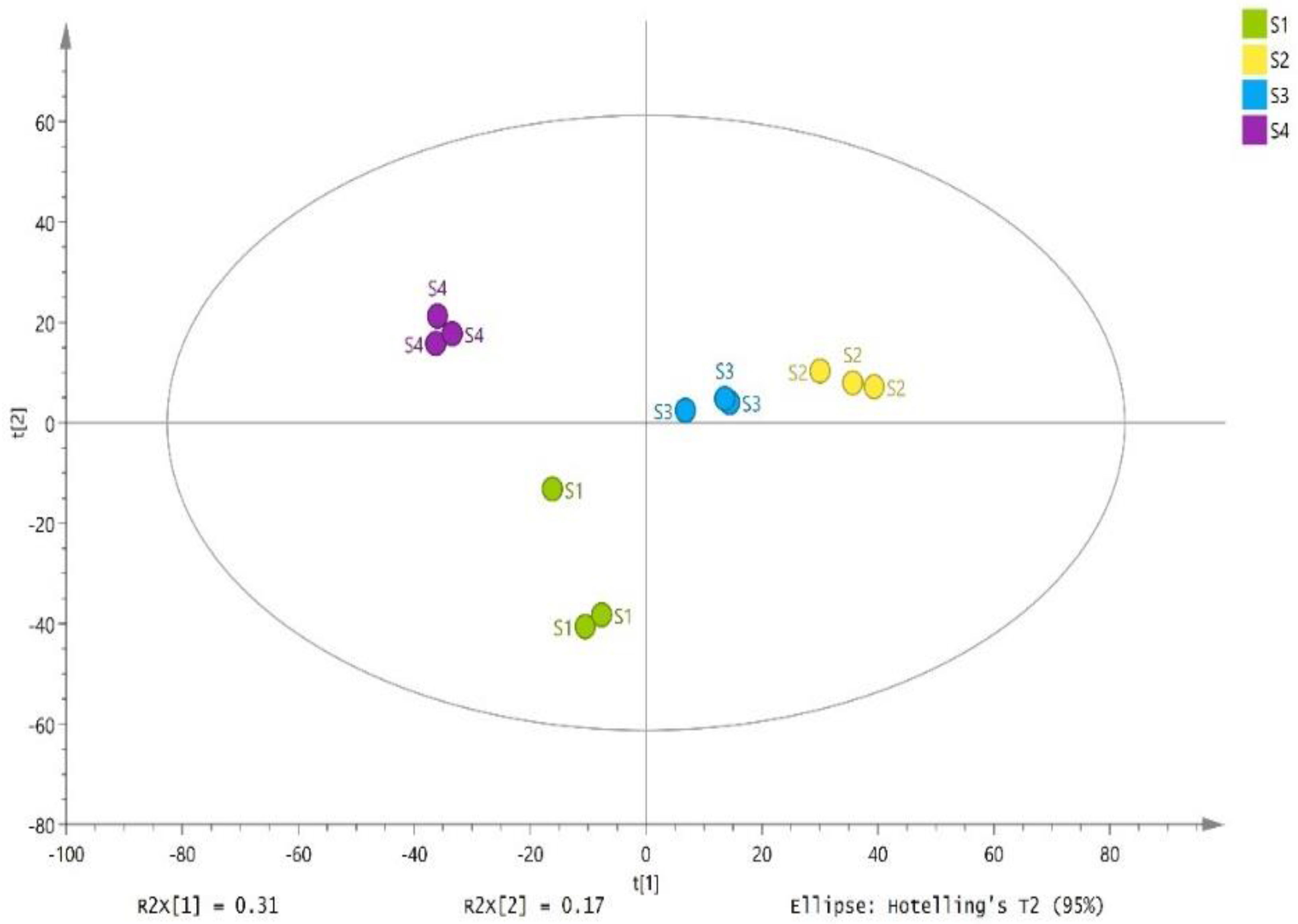
3.5. Metabolites
3.6. Fermentation Quality and Chemistry Composition
3.7. Aerobic Stability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gong, T.; Battese, G.E.; Villano, R.A. Family farms plus cooperatives in China: Technical efficiency in crop production. J. Asian Econ. 2019, 64, 101129. [Google Scholar] [CrossRef]
- Sachurina. The change of the pastoral management mode in Inner Mongolia: Joint households, cooperatives, household pastures and corporations. J. Arid Land Resour. Environ. 2017, 31, 56–63. [Google Scholar] [CrossRef]
- Wang, R.Z. Rational Forage and Livestock Allocation of Household Ranch in Inner Mongolia Pastoral Area. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2017. [Google Scholar]
- Romero, J.J.; Joo, Y.; Park, J.; Tiezzi, F.; Gutierrez-Rodriguez, E.; Castillo, M.S. Bacterial and fungal communities, fermentation, and aerobic stability of conventional hybrids and brown midrib hybrids ensiled at low moisture with or without a homo- and heterofermentative inoculant. J. Dairy Sci. 2018, 101, 3057–3076. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Bai, C.; Xu, H.; Na, N.; Jiang, Y.; Yin, G.; Liu, S.; Xue, Y. Succession of bacterial community during the initial aerobic, intense fermentation, and stable phases of whole-plant corn silages treated with lactic acid bacteria suspensions prepared from other silages. Front. Microbiol. 2021, 12, 655095. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wang, N.; Rinne, M.; Ke, W.; Weinberg, Z.G.; Da, M.; Bai, J.; Zhang, Y.; Li, F.; Guo, X. The bacterial community and metabolome dynamics and their interactions modulate fermentation process of whole crop corn silage prepared with or without inoculants. Microb. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Xu, D.; Ding, W.; Ke, W.; Li, F.; Zhang, P.; Guo, X. Modulation of metabolome and bacterial community in whole crop corn silage by inoculating homofermentative Lactobacillus plantarum and heterofermentative Lactobacillus buchneri. Front. Microbiol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Gharechahi, J.; Kharazian, Z.A.; Sarikhan, S.; Jouzani, G.S.; Aghdasi, M.; Salekdeh, G.H. The dynamics of the bacterial communities developed in maize silage. Microb. Biotechnol. 2017, 10, 1663–1676. [Google Scholar] [CrossRef]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]
