

Evaluation of Ilex guayusa and Piper marginatum Extract Cytotoxicity on Human Dental Pulp Mesenchymal Stem Cells

 ,
, 
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture of Human Dental Pulp Stem Cells (hDPSC)
2.2. Nanobiochip Elaboration
2.3. hDPSC Seeding on Nanobiochips
2.4. Ilex guayusa and Piper marginatum Extracts
2.5. Assessment of Plant Extract Cytotoxicity on hDPSC
2.6. Cellular Viability Analysis by Raman Spectroscopy
2.7. Cellular Viability Analysis by Fluorescence Microscopy
2.8. Cellular Viability Analysis Using Flow Cytometry
2.9. Statistical Analysis
3. Results
3.1. Cellular Viability Analysis Using Raman Spectroscopy
3.2. Cellular Viability Analysis Using Fluorescence Microscopy
3.3. Cellular Viability Analysis Using Flow Cytometry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bloch-Zupan, A.; Rey, T.; Jimenez-Armijo, A.; Kawczynski, M.; Kharouf, N.; O-Rare consortium; Dure-Molla, M.; Noirrit, E.; Hernandez, M.; Joseph-Beaudin, C.; et al. Amelogenesis imperfecta: Next-generation sequencing sheds light on Witkop’s classification. Front. Physiol. 2023, 14, 1130175. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P.J.M.; Aldred, M.; Bloch-Zupan, A. Amelogenesis imperfecta. Orphanet J. Rare Dis. 2007, 2, 17. [Google Scholar] [CrossRef]
- Sabandal, M.M.; Schafer, E. Amelogenesis imperfecta: Review of diagnostic findings and treatment concepts. Odontology 2016, 104, 245–256. [Google Scholar] [CrossRef]
- Seow, W.K. Developmental defects of enamel and dentine: Challenges for basic science research and clinical management. Aust. Dent. J. 2014, 59 (Suppl. S1), 143–154. [Google Scholar] [CrossRef]
- Strauch, S.; Hahnel, S. Restorative Treatment in Patients with Amelogenesis Imperfecta: A Review. J. Prosthodont. 2018, 27, 618–623. [Google Scholar] [CrossRef]
- Farooq, I.; Bugshan, A. The role of salivary contents and modern technologies in the remineralization of dental enamel: A narrative review. F1000Research 2020, 9, 171. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Joiner, A.; Chang, J. The remineralisation of enamel: A review of the literature. J. Dent. 2014, 42 (Suppl. S1), S12–S20. [Google Scholar] [CrossRef] [PubMed]
- Sequeda-Castañeda, L.G.; Gutiérrez, S.; Chacín, A.; Contreras, D.; Penedo, G.; Moreno, G.; Galvis, P.; Gamboa, F.; Luengas, P. Characterisation of the remineralisation effect of Piper marginatum Jacq. and Ilex guayusa Loes extracts on tooth enamel. In Proceedings of the IV Latin American Congress of Medicinal Plants, Barranquilla, Colombia, 17–19 August 2016. [Google Scholar]
- Schiffner, U. Use of fluorides for caries prevention. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2021, 64, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Krzeczkowski, J.E.; Hall, M.; Saint-Amour, D.; Oulhote, Y.; McGuckin, T.; Goodman, C.V.; Green, R.; Muckle, G.; Lanphear, B.; Till, C. Prenatal fluoride exposure, offspring visual acuity and autonomic nervous system function in 6-month-old infants. Environ. Int. 2024, 183, 108336. [Google Scholar] [CrossRef]
- Żwierełło, W.; Maruszewska, A.; Skórka-Majewicz, M.; Gutowska, I. Fluoride in the Central Nervous System and Its Potential Influence on the Development and Invasiveness of Brain Tumours-A Research Hypothesis. Int. J. Mol. Sci. 2023, 24, 1558. [Google Scholar] [CrossRef]
- Gong, Q.M.; Ling, J.Q.; Wei, X. Research progress in the pathogenesis mechanism of dental fluorosis. Zhonghua Kou Qiang Yi Xue Za Zhi 2023, 58, 217–223. [Google Scholar] [PubMed]
- Miranda, G.H.N.; Alencar de Oliveira Lima, L.; Bittencourt, L.O.; Dos Santos, S.M.; Platini Caldas de Souza, M.; Nogueira, L.S.; de Oliveira, E.H.C.; Monteiro, M.C.; Dionizio, A.; Leite, A.L.; et al. Effects of long-term fluoride exposure are associated with oxidative biochemistry impairment and global proteomic modulation, but not genotoxicity, in parotid glands of mice. PLoS ONE 2022, 17, e0261252. [Google Scholar] [CrossRef] [PubMed]
- Miranda, G.H.N.; Ferreira, M.K.M.; Bittencourt, L.O.; Lima, L.A.O.; Puty, B.; Lima, R.R. Chapter 17—The role of oxidative stress in fluoride toxicity. Toxicology 2021, 157–163. [Google Scholar] [CrossRef]
- Aggarwal, A.; Bhushan, P. Fluoride toxicity: A review article on adverse effects of fluoride. Santosh Univ. J. Health Sci. 2019, 5, 3–4. [Google Scholar] [CrossRef]
- Zuo, H.; Chen, L.; Kong, M.; Qiu, L.; Lu, P.; Wu, P.; Yang, Y.; Chen, K. Toxic effects of fluoride on organisms. Life Sci. 2018, 198, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, T.; Rawls, H.R. Fluoride Exposure in Early Life as the Possible Root Cause of Disease In Later Life. J. Clin. Pediatr. Dent. 2018, 42, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Goschorska, M.; Baranowska-Bosiacka, I.; Gutowska, I.; Metryka, E.; Skórka-Majewicz, M.; Chlubek, D. Potential Role of Fluoride in the Etiopathogenesis of Alzheimer’s Disease. Int. J. Mol. Sci. 2018, 19, 3965. [Google Scholar] [CrossRef]
