
Comparison of the Probiotic Potential between Lactiplantibacillus plantarum Isolated from Kimchi and Standard Probiotic Strains Isolated from Different Sources


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Cell Culture
2.3. Selection of Bacteria Showing Anti-Inflammatory Activity
2.4. Identification of the Strains
2.5. Tolerance of GI Environment and Heat
2.6. Antibacterial Activity
2.7. Antioxidant Activity
2.8. Adhesion Ability to Intestinal Epithelial Cells
2.9. Anti-Inflammatory Activity
2.10. Antibiotic Susceptibility
2.11. Statistical Analysis
3. Results and Discussion
3.1. Selection of Bacteria with Anti-Inflammatory Activity
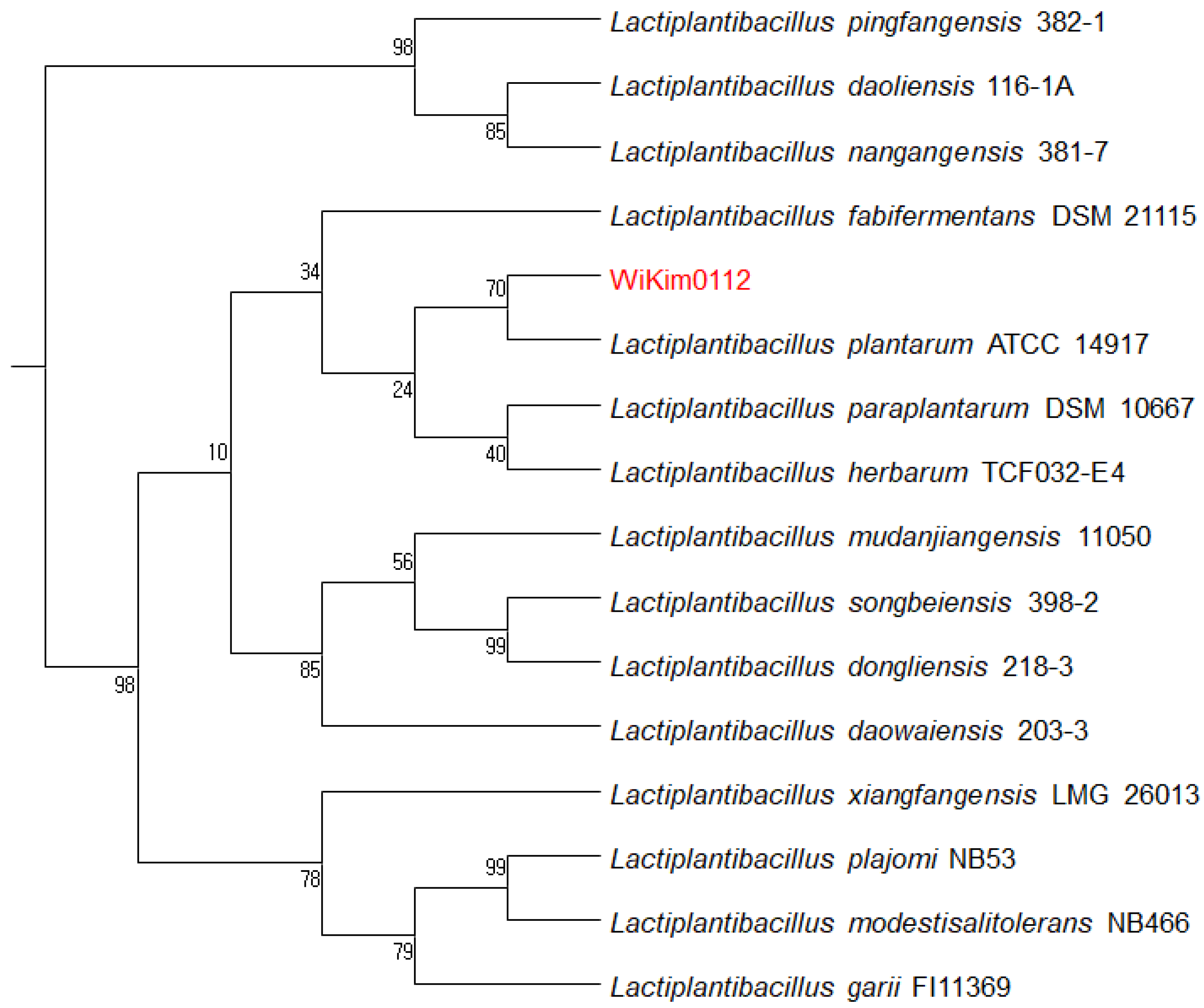
3.2. Identification of WiKim0112
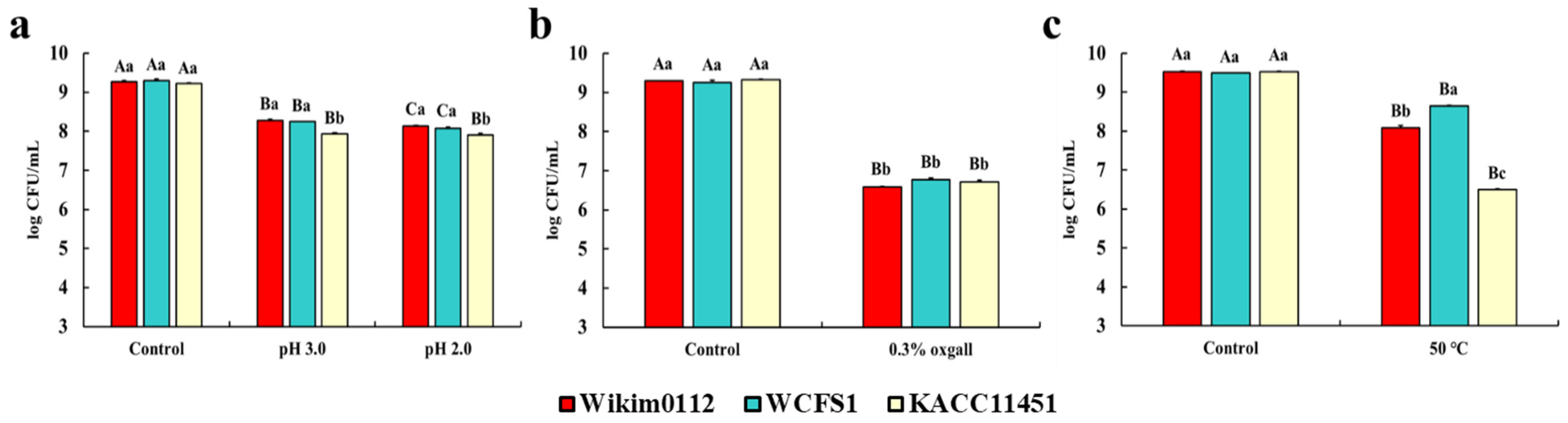
3.3. Viability of Lpb. plantarum Strains to GI Environment and Heat Treatment
3.4. Antibacterial Activity of Lpb. plantarum Strains
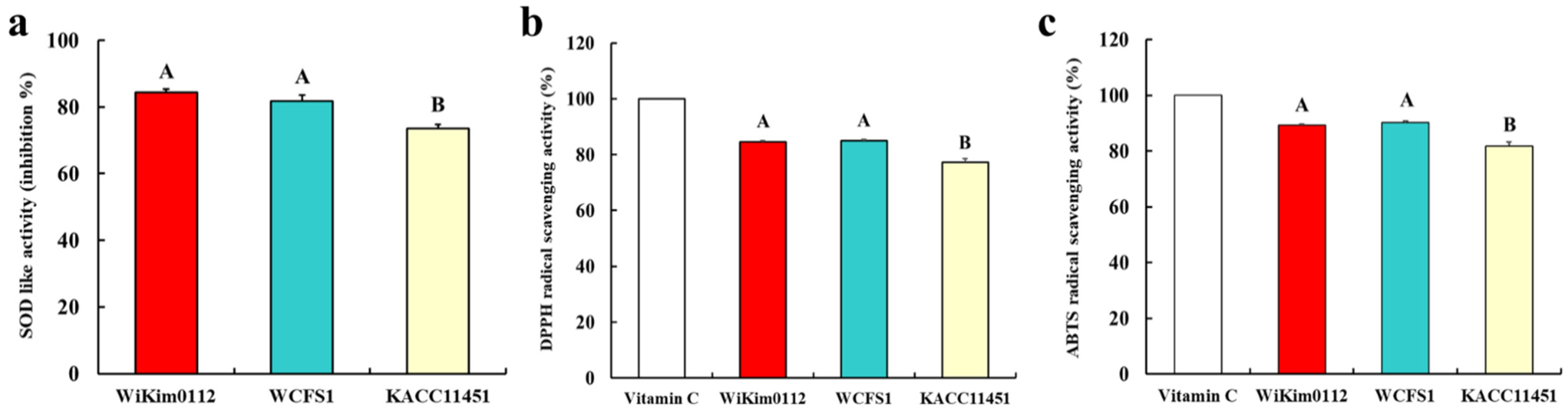
3.5. Antioxidant Activity of Lpb. plantarum Strains
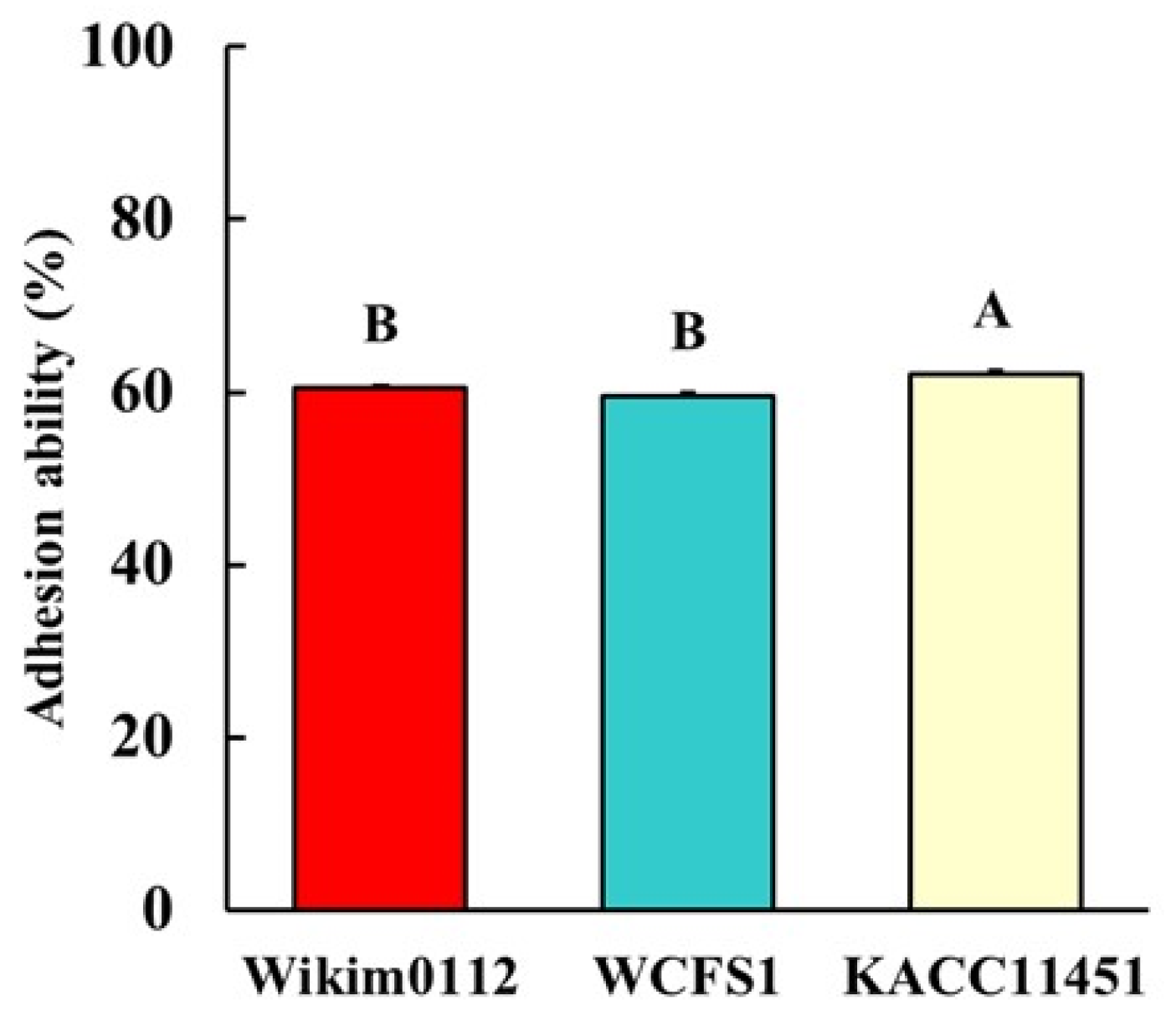
3.6. Adhesion Ability of Lpb. plantarum Strains to Intestinal Epithelial Cells
3.7. Anti-Inflammatory Activity of Lpb. plantarum Strains in Intestinal Epithelial Cells
3.8. Antibiotic Susceptibility of Lpb. plantarum Strains
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Kimchi microflora: History, current status, and perspectives for industrial kimchi production. Appl. Microbiol. Biotechnol. 2014, 98, 2385–2393. [Google Scholar] [CrossRef]
