
Metabolism of Phenolics of Tetrastigma hemsleyanum Roots under In Vitro Digestion and Colonic Fermentation as Well as Their In Vivo Antioxidant Activity in Rats

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Extract Preparation
2.3. In Vitro Gastrointestinal Digestion
2.3.1. Oral Digestion
2.3.2. Gastric Digestion
2.3.3. Intestinal Digestion
2.4. In Vitro Colonic Fermentation
2.4.1. Preparation of Anaerobic Medium
2.4.2. Preparation of Human Intestinal Bacterial Suspension
2.4.3. Intestinal Flora Fermentation
2.5. Qualitive and Quantitative Analysis of Phenolic Compounds
2.5.1. Qualitive Analysis by LC-QTOF-MS/MS
2.5.2. Quantitative Analysis by LC-QqQ-MS/MS
2.6. In Vivo Antioxidant Activities
2.6.1. Animals
2.6.2. Experimental Design
2.6.3. Antioxidant Assays
2.7. Statistical Analysis
3. Results and Discussion
3.1. Change in Phenolic Profiles of THR Extract during In Vitro Digestion
3.2. Change in Phenolic Profiles of THR Extract during In Vitro Colonic Fermentation
3.3. In Vivo Antioxidant Activities of THR
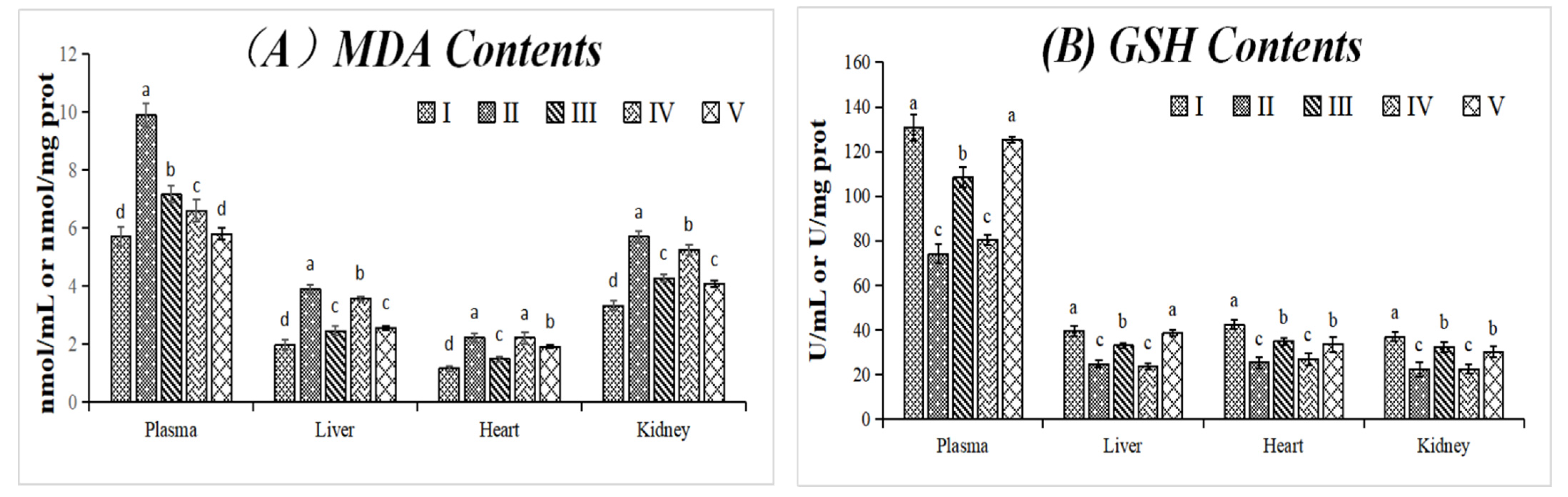
3.3.1. Effect on MDA and GSH Contents
3.3.2. Effects on SOD, GSH-Px, and T-AOC Activities
3.3.3. The Relationship between Metabolites and In Vivo Antioxidant Activities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Huang, Z.; Mao, Q.Q.; Wei, J.P. Evaluation of anti-inflammatory, analgesic and antipyretic actions for the extracts from Radix Tetrastigmae. Chin. New Drugs J. 2005, 14, 861. [Google Scholar]
- Sun, Y.; Li, H.; Hu, J.; Li, J.; Fan, Y.W.; Liu, X.R.; Deng, Z.Y. Qualitative and quantitative analysis of phenolics in Tetrastigma hemsleyanum and their antioxidant and antiproliferative activities. J. Agric. Food Chem. 2013, 61, 10507–10515. [Google Scholar] [CrossRef]
- Sun, Y.; Qin, Y.; Li, H.; Peng, H.; Chen, H.; Xie, H.R.; Deng, Z.Y. Rapid characterization of chemical constituents in Radix Tetrastigma, a functional herbal mixture, before and after metabolism and their antioxidant/antiproliferative activities. J. Funct. Foods 2015, 18, 300–318. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Yuan, Y.; Sun, Y.; Qin, Y.; Deng, Z.; Li, H. Protective effects of selenium, vitamin E, and purple carrot anthocyanins on D-galactose-induced oxidative damage in blood, liver, heart and kidney rats. Biol. Trace Elem. Res. 2016, 173, 433–442. [Google Scholar] [CrossRef]
- Ito, F.; Sono, Y.; Ito, T. Measurement and clinical significance of lipid peroxidation as a biomarker of oxidative stress: Oxidative stress in diabetes, atherosclerosis, and chronic inflammation. Antioxidants 2019, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apak, R.; Ozyurek, M.; Guclu, K.; Capanoglu, E. Antioxidant activity/capacity measurement. 3. Reactive oxygen and nitrogen species (ROS/RNS) scavenging assays, oxidative stress biomarkers, and chromatographic/chemometric assays. J. Agric. Food Chem. 2016, 64, 1046–1070. [Google Scholar] [CrossRef]
- Sun, Y.; Tsao, R.; Chen, F.; Li, H.; Wang, J.; Peng, H.; Zhang, K.; Deng, Z. The phytochemical composition, metabolites, bioavailability and in vivo antioxidant activity of Tetrastigma hemsleyanum leaves in rats. J. Funct. Foods 2017, 30, 179–193. [Google Scholar] [CrossRef]