- Kanduti, D.; Sterbenk, P.; Artnik, B. Fluoride: A Review of Use and Effects on Health. Mater. Sociomed. 2016, 28, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Bandoski, C.; Bartlett, J.D. Fluoride induces oxidative damage and SIRT1/autophagy through ROS-mediated JNK signaling. Free. Radic. Biol. Med. 2015, 89, 369–378. [Google Scholar] [CrossRef]
- Jha, S.K.; Singh, R.K.; Damodaran, T.; Mishra, V.K.; Sharma, D.K.; Rai, D. Fluoride in groundwater: Toxicological exposure and remedies. J. Toxicol. Environ. Health B Crit. Rev. 2013, 16, 52–66. [Google Scholar] [CrossRef]
- Martínez-Mier, E. Fluoride: Its Metabolism, Toxicity, and Role in Dental Health. J. Evid.-Based Complement. Altern. Med. 2012, 17, 28–32. [Google Scholar] [CrossRef]
- Whitford, G.M. Acute toxicity of ingested fluoride. Monogr. Oral. Sci. 2011, 22, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Jimenez, L.; Soria Fregozo, C.; Miranda Beltran, M.L.; Gutierrez Coronado, O.; Perez Vega, M.I. Effects of the fluoride on the central nervous system. Neurologia 2011, 26, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Barbier, O.; Arreola-Mendoza, L.; Del Razo, L.M. Molecular mechanisms of fluoride toxicity. Chem.-Biol. Interact. 2010, 188, 319–333. [Google Scholar] [CrossRef] [PubMed]
- He, L.F.; Chen, J.G. DNA damage, apoptosis and cell cycle changes induced by fluoride in rat oral mucosal cells and hepatocytes. World J. Gastroenterol. 2006, 12, 1144–1148. [Google Scholar] [CrossRef] [PubMed]
- Shulman, J.D.; Wells, L.M. Acute fluoride toxicity from ingesting home-use dental products in children, birth to 6 years of age. J. Public Health Dent. 1997, 57, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Whitford, G.M. Fluoride in dental products: Safety considerations. J. Dent. Res. 1987, 66, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Bayless, J.M.; Tinanoff, N. Diagnosis and treatment of acute fluoride toxicity. J. Am. Dent. Assoc. 1985, 110, 209–211. [Google Scholar] [CrossRef]
- Tubert-Jeannin, S.; Auclair, C.; Amsallem, E.; Tramini, P.; Gerbaud, L.; Ruffieux, C.; Schulte, A.G.; Koch, M.J.; Rège-Walther, M.; Ismail, A. Fluoride supplements (tablets, drops, lozenges or chewing gums) for preventing dental caries in children. Cochrane Database Syst. Rev. 2011, 2011, CD007592. [Google Scholar] [CrossRef]
- O’Mullane, D.M.; Baez, R.J.; Jones, S.; Lennon, M.A.; Petersen, P.E.; Rugg-Gunn, A.J.; Whelton, H.; Whitford, G.M. Fluoride and Oral Health. Community Dent. Health 2016, 33, 69–99. [Google Scholar]
- Gonzalez-Gonzalez, R.; Bologna-Molina, R.; Molina-Frechero, N. Editorial: Fluoride exposure, dental fluorosis, and health. Front. Oral Health 2023, 4, 1256495. [Google Scholar] [CrossRef] [PubMed]
- Peckham, S.; Awofeso, N. Water fluoridation: A critical review of the physiological effects of ingested fluoride as a public health intervention. Sci. World J. 2014, 2014, 293019. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.J.; Connor, R.O.; Holmes, R.; Landes, D.; Shah, K.; Tanday, A.; Vernazza, C. Dental fluorosis. Br. Dent. J. 2022, 232, 492. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.S.; Wei, X.; Ling, J.Q. Etiology, diagnosis, prevention and treatment of dental fluorosis. Zhonghua Kou Qiang Yi Xue Za Zhi 2020, 55, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Lima, I.F.P.; Nóbrega, D.F.; Cericato, G.O.; Ziegelmann, P.K.; Paranhos, L.R. Prevalence of dental fluorosis in regions supplied with non-fluoridated water in the Brazilian territory: A systematic review and meta-analysis. Cienc. Saude Coletiva 2019, 24, 2909–2922. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.O.; Martins Ferreira, M.K.; Davis, L.; Bittencourt, L.O.; Bragança Aragão, W.A.; Dionizio, A.; Rabelo Buzalaf, M.A.; Crespo-Lopez, M.E.; Maia, C.S.F.; Lima, R.R. Effects of Fluoride Long-Term Exposure over the Cerebellum: Global Proteomic Profile, Oxidative Biochemistry, Cell Density, and Motor Behavior Evaluation. Int. J. Mol. Sci. 2020, 21, 7297. [Google Scholar] [CrossRef] [PubMed]
- Taher, M.K.; Momoli, F.; Go, J.; Hagiwara, S.; Ramoju, S.; Hu, X.; Jensen, N.; Terrell, R.; Hemmerich, A.; Krewski, D. Systematic review of epidemiological and toxicological evidence on health effects of fluoride in drinking water. Crit. Rev. Toxicol. 2024, 54, 2–34. [Google Scholar] [CrossRef] [PubMed]
- Adkins, E.A.; Brunst, K.J. Impacts of Fluoride Neurotoxicity and Mitochondrial Dysfunction on Cognition and Mental Health: A Literature Review. Int. J. Environ. Res. Public Health 2021, 18, 12884. [Google Scholar] [CrossRef] [PubMed]
- Susheela, A.K.; Kumar, A.; Bhatnagar, M.; Bahadur, R. Prevalence of endemic fluorosis with gastrointestinal manifestations in people living in some North-Indian villages. Fluoride—Q. Rep. 1993, 26, 97–104. [Google Scholar]
- Anushri, M.; Yashoda, R.; Puranik, M.P. Herbs: A Good Alternatives to Current Treatments for Oral Health Problems. Int. J. Adv. Health Sci. 2015, 1, 26–32. [Google Scholar]