- Drouin, P.; Tremblay, J.; Renaud, J.; Apper, E. Microbiota succession during aerobic stability of maize silage inoculated with Lentilactobacillus buchneri NCIMB 40788 and Lentilactobacillus hilgardii CNCM-I-4785. MicrobiologyOpen 2020, 10, e1153. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Muck, R.E. Ensiling in 2050: Some challenges and opportunities. Grass Forage sci. 2019, 74, 178–187. [Google Scholar] [CrossRef]
- Xu, D.; Ding, Z.; Wang, M.; Bai, J.; Ke, W.; Zhang, Y.; Guo, X. Characterization of the microbial community, metabolome and biotransformation of phenolic compounds of sainfoin (Onobrychis viciifolia) silage ensiled with or without inoculation of Lactobacillus plantarum. Bioresour. Technol. 2020, 316, 123910. [Google Scholar] [CrossRef]
- Wu, Z.; Luo, Y.; Bao, J.; Luo, Y.; Yu, Z. Additives affect the distribution of metabolic profile, microbial communities and antibiotic resistance genes in high-moisture sweet corn kernel silage. Bioresour. Technol. 2020, 315, 123821. [Google Scholar] [CrossRef] [PubMed]
- Steidlová, Š.; Kalac, P. Levels of biogenic amines in maize silages. Anim. Feed Sci. Technol. 2002, 102, 197–205. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123600. [Google Scholar] [CrossRef]
- Owens, V.N.; Albrecht, K.A.; Muck, R. EProtein degradation and ensiling characteristics of red clover and alfalfa wilted under varying levels of shade. Can. J. Plant Sci. 1999, 79, 209–222. [Google Scholar] [CrossRef]
- Cai, Y. Identification and characterization of Enterococcus species isolated from forage crops and their influence on silage fermentation. J. Dairy Sci. 1999, 82, 2466–2471. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, X.J.; Zhao, M.M.; Yu, Z. Isolating and evaluating lactic acid bacteria strains for effectiveness of Leymus chinensis silage fermentation. Lett. Appl. Microbiol. 2014, 59, 391–397. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3594. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.; Liu, X.; Chu, M.; Shi, J.; Zhang, H.; Liu, Y. A comparative UHPLCQqQ-MS-based metabolomics approach for evaluating Chinese and North American wild rice. Food Chem. 2019, 275, 618–627. [Google Scholar] [CrossRef] [PubMed]
- McEniry, J.; O’Kiely, P.; Clipson, N.; Forristal, P.; Doyle, E. Bacterial community dynamics during the ensilage of wilted grass. J. Appl. Microbiol. 2008, 105, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, X.; Gu, Q.; Liang, M.; Mu, S.; Zhou, B.; Huang, F.; Lin, B.; Zou, C. Analysis of the correlation between bacteria and fungi in sugarcane tops silage prior to and after aerobic exposure. Bioresour. Technol. 