- Samarghandian, S.; Borji, A.; Delkhosh, M.B.; Samini, F. Safranal treatment improves hyperglycemia, hyperlipidemia and oxidative stress in streptozotocin-induced diabetic rats. J. Pharm. Pharm. Sci. 2013, 16, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D. Methods for determination of antioxidant capacity: A review. Int. J. Pharm. Sci. Res. 2015, 6, 546–566. [Google Scholar]
- Gowd, V.; Bao, T.; Wang, L.; Huang, Y.; Chen, S.; Zheng, X.; Cui, S.L.; Chen, W. Antioxidant and antidiabetic activity of blackberry after gastrointestinal digestion and human gut microbiota fermentation. Food Chem. 2018, 269, 618–627. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J. Stability and metabolism of Arbutus unedo bioactive compounds (phenolics and antioxidants) under in vitro digestion and colonic fermentation. Food Chem. 2016, 201, 120–130. [Google Scholar] [CrossRef]
- Wu, B.; Kulkarni, K.; Basu, S.; Zhang, S.; Hu, M. First-Pass Metabolism via UDP-Glucuronosyltransferase: A Barrier to Oral Bioavailability of Phenolics. J. Pharm. Sci. 2011, 100, 3655–3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J.; Rubiò, L. Application of in vitro gastrointestinal digestion and colonic fermentation models to pomegranate products (juice, pulp and peel extract) to study the stability and catabolism of phenolic compounds. J. Funct. Foods 2015, 14, 529–540. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- McClements, D.J.; Li, Y. Review of in vitro digestion models for rapid screening of emulsion-based systems. Food Funct. 2010, 1, 32–59. [Google Scholar] [CrossRef]
- Maccaferri, S.; Klinder, A.; Cacciatore, S.; Chitarrari, R.; Honda, H.; Luchinat, C.; Bertini, I.; Carnevali, P.; Gibson, G.R.; Brigidi, P. In vitro fermentation of potential prebiotic flours from natural sources: Impact on the human colonic microbiota and metabolome. Mol. Nutr. Food Res. 2012, 56, 1342–1352. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Moreno Rojas, J.M.; Brindani, N.; Del Rio, D.; Lean, M.; Hara, Y.; Crozier, A. Bioavailability of Black Tea Theaflavins: Absorption, Metabolism and Colonic Catabolism. J. Agric. Food Chem. 2017, 65, 5365–5374. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.; Qian, D.; Duan, J.A.; Guo, J.; Shang, E.X.; Shu, Y.; Xue, C. UPLC–QTOF/MS-based screening and identification of the constituents and their metabolites in rat plasma and urine after oral administration of Glechoma longituba extract. J. Chromatogr. B 2010, 878, 2741–2750. [Google Scholar] [CrossRef]
- Ekbatan, S.S.; Sleno, L.; Sabally, K.; Khairallah, J.; Azadi, B.; Rodes, L.; Prakash, S.J.; Donnelly, D.; Kubow, S. Biotransformation of polyphenols in a dynamic multistage gastrointestinal model. Food Chem. 2016, 204, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Bahloul, N.; Bellili, S.; Aazza, S.; Chérif, A.; Faleiro, M.L.; Antunes, M.D.; Miguel, M.G.; Mnif, W. Aqueous extracts from tunisian diplotaxis: Phenol content, antioxidant and anti-acetylcholinesterase activities, and impact of exposure to simulated gastrointestinal fluids. Antioxidants 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef]