- Celik, Z.C.; Yavlal, G.O.; Yanıkoglu, F.; Kargul, B.; Tagtekin, D.; Stookey, G.K.; Peker, S.; Hayran, O. Do Ginger Extract, Natural Honey and Bitter Chocolate Remineralize Enamel Surface as Fluoride Toothpastes? An In-Vitro Study. Niger. J. Clin. Pract. 2021, 24, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Chandna, P.; Srivastava, N.; Ali, S. Remineralizing Agents: The Next Frontier. Curr. Clin. Pharmacol. 2016, 11, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Kanth, M.R.; Prakash, A.R.; Sreenath, G.; Reddy, V.S.; Huldah, S. Efficacy of Specific Plant Products on Microorganisms Causing Dental Caries. J. Clin. Diagn. Res. 2016, 10, ZM01–ZM03. [Google Scholar] [CrossRef] [PubMed]
- Taheri, J.B.; Azimi, S.; Rafieian, N.; Akhavan Zanjani, H. Herbs in dentistry. Int. Dent. J. 2011, 61, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.P.; Li, J.Y.; Hao, Y.Q.; Zhou, X.D. Effect of compounds of Galla chinensis on remineralisation of initial enamel carious lesions in vitro. J. Dent. 2007, 35, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Liu, M.D.; Li, J.Y.; Zhou, X.D.; ten Cate, J.M. Chemical composition of Galla chinensis extract and the effect of its main component(s) on the prevention of enamel demineralization in vitro. Int. J. Oral. Sci. 2012, 4, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Sequeda-Castañeda, L.G.; Gutiérrez-Prieto, S.J.; Luengas-Caicedo, P.E. Plants native to colombia and their potential use in diseases of the oral cavity. In Proceedings of the XVIII Colombian Chemistry Congress, Popayán, Colombia, 6–8 November 2019. [Google Scholar]
- Sequeda-Castañeda, L.G.; Costa, M.G.; Célis, C.; Gutiérrez-Prieto, S.J.; Luengas, P. Ilex guayusa Loes (Aquifoliaceae): Amazon and andean native plant. Pharmacologyonline 2016, 3, 193–202. [Google Scholar]
- Wise, G.; Negrin, A. A critical review of the composition and history of safe use of guayusa: A stimulant and antioxidant novel food. Crit. Rev. Food Sci. Nutr. 2020, 60, 2393–2404. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Khan, T.; Sharifi-Rad, J.; Ozleyen, A.; Turkdonmez, E.; et al. Piper Species: A Comprehensive Review on Their Phytochemistry, Biological Activities and Applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef]
- Sequeda-Castañeda, L.G.; Célis, C.; Gutiérrez-Prieto, S.J. Piper marginatum Jacq. (Piperaceae): Phytochemical, therapeutic, botanical insecticidal and phytosanitary uses. Pharmacologyonline 2015, 3, 136–145. [Google Scholar]
- Brú, J.; Guzman, J.D. Folk medicine, phytochemistry and pharmacological application of Piper marginatum. Rev. Bras. Farmacogn. 2016, 26, 767–779. [Google Scholar] [CrossRef]
- Erazo-Garcia, M.P.; Guadalupe, J.J.; Rowntree, J.K.; Borja-Serrano, P.; Espinosa de los Monteros-Silva, N.; Torres, M.d.L. Assessing the Genetic Diversity of Ilex guayusa Loes., a Medicinal Plant from the Ecuadorian Amazon. Diversity 2021, 13, 182. [Google Scholar] [CrossRef]
- Pereira, L.A.; Barboza, G.E.; Bovini, M.G.; De Almeida, M.Z.; Guimarães, E.F. Caracterización y uso de “Pimientas” en una comunidad quilombola de la amazonía oriental (Brasil). J. Bot. Res. Inst. Tex. 2011, 5, 255–272. [Google Scholar]
- Gamboa, F.; Munoz, C.C.; Numpaque, G.; Sequeda-Castaneda, L.G.; Gutierrez, S.J.; Tellez, N. Antimicrobial Activity of Piper marginatum Jacq and Ilex guayusa Loes on Microorganisms Associated with Periodontal Disease. Int. J. Microbiol. 2018, 2018, 4147383. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Prieto, S.J.; Sequeda-Castaneda, L.G.; Penedo-Jaramillo, G.M.; Chacin-Nieto, A.V.; Contreras-Caceres, D.R.; Moreno-Abello, G.C.; Galvis-Rincon, M.P.; Gamboa-Jaimes, F.O.; Luengas-Caicedo, P.E. In vitro mineral apposition analysis of two Colombian plant extracts on Amelogenesis imperfecta teeth. Clin. Exp. Dent. Res. 2022, 8, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Ardalan, M.; Rafieian-kopaei, M. Is the safety of herbal medicines for kidneys under question? J. Nephropharmacol. 2013, 2, 11–12. [Google Scholar] [PubMed]
- Nasri, H.; Shirzad, H. Toxicity and Safety of Medicinal Plants. J. HerbMed Pharmacol. 2013, 2, 21–22. [Google Scholar]
- Lanini, J.; Duarte-Almeida, J.M.; Nappo, S.A.; Carlini, E.A. Are medicinal herbs safe? The opinion of plant vendors from Diadema (São Paulo, southeastern Brazil). Rev. Bras. Farmacogn. 2012, 22, 21–28. [Google Scholar] [CrossRef]
- ATSDR. Module I—Introduction to Toxicology. Available online: https://www.atsdr.cdc.gov/index.html (accessed on 3 March 2024).
- Hassan, A.K.; Venkatesh, Y.P. An overview of fruit allergy and the causative allergens. Eur. Ann. Allergy Clin. Immunol. 2015, 47, 180–187. [Google Scholar]
- Phillips, J.F.; Jelks, M.L.; Lockey, R.F. Important Florida botanical aeroallergens. Allergy Asthma Proc. 2010, 31, 337–340. [Google Scholar] [CrossRef]
- Ghorani-Azam, A.; Sepahi, S.; Riahi-Zanjani, B.; Alizadeh Ghamsari, A.; Mohajeri, S.A.; Balali-Mood, M. Plant toxins and acute medicinal plant poisoning in children: A systematic literature review. J. Res. Med. Sci. 2018, 23, 26. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.H.; Bayrami, Z.; Farzaei, F.; Aneva, I.Y.; Das, S.K.; Patra, J.K.; Das, G.; Abdollahi, M. Poisoning by Medical Plants. Arch. Iran. Med. 2020, 23, 117–127. [Google Scholar] [PubMed]
- Moes, S.; van-Gestel, K.; van-Beek, G. Toxicology. Available online: https://maken.wikiwijs.nl/147644/ (accessed on 3 March 2024).