2019, 291, 121835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Meng, Q.; Zhou, Z.; Wu, H. A mixture of potassium sorbate and sodium benzoate improved fermentation quality of whole-plant corn silage by shifting bacterial communities. J. Appl. Microbiol. 2020, 128, 1312–1323. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Nazar, M.; Shao, T. Microbial diversity and fermentation profile of red clover silage inoculated with reconstituted indigenous and exogenous epiphytic microbiota. Bioresour. Technol. 2020, 123606. [Google Scholar] [CrossRef] [PubMed]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Guo, X.S.; Ke, W.C.; Ding, W.R.; Ding, L.M.; Xu, D.M.; Wang, W.W.; Zhang, P.; Yang, F.Y. Profiling of metabolome and bacterial community dynamics in ensiled Medicago sativa inoculated without or with Lactobacillus plantarum or Lactobacillus buchneri. Sci. Rep. 2018, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Xie, J. Assessment of metabolic changes in Acinetobacter johnsonii and Pseudomonas fluorescens co-culture from bigeye tuna (Thunnus obesus) spoilage by ultrahigh-performance liquid chromatography-tandem mass spectrometry. LWT Food Sci. Technol. 2020, 123, 109073. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biot. 2018, 102, 4025–4037. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.J.; Zhao, Y.; Balseca-Paredes, M.A.; Tiezzi, F.; Gutierrez-Rodriguez, E.; Castillo, M.S. Laboratory silo type and inoculation effects on nutritional composition, fermentation, and bacterial and fungal communities of oat silage. J. Dairy Sci. 2017, 100, 1812–1828. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Nishino, N. Effects of inoculation of Lactobacillus rhamnosus and Lactobacillus buchneri on fermentation, aerobic stability and microbial communities in whole crop corn silage. Grassl. Sci. 2011, 57, 184–191. [Google Scholar] [CrossRef]
- Boyaci-Gunduz, C.P.; Erten, H. Predominant yeasts in the sourdoughs collected from some parts of Turkey. Yeast 2020, 37. [Google Scholar] [CrossRef] [PubMed]
- Middelhoven, W.J.; Kurtzman, C.P.; Vaughan-Martini, A. Saccharomyces bulderi sp. nov., a yeast that ferments gluconolactone. Antonie Van Leeuwenhoek 2000, 77, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Basilio, A.C.M.; de Araujo, P.R.L.; de Morais, J.O.F.; da Silva Filho, E.A.; de Morais, M.A.; Simoes, D.A. Detection and identification of wild yeast contaminants of the industrial fuel ethanol fermentation process. Curr. Microbiol. 2008, 56, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Chen, X.; Guang, J. Bacterial and fungal diversity in the traditional Chinese strong flavour liquor Daqu. J. Inst. Brew. 2019, 125, 443–452. [Google Scholar] [CrossRef]