- Crespy, V.; Morand, C.; Manach, C.; Besson, C.; Demigne, C.; Remesy, C. Part of quercetin absorbed in the small intestine is conjugated and further secreted in the intestinal lumen. Am. J. Physiol.-Gastrointest. Liver Physiol. 1999, 277, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Luzardo-Ocampo, I.; Campos-Vega, R.; Gaytán-Martínez, M.; Preciado-Ortiz, R.; Mendoza, S.; Loarca-Piña, G. Bioaccessibility and antioxidant activity of free phenolic compounds and oligosaccharides from corn (Zea mays L.) and common bean (Phaseolus vulgaris L.) chips during in vitro gastrointestinal digestion and simulated colonic fermentation. Food Res. Int. 2017, 100, 304–311. [Google Scholar] [CrossRef]
- Gayoso, L.; Claerbout, A.S.; Calvo, M.I.; Cavero, R.Y.; Astiasarán, I.; Ansorena, D. Bioaccessibility of rutin, caffeic acid and rosmarinic acid: Influence of the in vitro gastrointestinal digestion models. J. Funct. Foods 2016, 26, 428–438. [Google Scholar] [CrossRef]
- Attri, S.; Sharma, K.; Raigond, P.; Goel, G. Colonic fermentation of polyphenolics from Sea buckthorn (Hippophae rhamnoides) berries: Assessment of effects on microbial diversity by Principal Component Analysis. Food Res. Int. 2018, 105, 324–332. [Google Scholar] [CrossRef]
- Chang, Q.; Zuo, Z.; Chow, M.S.; Ho, W.K. Difference in absorption of the two structurally similar flavonoid glycosides, hyperoside and isoquercitrin, in rats. Eur. J. Pharm. Biopharm. 2005, 59, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Takahama, U.; Hirota, S. Effects of starch on nitrous acid-induced oxidation of kaempferol and inhibition of α-amylase-catalysed digestion of starch by kaempferol under conditions simulating the stomach and the intestine. Food Chem. 2013, 141, 313–319. [Google Scholar] [CrossRef]
- Gião, M.S.; Gomes, S.; Madureira, A.R.; Faria, A.; Pestana, D.; Calhau, C.; Pintado, M.E.; Azevedo, I.; Malcata, F.X. Effect of in vitro digestion upon the antioxidant capacity of aqueous extracts of Agrimonia eupatoria, Rubus idaeus, Salvia sp. and Satureja montana. Food Chem. 2012, 131, 761–767. [Google Scholar] [CrossRef]
- McGhie, T.K.; Walton, M.C. The bioavailability and absorption of anthocyanins: Towards a better understanding. Mol. Nutr. Food Res. 2007, 51, 702–713. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Moon, J.S.; Shin, S.Y.; Li, L.; Lee, Y.B.; Kim, T.J.; Han, N.S. Isolation and characterization of human intestinal Enterococcus avium EFEL009 converting rutin to quercetin. Lett. Appl. Microbiol. 2016, 62, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Shi, D.Z.; Wang, T.Y.; Zheng, S.Q.; Liu, L.J.; Sun, Z.X.; Wang, T.Y.; Ding, Y. Transformation of trollioside and isoquercetin by human intestinal flora in vitro. Chin. J. Nat. Med. 2016, 14, 220–226. [Google Scholar] [CrossRef]
- Vollmer, M.; Esders, S.; Farquharson, F.M.; Neugart, S.; Duncan, S.H.; Schreiner, M.; Louis, P.; Maul, R.; Rohn, S. Mutual interaction of phenolic compounds and microbiota: Metabolism of complex phenolic apigenin-c-and kaempferol-o-derivatives by human fecal samples. J. Agric. Food Chem. 2018, 66, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafay, S.; Morand, C.; Manach, C.; Besson, C.; Scalbert, A. Absorption and metabolism of caffeic acid and chlorogenic acid in the small intestine of rats. Br. J. Nutr. 2006, 96, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, I.A.; de Peña, M.; Cid, C.; Crozier, A. Catabolism of coffee chlorogenic acids by human colonic microbiota. Biofactors 2013, 39, 623–632. [Google Scholar] [CrossRef]