- Lee, J.; Mohns, M. How to Choose a Cell Viability or Cytotoxicity Assay. Available online: https://www.promega.com/products/cell-health-assays/ (accessed on 28 May 2024).
- Gilbert, D.F.; Friedrich, O. Cell Viability Assays: Methods and Protocols; Springer: New York, NY, USA, 2023. [Google Scholar]
- Molecular-Probes. LIVE/DEAD® Viability/Cytotoxicity Kit *for mammalian cells. Invit. Detect. Technol. 2005, 7. Available online: https://assets.fishersci.com/TFS-Assets/LSG/manuals/LIVE_DEAD_Viability_Cytotoxicity_Kit_for_mammalian_cells_J1_21Dec2005.pdf (accessed on 2 February 2017).
- Verrier, S.; Zoladek, A.; Notingher, I. Raman micro-spectroscopy as a non-invasive cell viability test. Methods Mol. Biol. 2011, 740, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Kummrow, A.; Frankowski, M.; Bock, N.; Werner, C.; Dziekan, T.; Neukammer, J. Quantitative assessment of cell viability based on flow cytometry and microscopy. Cytometry A 2013, 83, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Han, X.X.; Rodriguez, R.S.; Haynes, C.L.; Ozaki, Y.; Zhao, B. Surface-enhanced Raman spectroscopy. Nat. Rev. Methods Primers 2022, 1, 17. [Google Scholar] [CrossRef]
- Ma, X.; Sun, X.; Wang, H.; Wang, Y.; Chen, D.; Li, Q. Raman Spectroscopy for Pharmaceutical Quantitative Analysis by Low-Rank Estimation. Front. Chem. 2018, 6, 400. [Google Scholar] [CrossRef] [PubMed]
- Movasaghi, Z.; Rehman, S.; Rehman, I.U. Raman Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2007, 42, 493–541. [Google Scholar] [CrossRef]
- Nie, S.; Emory, S.R. Probing Single Molecules and Single Nanoparticles by Surface-Enhanced Raman Scattering. Science 1997, 275, 1102–1106. [Google Scholar]
- Otel, I. Overall Review on Recent Applications of Raman Spectroscopy Technique in Dentistry. Quantum Beam Sci. 2023, 7, 5. [Google Scholar] [CrossRef]
- Petersen, M.; Yu, Z.; Lu, X. Application of Raman Spectroscopic Methods in Food Safety: A Review. Biosensors 2021, 11, 187. [Google Scholar] [CrossRef]
- Pezzotti, G. Raman spectroscopy in cell biology and microbiology. J. Raman Spectrosc. 2021, 52, 2348–2443. [Google Scholar] [CrossRef]
- Zhang, X.; Yonzon, C.R.; Young, M.A.; Stuart, D.A.; Van Duyne, R.P. Surface-enhanced Raman spectroscopy biosensors: Excitation spectroscopy for optimisation of substrates fabricated by nanosphere lithography. IEE Proc. Nanobiotechnol. 2005, 152, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Combs, C.A.; Shroff, H. Fluorescence Microscopy: A Concise Guide to Current Imaging Methods. Curr. Protoc. Neurosci. 2017, 79, 2.1.1–2.1.25. [Google Scholar] [CrossRef]
- Gabriel, M.V. Advanced Fluorescence Microscopy Techniques Applied to the Quantitative Study of the Dynamics Intracellular. Ph.D. Thesis, Universidad de Buenos Aires, Buenos Aires, Argentina, 2017. [Google Scholar]
- Kubitscheck, U.; Bloeck, F.; Wiley-Blackwell. Fluorescence Microscopy: From Principles to Biological Applications; Wiley-VCH: Weinheim, Germany, 2017. [Google Scholar]
- Weaver, J.L. Estimation of Cell Viability by Flow Cytometry. In Flow Cytometry Protocols; Jaroszeski, M.J., Heller, R., Eds.; Humana Press: Totowa, NJ, USA, 1998; pp. 77–83. [Google Scholar]
- DPSC. Poietics™ Human Dental Pulp Stem Cells—Instructions for Use. EU/ROW Sci. Support 2013, 4. Available online: https://bioscience.lonza.com/lonza_bs/CO/en/document/download/29343 (accessed on 2 February 2017).
- Varela-Fonseca, S.; Montero-Zeledón, E.; Rojas-Rojas, L.; Varela-Fonseca, A.; Gutiérrez-Fallas, D. Raman spectroscopy analysis of DNA using the SERS method. Rev. Tecnol. Marcha 2019, 32, 118–125. [Google Scholar] [CrossRef]
- Castañeda-Uribe, O.A.; Salcedo-Reyes, J.C.; Méndez-Pinzón, H.A.; Pedroza-Rodríguez, A.M. Fabrication and optical characterisation of a high quality photonic crystal based on fcc-opal grown by vertical convective self-assembly method. Univ. Sci. 2010, 15, 150–158. [Google Scholar] [CrossRef]
- Hallab, N.J.; Bundy, K.J.; O’Connor, K.; Moses, R.L.; Jacobs, J.J. Evaluation of metallic and polymeric biomaterial surface energy and surface roughness characteristics for directed cell adhesion. Tissue Eng. 2001, 7, 55–71. [Google Scholar] [CrossRef]
- Kneipp, K.; Kneipp, H.; Corio, P.; Brown, S.D.; Shafer, K.; Motz, J.; Perelman, L.T.; Hanlon, E.B.; Marucci, A.; Dresselhaus, G.; et al. Surface-enhanced and normal stokes and anti-stokes Raman spectroscopy of single-walled carbon nanotubes. Phys. Rev. Lett. 2000, 84, 3470–3473. [Google Scholar] [CrossRef]
- Salcedo-Reyes, J.C.; Méndez-Pinzón, H.A.; Jiménez-Borrego, L.C. Method for the Fabrication of a Thin Film Consisting of a Colloidal Crystal Infiltrated with the Luminescent Polymer MDMO-PPV Formed from Silica Spheres (SIO2), with Face-Centred Cubic (fcc) Structure. U.S. Patent 9,859,497 B2, 2 January 2018. [Google Scholar]
- Sequeda-Castañeda, L.G. Phytochemical Study of Some Colombian Native Plants and Evaluation of Their Antimicrobial Activity and Their Remineralizing Effect on Tooth Enamel. Ph.D. Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 2024. [Google Scholar]
- Rasband, W. ImageJ. Available online: https://imagej.net/ij/ (accessed on 30 June 2017).