- Hadaschik, E.; Koschny, R.; Willinger, B.; Hallscheidt, P.; Enk, A.; Hartschuh, W. Pulmonary, rhino-orbital and cutaneous mucormycosis caused by Rhizomucor pusillus in an immunocompromised patient. Clin. Exp. Dermatol. 2012, 37, 355–357. [Google Scholar] [CrossRef]
- Nishino, N.; Hattori, H.; Wada, H.; Touno, E. Biogenic amine production in grass, maize and total mixed ration silages inoculated with Lactobacillus casei or Lactobacillus buchneri. J. Appl. Microbiol. 2007, 103, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Scherer, R.; Gerlach, K.; Südekum, K.H. Biogenic amines and gamma-amino butyric acid in silages: Formation, occurrence and influence on dry matter intake and ruminant production. Anim. Feed Sci. Technol. 2015, 210, 1–16. [Google Scholar] [CrossRef]
- Santos, M.H.S. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | S1 | S2 | S3 | S4 | SEM | p-Value |
|---|---|---|---|---|---|---|
| pH | 6.05 | 5.99 | 6.03 | 6.08 | 0.027 | 0.160 |
| Lactic acid bacteria | 5.08b | 5.78a | 5.69a | 5.10b | 0.122 | 0.011 |
| Escherichia coli | 6.48 | 6.54 | 6.75 | 6.67 | 0.100 | 0.267 |
| Bacteria | 7.18 | 7.28 | 7.21 | 7.24 | 0.074 | 0.782 |
| Yeast | 6.42 | 6.38 | 6.41 | 6.21 | 0.086 | 0.346 |
| DM | 429a | 300c | 304c | 318b | 2.74 | <0.001 |
| Water-soluble carbohydrates | 97.3b | 123a | 128a | 106b | 3.70 | 0.001 |
| Crude protein | 70.0b | 77.8a | 72.2b | 67.3b | 1.30 | 0.003 |
| Neutral detergent fibre | 483b | 473b | 467b | 531a | 4.15 | <0.001 |
| Acid detergent fibre | 269b | 270b | 261c | 281a | 1.40 | <0.001 |
| Acid detergent lignin | 37.8a | 35.4a | 28.1b | 34.0a | 1.18 | 0.002 |
| Items | Days | S1 | S2 | S3 | S4 | SEM | p-Value | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|
| A | D | A*D | ||||||||
| Lactic acid bacteria | 0 | 7.95a | 4.98Bc | 7.03Bb | 8.35a | 0.273 | <0.001 | *** | *** | *** |
| 2 | 8.13a | 5.14Bc | 7.53Bb | 8.47a | 0.159 | <0.001 | ||||
| 5 | 7.69b | 7.03Ac | 8.45Aa | 8.27ab | 0.185 | 0.002 | ||||
| SEM | 0.214 | 0.281 | 0.214 | 0.091 | ||||||
| p-value | 0.389 | 0.004 | 0.009 | 0.373 | ||||||
| Bacteria | 0 | 7.48Aa | 5.92Ab | 4.81Cc | 6.20Cb | 0.272 | <0.001 | *** | *** | *** |
| 2 | 7.53Aa | 4.91Bb | 7.22Aa | 7.92Aa | 0.220 | <0.001 | ||||
| 5 | 4.81Bc | 6.15Ab | 6.29Bb | 7.44Ba | 0.119 | <0.001 | ||||
| SEM | 0.184 | 0.265 | 0.255 | 0.113 | ||||||
| p-value | <0.001 | 0.035 | 0.002 | <0.001 | ||||||