- Sadeghi Ekbatan, S.; Iskandar, M.M.; Sleno, L.; Sabally, K.; Khairallah, J.; Prakash, S.; Kubow, S. Absorption and metabolism of phenolics from digests of polyphenol-rich potato extracts using the Caco-2/HepG2 co-culture system. Foods 2018, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, T.; Wang, Y.; Li, Y.T.; Gowd, V.; Niu, X.H.; Yang, H.Y.; Chen, L.S.; Chen, W.; Sun, C.D. Antioxidant and antidiabetic properties of tartary buckwheat rice flavonoids after in vitro digestion. J. Zhejiang Univ. Sci. B 2016, 17, 941–951. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Phenolic Content (mg GAE/g DW) B | Total Flavonoid Content (mg CAE/g DW) C |
|---|---|---|
| Raw | 225.38 ± 2.62 h | 124.95 ± 3.31 g |
| In vitro gastrointestinal digestion | ||
| Oral digestion | 214.13 ± 2.34 g | 109.47 ± 1.78 f |
| Gastric digestion | 104.61 ± 1.51 f | 83.64 ± 2.14 e |
| Intestinal digestion | 97.53 ± 2.47 e | 81.98 ± 1.37 e |
| In vitro colonic fermentation by human microflora | ||
| 0 h | 86.93 ± 2.09 d | 72.44 ± 1.56 d |
| 12 h | 78.27 ± 1.88 c | 61.25 ± 1.08 c |
| 24 h | 65.42 ± 3.53 b | 49.84 ± 1.67 b |
| 48 h | 33.85 ± 2.05 a | 17.56 ± 1.29 a |
| No. | Compounds B | tR (min) | Parent/Daughter Ions (m/z) | Raw | Gastrointestinal Digestion | Colonic Fermentation by Human Microflora | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Oral | Gastric | Intestinal | 0 h | 12 h | 24 h | 48 h | |||||
| 1 | Quercetin-3-rutinoside | 27.41 | 609/301 | 16.86 ± 1.19 h | 14.09 ± 1.25 g | 8.77 ± 0.86 f | 8.04 ± 0.45 d | 8.12 ± 0.79 e | 6.70 ± 0.26 c | 3.37 ± 0.03 b | 3.19 ± 0.40 a |
| 2 | Quercetin-3-glucoside | 29.18 | 463/301 | 12.55 ± 1.03 h | 11.24 ± 1.31 g | 7.18 ± 1.12 f | 6.63 ± 0.58 e | 6.57 ± 0.58 d | 4.92 ± 0.68 c | 2.48 ± 0.11 b | 2.42 ± 0.21 a |
| 3 | Kaempferol-3-rutinoside | 30.39 | 593/285 | 53.48 ± 3.47 h | 49.13 ± 2.76 g | 27.94 ± 1.34 f | 27.88 ± 1.07 e | 24.5 ± 2.01 d | 21.98 ± 1.91 c | 21.46 ± 1.31 b | 7.67 ± 0.55 a |
| 4 | Kaempferol-3-glucoside | 31.26 | 447/285 | 19.97 ± 1.57 h | 16.40 ± 1.44 g | 9.54 ± 0.95 e | 9.73 ± 1.04 f | 9.38 ± 0.82 d | 7.60 ± 0.64 c | 4.54 ± 0.08 b | 2.88 ± 0.09 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Guo, F.; Peng, X.; Cheng, K.; Xiao, L.; Zhang, H.; Li, H.; Jiang, L.; Deng, Z. Metabolism of Phenolics of Tetrastigma hemsleyanum Roots under In Vitro Digestion and Colonic Fermentation as Well as Their In Vivo Antioxidant Activity in Rats. Foods 2021, 10, 2123. https://doi.org/10.3390/foods10092123
Sun Y, Guo F, Peng X, Cheng K, Xiao L, Zhang H, Li H, Jiang L, Deng Z. Metabolism of Phenolics of Tetrastigma hemsleyanum Roots under In Vitro Digestion and Colonic Fermentation as Well as Their In Vivo Antioxidant Activity in Rats. Foods. 2021; 10(9):2123. https://doi.org/10.3390/foods10092123
Chicago/Turabian StyleSun, Yong, Fanghua Guo, Xin Peng, Kejun Cheng, Lu Xiao, Hua Zhang, Hongyan Li, Li Jiang, and Zeyuan Deng. 2021. "Metabolism of Phenolics of Tetrastigma hemsleyanum Roots under In Vitro Digestion and Colonic Fermentation as Well as Their In Vivo Antioxidant Activity in Rats" Foods 10, no. 9: 2123. https://doi.org/10.3390/foods10092123
APA StyleSun, Y., Guo, F., Peng, X., Cheng, K., Xiao, L., Zhang, H., Li, H., Jiang, L., & Deng, Z. (2021). Metabolism of Phenolics of Tetrastigma hemsleyanum Roots under In Vitro Digestion and Colonic Fermentation as Well as Their In Vivo Antioxidant Activity in Rats. Foods, 10(9), 2123. https://doi.org/10.3390/foods10092123