- Grishagin, I.V. Automatic cell counting with ImageJ. Anal. Biochem. 2015, 473, 63–65. [Google Scholar] [CrossRef]
- BD-Pharmigen. PE Annexin V Apoptosis Detection Kit I. Tech. Data Sheet 1998, 559763 Rev. 8, 3. Available online: https://www.bdbiosciences.com/en-us/products/reagents/flow-cytometry-reagents/research-reagents/panels-multicolor-cocktails-ruo/pe-annexin-v-apoptosis-detection-kit-i.559763 (accessed on 2 February 2017).
- FlowJo. FlowJoTM Portal Documentation. Available online: https://www.flowjo.com/solutions/flowjo/request-quote (accessed on 30 June 2017).
- Di-Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat. Available online: http://www.infostat.com.ar/ (accessed on 30 June 2017).
- IBM-Corp. IBM SPSS Statistics 29.0. Available online: https://www.ibm.com/products/spss-statistics?lot=5 (accessed on 30 June 2017).
- González-Solís, L.; Martinez-Espinosa, J.C.; Martínez-Zérega, B.; Palomares-Anda, P.; Torres-Gonzalez, L.; Vargas-Rodriguez, H.; Gallegos, A.; Gonzalez Silva, R.; Juárez-López, A.; Espinoza-Padilla, P. Aplicaciones de la técnica de espectroscopia Raman de superficie amplificada en el diagnóstico del cáncer basado en el análisis de muestras de suero sanguíneo. Ide@s CONCYTEG 2012, 7, 1091. [Google Scholar]
- Jung, G.B.; Lee, Y.J.; Lee, G.; Park, H.K. A simple and rapid detection of tissue adhesive-induced biochemical changes in cells and DNA using Raman spectroscopy. Biomed. Opt. Express 2013, 4, 2673–2682. [Google Scholar] [CrossRef]
- Nothinger. Raman Spectroscopy Cell-based Biosensors. Semnsor 2007, 7, 1343–1358. [Google Scholar]
- Notingher, I.; Hench, L.L. Raman microspectroscopy: A noninvasive tool for studies of individual living cells in vitro. Expert. Rev. Med. Devices 2006, 3, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Talari, A.; Movasaghi, Z.; Rehman, S.; Rehman, I. Raman Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2014, 50, 46. [Google Scholar] [CrossRef]
- Alraies, A.; Canetta, E.; Waddington, R.J.; Moseley, R.; Sloan, A.J. Discrimination of Dental Pulp Stem Cell Regenerative Heterogeneity by Single-Cell Raman Spectroscopy. Tissue Eng. Part C Methods 2019, 25, 489–499. [Google Scholar] [CrossRef]
- Colceriu-Simon, I.M.; Virag, P.; Salmon, B.; Tarmure, V.; Baciut, M.; Bran, S.; Jacobs, R.; Falamas, A. Raman Micro-Spectroscopy of Dental Pulp Stem Cells: An Approach to Monitor the Effects of Cone Beam Computed Tomography Low-Dose Ionizing Radiation. Anal. Lett. 2018, 52, 1–15. [Google Scholar] [CrossRef]
- Colthup, N.B.; Daly, L.H.; Wiberley, S.E. Introduction to Infrared and Raman Spectroscopy; Elsevier Science: Amsterdam, The Netherlands, 1990. [Google Scholar]
- Hobro, A.J.; Rouhi, M.; Blanch, E.W.; Conn, G.L. Raman and Raman optical activity (ROA) analysis of RNA structural motifs in Domain I of the EMCV IRES. Nucleic Acids Res. 2007, 35, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, G.G. Infrared and Raman Spectroscopy: Principles and Applications; De Gruyter: Berlin, Germany, 2023. [Google Scholar]
- Larkin, P. Infrared and Raman Spectroscopy: Principles and Spectral Interpretation; Elsevier Science: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Lin-Vien, D.; Colthup, N.B.; Fateley, W.G.; Grasselli, J.G. The Handbook of Infrared and Raman Characteristic Frequencies of Organic Molecules; Elsevier Science: Amsterdam, The Netherlands, 1991. [Google Scholar]
- Madzharova, F.; Heiner, Z.; Guhlke, M.; Kneipp, J. Surface-Enhanced Hyper-Raman Spectra of Adenine, Guanine, Cytosine, Thymine, and Uracil. J. Phys. Chem. C Nanomater Interfaces 2016, 120, 15415–15423. [Google Scholar] [CrossRef]