- Chang, J.H.; Shim, Y.; Cha, S.K.; Chee, K. Probiotic characteristics of lactic acid bacteria isolated from kimchi. J. Appl. Microbiol. 2010, 109, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-T.; Yang, S.J.; Paik, H.-D. Probiotic properties of novel probiotic Levilactobacillus brevis KU15147 isolated from radish kimchi and its antioxidant and immune-enhancing activities. Food Sci. Biotechnol. 2021, 30, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-promoting role of lactiplantibacillus plantarum isolated from fermented foods. Microorganisms 2021, 9, 349. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus plantarum with functional properties: An approach to increase safety and shelf-life of fermented foods. BioMed Res. Int. 2018, 2018, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atrih, A.; Rekhif, N.; Milliere, J.; Lefebvre, G. Detection and characterization of a bacteriocin produced by Lactobacillus plantarum C19. Can. J. Microbiol. 1993, 39, 1173–1179. [Google Scholar] [CrossRef]
- Wen, L.S.; Philip, K.; Ajam, N. Purification, characterization and mode of action of plantaricin K25 produced by Lactobacillus plantarum. Food Control 2016, 60, 430–439. [Google Scholar] [CrossRef]
- Hu, M.; Zhao, H.; Zhang, C.; Yu, J.; Lu, Z. Purification and characterization of plantaricin 163, a novel bacteriocin produced by Lactobacillus plantarum 163 isolated from traditional Chinese fermented vegetables. J. Agric. Food Chem. 2013, 61, 11676–11682. [Google Scholar] [CrossRef]
- Gonzalez, B.; Arca, P.; Mayo, B.; Suárez, J.E. Detection, purification, and partial characterization of plantaricin C, a bacteriocin produced by a Lactobacillus plantarum strain of dairy origin. Appl. Environ. Microbiol. 1994, 60, 2158–2163. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhao, Y.; Sun, Y.; Gu, Q. Purification and characterisation of plantaricin ZJ008, a novel bacteriocin against Staphylococcus spp. from Lactobacillus plantarum ZJ008. Food Chem. 2014, 165, 216–223. [Google Scholar] [CrossRef]
- Todorov, S.D.; Perin, L.M.; Carneiro, B.M.; Rahal, P.; Holzapfel, W.; Nero, L.A. Safety of Lactobacillus plantarum ST8Sh and its bacteriocin. Probiotics Antimicrob. Proteins 2017, 9, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Rekhif, N.; Atrih, A.; Lefebvrexy, G. Activity of plantaricin SA6, a bacteriocin produced by Lactobacillus plantarum SA6 isolated from fermented sausage. J. Appl. Bacteriol. 1995, 78, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Dixit, G.; Samarth, D.; Tale, V.; Bhadekar, R. Comparative studies on potential probiotic characteristics of Lactobacillus acidophilus strains. EurAsian J. BioSci. 2013, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Wang, J.; Yan, L.; Chen, W.; Liu, X.