| Yeasts | 0 | 7.73Aa | 4.74c | 6.57b | 7.98a | 0.281 | 0.001 | *** | *** | |
| 2 | 7.79Aa | 5.36b | 6.26b | 7.68a | 0.566 | 0.044 | ||||
| 5 | 5.66Bd | 6.31c | 7.27b | 7.84a | 0.092 | <0.001 | ||||
| SEM | 0.103 | 0.633 | 0.353 | 0.085 | ||||||
| p-value | <0.001 | 0.284 | 0.197 | 0.116 | ||||||
| Items | S1 | S2 | S3 | S4 | SEM | p-Value | |
|---|---|---|---|---|---|---|---|
| Bacteria | Raw reads | 12,485b | 14,773ab | 15,728ab | 17,517a | 1060 | 0.055 |
| Clean reads | 11,407b | 12,746ab | 14,549ab | 16,815a | 1084 | 0.036 | |
| Observed species | 247ab | 347a | 268ab | 183b | 25.6 | 0.013 | |
| Shannon | 2.71b | 5.18a | 2.17b | 2.59b | 0.180 | <0.001 | |
| Simpson | 0.672c | 0.911a | 0.580d | 0.754b | 0.020 | <0.001 | |
| Chao1 | 398a | 506a | 426a | 252b | 41.2 | 0.014 | |
| Fungus | Raw reads | 10,334 | 12,450 | 12,283 | 13,967 | 1203 | 0.279 |
| Clean reads | 9250 | 12,354 | 12,205 | 13,264 | 1394 | 0.272 | |
| Observed species | 144a | 73.3b | 55.7b | 164a | 15.3 | 0.002 | |
| Shannon | 4.14a | 0.779b | 0.548b | 3.67a | 0.22 | <0.001 | |
| Simpson | 0.874a | 0.159b | 0.129b | 0.825a | 0.020 | <0.001 | |
| Chao1 | 195a | 112b | 82.0b | 193a | 18.5 | 0.005 | |
| Items | S1 | S2 | S3 | S4 | SEM | p-Value | |
|---|---|---|---|---|---|---|---|
| Total metabolites | 353b | 4220a | 399b | 753b | 571 | 0.004 | |
| Biogenic amines | Tyramine | 10.5 | 107 | 1.30 | >0 | 26.7 | 0.061 |
| Putrescine | 3.08 | 34.1 | 2.06 | >0 | 8.98 | 0.080 | |
| Phenylethylamine | 0.089b | 0.804a | 0.057b | 0.198b | 0.062 | <0.001 | |
| Spermidine | 0.065b | 0.518a | 0.090b | >0 b | 0.055 | <0.001 | |
| Noradrenaline | 0.013 | 0.019 | 0.004 | 0.019 | 0.010 | 0.694 | |
| Items | Days | S1 | S2 | S3 | S4 | SEM | p-Value | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|
| A | D | A*D | ||||||||
| pH | 0 | 3.97a | 3.61Bb | 3.63Cb | 3.97Ba | 0.011 | <0.001 | *** | *** | *** |
| 2 | 4.00a | 3.69Bc | 3.81Bb | 4.02Ba | 0.006 | <0.001 | ||||
| 5 | 4.01c | 6.32Aa | 4.04Ac | 6.00Ab | 0.051 | <0.001 | ||||
| SEM | 0.017 | 0.049 | 0.013 | 0.030 | ||||||
| p-Value | 0.307 | <0.001 | <0.001 | <0.001 | ||||||
| Lactic acid | 0 | 65.8Ac | 124Aa | 93.7Ab | 53.5Ad | 3.48 | <0.001 | *** | *** | *** |
| 2 | 57.3ABc | 121Aa | 78.8Bb | 49.6Ad | 2.07 | <0.001 | ||||
| 5 | 46.4Ba | 20.1Bbc | 37.6Cab | 9.70Bc | 5.46 | 0.006 | ||||
| SEM | 3.77 | 6.28 | 1.23 | 2.54 | ||||||
| p-Value | 0.029 | <0.001 | <0.001 | <0.001 | ||||||
| Acetic acid | 0 | 42.8b | 28.1Ac | 14.1Bd | 54.6Aa | 2.49 | <0.001 | *** | *** | *** |