- Mitchell, A.; Ashton, L.; Yang, X.B.; Goodacre, R.; Tomlinson, M.J.; Smith, A.; Kirkham, J. Aseptic Raman spectroscopy can detect changes associated with the culture of human dental pulp stromal cells in osteoinductive culture. Analyst 2015, 140, 7347–7354. [Google Scholar] [CrossRef]
- Simonović, J.; Toljić, B.; Rašković, B.; Jovanović, V.; Lazarević, M.; Milošević, M.; Nikolić, N.; Panajotović, R.; Milašin, J. Raman microspectroscopy: Toward a better distinction and profiling of different populations of dental stem cells. Croat Med. J. 2019, 60, 78–86. [Google Scholar] [CrossRef]
- Wang, J.; Qi, G.; Qu, X.; Ling, X.; Zhang, Z.; Jin, Y. Molecular Profiling of Dental Pulp Stem Cells during Cell Differentiation by Surface Enhanced Raman Spectroscopy. Anal. Chem. 2020, 92, 3735–3741. [Google Scholar] [CrossRef] [PubMed]
- Zoladek, A.; Pascut, F.C.; Patel, P.; Notingher, I. Non-invasive time-course imaging of apoptotic cells by confocal Raman micro-spectroscopy. J. Raman Spectrosc. 2011, 42, 251–258. [Google Scholar] [CrossRef]
- Bergholt, M.S.; Serio, A.; Albro, M.B. Raman Spectroscopy: Guiding Light for the Extracellular Matrix. Front. Bioeng. Biotechnol. 2019, 7, 303. [Google Scholar] [CrossRef] [PubMed]
- Ember, K.J.I.; Hoeve, M.A.; McAughtrie, S.L.; Bergholt, M.S.; Dwyer, B.J.; Stevens, M.M.; Faulds, K.; Forbes, S.J.; Campbell, C.J. Raman spectroscopy and regenerative medicine: A review. NPJ Regen. Med. 2017, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.R.; Hooper, D.C.; Zhang, L.; Wolverson, D.; Valev, V.K. Raman Techniques: Fundamentals and Frontiers. Nanoscale Res. Lett. 2019, 14, 231. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Ishigaki, M.; Taketani, A.; Andriana, B.B. Raman spectroscopy and its use for live cell and tissue analysis. Biomed. Spectrosc. Imaging 2018, 7, 97–104. [Google Scholar] [CrossRef]
- Sato, H.; Popp, J.; Wood, B.R.; Ozaki, Y. Raman Spectroscopy in Human Health And Biomedicine; World Scientific Publishing Company: Singapore, 2023. [Google Scholar]
- Fazio, E.; Speciale, A.; Spadaro, S.; Bonsignore, M.; Cimino, F.; Cristani, M.; Trombetta, D.; Saija, A.; Neri, F. Evaluation of biological response induced by molybdenum oxide nanocolloids on in vitro cultured NIH/3T3 fibroblast cells by micro-Raman spectroscopy. Colloids Surf. B Biointerfaces 2018, 170, 233–241. [Google Scholar] [CrossRef] [PubMed]
- De-Gelder, J.; De-Gussem, K.; Vandenabeele, P.; Moens, L. Reference database of Raman spectra of biological molecules. J. Raman Spectrosc. 2007, 38, 1133–1147. [Google Scholar] [CrossRef]
- Vandenabeele, P. Practical Raman Spectroscopy—An Introduction; Wiley: Ghent, Belgium, 2013. [Google Scholar]
- Langer, J.; JimenezdeAberasturi, D.; Aizpurua, J.; Alvarez-Puebla, R.A.; Auguié, B.; Baumberg, J.J.; Bazan, G.C.; Bell, S.E.J.; Boisen, A.; Brolo, A.G.; et al. Present and Future of Surface-Enhanced Raman Scattering. ACS Nano 2020, 14, 28–117. [Google Scholar] [CrossRef]
- Li, C.; Huang, Y.; Li, X.; Zhang, Y.; Chen, Q.; Ye, Z.; Alqarni, Z.; Bell, S.E.J.; Xu, Y. Towards practical and sustainable SERS: A review of recent developments in the construction of multifunctional enhancing substrates. J. Mater. Chem. C 2021, 9, 11517–11552. [Google Scholar] [CrossRef]
- Pilot, R.; Signorini, R.; Durante, C.; Orian, L.; Bhamidipati, M.; Fabris, L. A Review on Surface-Enhanced Raman Scattering. Biosensors 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, K.; Stiebing, C.; Matthäus, C.; Schmitt, M.; Popp, J. Advantages and limitations of Raman spectroscopy for molecular diagnostics: An update. Expert Rev. Mol. Diagn. 2015, 15, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-J.; Lei, Z.-C.; Li, J.; Zong, C.; Yang, C.J.; Ren, B. Label-Free Surface-Enhanced Raman Spectroscopy Detection of DNA with Single-Base Sensitivity. J. Am. Chem. Soc. 2015, 137, 5149–5154. [Google Scholar] [CrossRef] [PubMed]
- Borges-Walmsley, M.I.; Walmsley, A.R. The structure and function of drug pumps. Trends Microbiol. 2001, 9, 71–79. [Google Scholar] [CrossRef] [PubMed]
- McKeegan, K.S.; Borges-Walmsley, M.I.; Walmsley, A.R. The structure and function of drug pumps: An update. Trends Microbiol. 2003, 11, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chan, A.M.; Liu, Y.; Wang, X.; Holbrook, N.J. Serum withdrawal and etoposide induce apoptosis in human lung carcinoma cell line A549 via distinct pathways. Apoptosis 1997, 2, 199–206. [Google Scholar] [CrossRef]
- Graziano, A.; d’Aquino, R.; Laino, G.; Papaccio, G. Dental pulp stem cells: A promising tool for bone regeneration. Stem Cell Rev. 2008, 4, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Bartold, P.M.; Miura, M.; Seo, B.M.; Robey, P.G.; Gronthos, S. The efficacy of mesenchymal stem cells to regenerate and repair dental structures. Orthod. Craniofacial Res. 2005, 8, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Gioventu, S.; Andriolo, G.; Bonino, F.; Frasca, S.; Lazzari, L.; Montelatici, E.; Santoro, F.; Rebulla, P. A novel method for banking dental pulp stem cells. Transfus. Apher. Sci. 2012, 47, 199–206. [Google Scholar] [CrossRef]
- Luzuriaga, J.; Polo, Y.; Pastor-Alonso, O.; Pardo-Rodriguez, B.; Larranaga, A.; Unda, F.; Sarasua, J.R.; Pineda, J.R.; Ibarretxe, G. Advances and Perspectives in Dental Pulp Stem Cell Based Neuroregeneration Therapies. Int. J. Mol. Sci. 2021, 22, 3546. [Google Scholar] [CrossRef]
- Carda, C.; Peydró, A. Ultrastructural patterns of human dentinal tubules, odontoblasts processes and nerve fibres. Tissue Cell 2006, 38, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Martínez Macal, J.C.; Olin Moreno, I. Microscopic characterization of the dentin of temporary teeth. Rev. De La Asoc. Dent. Mex. 2021, 78, 314–331. [Google Scholar] [CrossRef] [PubMed]
- Pashley, D.H. How can sensitive dentine become hypersensitive and can it be reversed? J. Dent. 2013, 41 (Suppl. S4), S49–S55. [Google Scholar] [CrossRef] [PubMed]
- Ghazali, F.B.C. Permeability of dentine. Malays. J. Med. Sci. 2003, 10, 27–36. [Google Scholar] [PubMed]
- Mjör, I.A. Dentin permeability: The basis for understanding pulp reactions and adhesive technology. Braz. Dent. J. 2009, 20, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Bleicher, F. Odontoblast physiology. Exp. Cell Res. 2014, 325, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Narayanan, K.; Ramachandran, A.; He, G.; Almushayt, A.; Evans, C.; George, A. Odontoblast cells immortalized by telomerase produce mineralized dentin-like tissue both in vitro and in vivo. J. Biol. Chem. 2002, 277, 19976–19981. [Google Scholar] [CrossRef] [PubMed]
- Ruch, J.V. Odontoblast commitment and differentiation. Biochem. Cell Biol. 1998, 76, 923–938. [Google Scholar] [CrossRef] [PubMed]
- Ruch, J.V.; Lesot, H.; Bègue-Kirn, C. Odontoblast differentiation. Int. J. Dev. Biol. 1995, 39, 51–68. [Google Scholar]
- Ching, H.S.; Luddin, N.; Rahman, I.A.; Ponnuraj, K.T. Expression of Odontogenic and Osteogenic Markers in DPSCs and SHED: A Review. Curr. Stem Cell Res. Ther. 2017, 12, 71–79. [Google Scholar] [CrossRef]