| 2 | 36.6b | 23.7Bc | 5.44Cd | 53.1Aa | 1.86 | <0.001 | ||||
| 5 | 33.1b | ND | 44.1Aa | ND | 0.896 | <0.001 | ||||
| SEM | 3.31 | 0.542 | 0.811 | 1.45 | ||||||
| p-Value | 0.118 | <0.001 | <0.001 | <0.001 | ||||||
| Propionic acid | 0 | ND | ND | ND | ND | - | - | *** | *** | *** |
| 2 | ND | ND | ND | ND | - | - | ||||
| 5 | 1.95c | 12.7Aa | ND | 8.95Ab | 0.726 | <0.001 | ||||
| SEM | 0.562 | 0.541 | - | 0.304 | ||||||
| p-Valuee | 0.078 | <0.001 | - | <0.001 | ||||||
| Ammonia nitrogen | 0 | 48.1Ab | 45.0Ab | 39.5Ac | 53.1a | 1.33 | <0.001 | *** | *** | *** |
| 2 | 47.4Ab | 41.9Ac | 40.2Ac | 53.1a | 0.689 | <0.001 | ||||
| 5 | 40.5Bb | 28.6Bc | 22.2Bd | 55.2a | 1.30 | <0.001 | ||||
| SEM | 0.841 | 1.38 | 1.10 | 1.20 | ||||||
| p-Value | 0.001 | <0.001 | <0.001 | 0.403 | ||||||
| Items | S1 | S2 | S3 | S4 | SEM | p-Value |
|---|---|---|---|---|---|---|
| DM | 435a | 299b | 305b | 316b | 4.20 | <0.001 |
| Water-soluble carbohydrates | 12.1c | 23.7b | 27.2b | 35.6a | 1.15 | <0.001 |
| Crude protein | 68.5b | 72.4a | 70.9a | 66.7c | 0.555 | <0.001 |
| Neutral detergent fibre | 471b | 465b | 460b | 526a | 7.21 | <0.001 |
| Acid detergent fibre | 265 | 267 | 255 | 266 | 9.87 | 0.832 |
| Acid detergent lignin | 35.5 | 31.7 | 21.3 | 33.9 | 4.37 | 0.173 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Sun, L.; Xu, H.; Na, N.; Yin, G.; Liu, S.; Jiang, Y.; Xue, Y. Microbial Communities, Metabolites, Fermentation Quality and Aerobic Stability of Whole-Plant Corn Silage Collected from Family Farms in Desert Steppe of North China. Processes 2021, 9, 784. https://doi.org/10.3390/pr9050784
Wang C, Sun L, Xu H, Na N, Yin G, Liu S, Jiang Y, Xue Y. Microbial Communities, Metabolites, Fermentation Quality and Aerobic Stability of Whole-Plant Corn Silage Collected from Family Farms in Desert Steppe of North China. Processes. 2021; 9(5):784. https://doi.org/10.3390/pr9050784
Chicago/Turabian StyleWang, Chao, Lin Sun, Haiwen Xu, Na Na, Guomei Yin, Sibo Liu, Yun Jiang, and Yanlin Xue. 2021. "Microbial Communities, Metabolites, Fermentation Quality and Aerobic Stability of Whole-Plant Corn Silage Collected from Family Farms in Desert Steppe of North China" Processes 9, no. 5: 784. https://doi.org/10.3390/pr9050784
APA StyleWang, C., Sun, L., Xu, H., Na, N., Yin, G., Liu, S., Jiang, Y., & Xue, Y. (2021). Microbial Communities, Metabolites, Fermentation Quality and Aerobic Stability of Whole-Plant Corn Silage Collected from Family Farms in Desert Steppe of North China. Processes, 9(5), 784. https://doi.org/10.3390/pr9050784