- Hanks, C.T.; Fang, D.; Sun, Z.; Edwards, C.A.; Butler, W.T. Dentin-specific proteins in MDPC-23 cell line. Eur. J. Oral Sci. 1998, 106 (Suppl. S1), 260–266. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, J.; Ghassibe-Sabbagh, M.; Fayyad-Kazan, M.; Al-Nemer, F.; Fahed, J.C.; Berberi, A.; Badran, B. Differences in osteogenic and odontogenic differentiation potential of DPSCs and SHED. J. Dent. 2020, 101, 103413. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, G.; D’Anto, V.; Cosentino, C.; Schmalz, G.; Schweikl, H.; Rengo, S. Effect of N-acetyl-L-cysteine on ROS production and cell death caused by HEMA in human primary gingival fibroblasts. Biomaterials 2006, 27, 1803–1809. [Google Scholar] [CrossRef]
- Sequeda-Castañeda, L.G.; Gutiérrez-Prieto, S.J.; Pinilla-Cruz, D.C.; Ospina-Giraldo, L.F.; Luengas-Caicedo, P.E. Use of Ilex guayusa and Piper marginatum on Oral Cavity Diseases: Remineralizacion on Enamel, Anticariogenic-Antiperiodontal-Antipathogenic Activities, Phytochemical Analysis, and Evaluation of Acute Toxicity in Mice. Pharmaceuticals 2024. Manuscript in preparation. [Google Scholar]
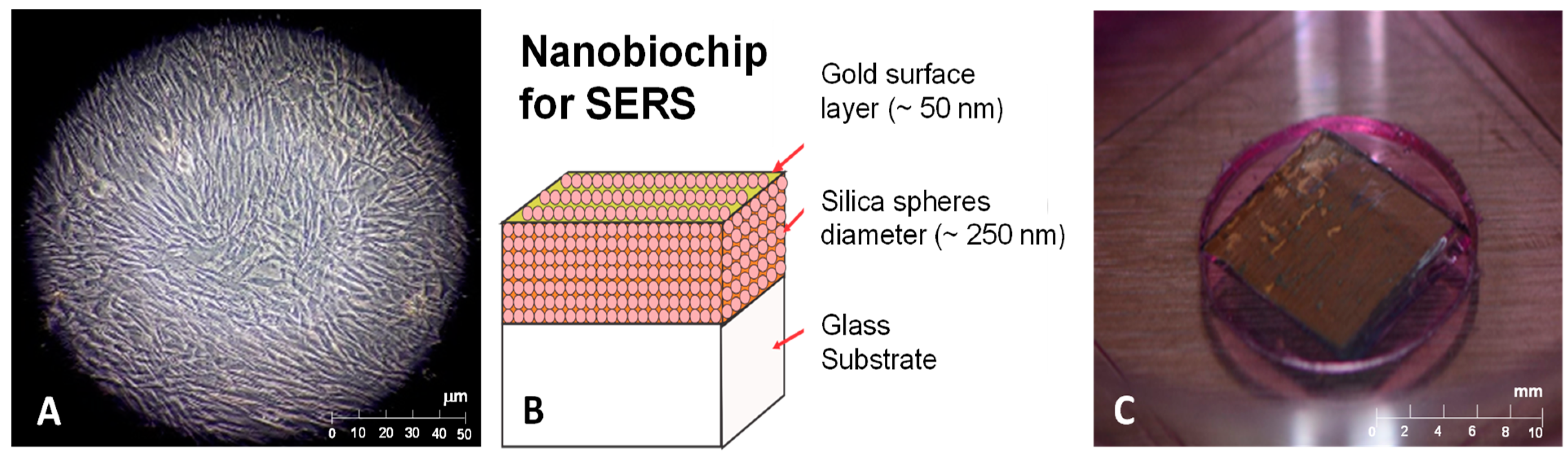


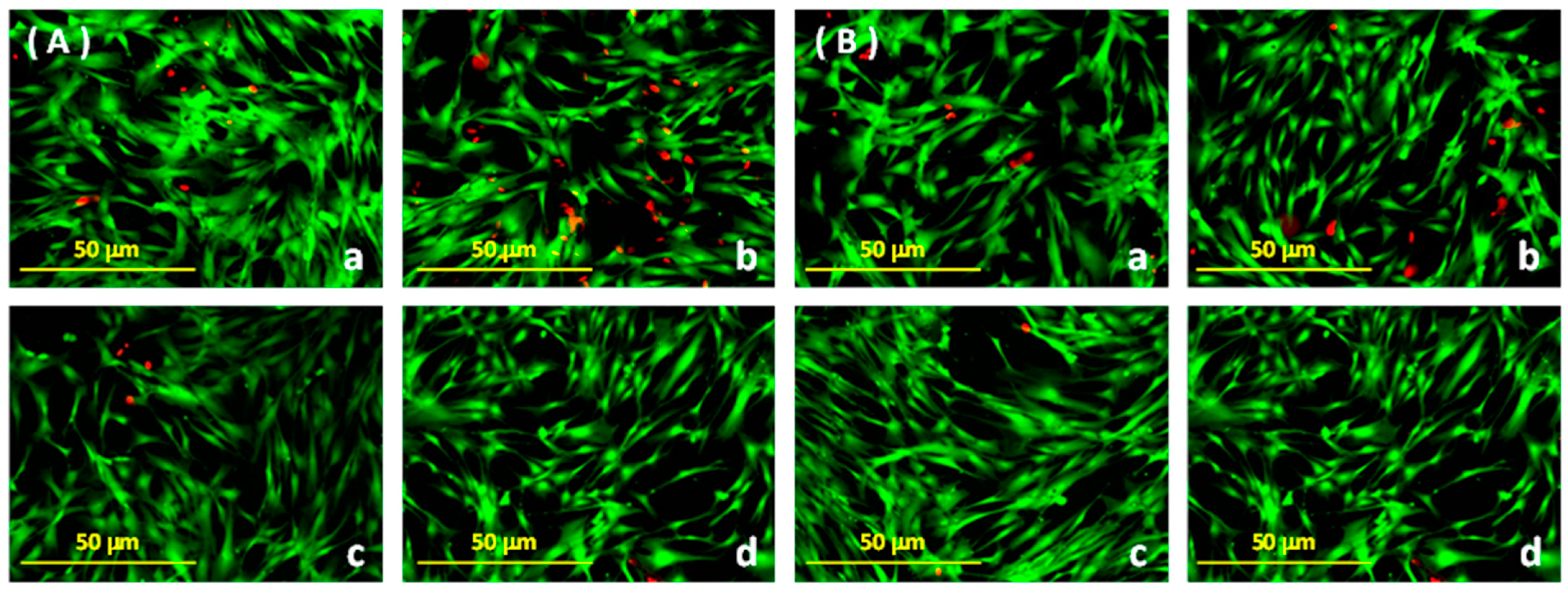

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hours | Ilex guayusa | |||
| 0 ppm * | 10 ppm | 50 ppm | 100 ppm | |
| 0 | 83.3 ± 27.6 | 87.8 ± 27.4 | 78.8 ± 31.1 | 72.7 ± 33.5 |
| 12 | 85.2 ± 21.9 | 69.2 ± 28.0 | 91.9 ± 24.2 | 92.7 ± 23.1 |
| 24 | 89.1 ± 27.5 | 91.5 ± 24.0 | 95.2 ± 22.1 | 93.8 ± 22.2 |
| Hours | Piper marginatum | |||
| 0 ppm * | 10 ppm | 50 ppm | 100 ppm | |
| 0 | 94.7 ± 27.7 | 95.8 ± 25.6 | 93.4 ± 23.2 | 92.4 ± 26.5 |
| 12 | 96.3 ± 25.6 | 90.6 ± 20.2 | 94.1 ± 21.3 | 95.2 ± 22.3 |
| 24 | 97.1 ± 21.5 | 95.2 ± 21.6 | 97.5 ± 18.7 | 96.1 ± 16.7 |
| Hours | Controls * | |||
| 0 ppm | 50 ppm | |||
| 0 | 90.4 ± 19.9 | 89.6 ± 21.8 | ||
| 12 | 88.1 ± 18.5 | 89.4 ± 19.0 | ||
| 24 | 89.8 ± 15.9 | 88.9 ± 22.1 | ||
| Hours | Ilex guayusa | |||
| 0 ppm * | 10 ppm | 50 ppm | 100 ppm | |
| 0 | 88.4 ± 21.2 | 87.3 ± 16.5 | 90.1 ± 23.2 | 89.7 ± 22.2 |
| 12 | 87.6 ± 23.5 | 83.8 ± 19.6 | 88.9 ± 18.4 | 79.9 ± 17.1 |
| 24 | 90.1 ± 17.4 | 85.6 ± 17.6 | 91.8 ± 20.0 | 91.5 ± 19.8 |
| Hours | Piper marginatum | |||
| 0 ppm * | 10 ppm | 50 ppm | 100 ppm | |
| 0 | 88.4 ± 21.2 | 88.9 ± 19.0 | 90.4 ± 15.4 | 89.6 ± 15.1 |
| 12 | 87.6 ± 23.5 | 85.6 ± 19.9 | 91.8 ± 14.5 | 91.5 ± 11.4 |
| 24 | 90.1 ± 17.4 | 92.5 ± 19.3 | 94.4 ± 20.1 | 91.7 ± 19.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sequeda-Castañeda, L.G.; Suárez-Carvajal, L.F.; Téllez-Corral, M.A.; Gutiérrez-Prieto, S.J.; Méndez-Pinzón, H.A. Evaluation of Ilex guayusa and Piper marginatum Extract Cytotoxicity on Human Dental Pulp Mesenchymal Stem Cells. Dent. J. 2024, 12, 189. https://doi.org/10.3390/dj12060189
Sequeda-Castañeda LG, Suárez-Carvajal LF, Téllez-Corral MA, Gutiérrez-Prieto SJ, Méndez-Pinzón HA. Evaluation of Ilex guayusa and Piper marginatum Extract Cytotoxicity on Human Dental Pulp Mesenchymal Stem Cells. Dentistry Journal. 2024; 12(6):189. https://doi.org/10.3390/dj12060189
Chicago/Turabian StyleSequeda-Castañeda, Luis G., Luisa F. Suárez-Carvajal, Mayra A. Téllez-Corral, Sandra J. Gutiérrez-Prieto, and Henry A. Méndez-Pinzón. 2024. "Evaluation of Ilex guayusa and Piper marginatum Extract Cytotoxicity on Human Dental Pulp Mesenchymal Stem Cells" Dentistry Journal 12, no. 6: 189. https://doi.org/10.3390/dj12060189
APA StyleSequeda-Castañeda, L. G., Suárez-Carvajal, L. F., Téllez-Corral, M. A., Gutiérrez-Prieto, S. J., & Méndez-Pinzón, H. A. (2024). Evaluation of Ilex guayusa and Piper marginatum Extract Cytotoxicity on Human Dental Pulp Mesenchymal Stem Cells. Dentistry Journal, 12(6), 189. https://doi.org/10.3390/dj12060189