-M.; Zhang, H.-P. In vitro comparison of probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected probiotic strains. LWT Food Sci. Technol. 2009, 42, 1640–1646. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Evans, C.; Adams, M.; Baines, S. In vitro analysis of gastrointestinal tolerance and intestinal cell adhesion of probiotics in goat’s milk ice cream and yogurt. Food Res. Int. 2012, 49, 619–625. [Google Scholar] [CrossRef]
- Jeong, C.H.; Ryu, H.; Zhang, T.; Lee, C.H.; Seo, H.G.; Han, S.G. Green tea powder supplementation enhances fermentation and antioxidant activity of set-type yogurt. Food Sci. Biotechnol. 2018, 27, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jeong, C.H.; Cheng, W.N.; Bae, H.; Seo, H.G.; Petriello, M.C.; Han, S.G. Moringa extract enhances the fermentative, textural, and bioactive properties of yogurt. LWT 2019, 101, 276–284. [Google Scholar] [CrossRef]
- Cheng, W.; Jeong, C.; Kim, D.; Han, S. Effects of moringa extract on adhesion and invasion of Escherichia coli O55 in bovine mammary epithelial cells. J. Dairy Sci. 2020, 103, 7416–7424. [Google Scholar] [CrossRef]
- Won, S.-M.; Chen, S.; Park, K.W.; Yoon, J.-H. Isolation of lactic acid bacteria from kimchi and screening of Lactobacillus sakei ADM14 with anti-adipogenic effect and potential probiotic properties. LWT 2020, 126, 109296. [Google Scholar] [CrossRef]
- Cockerill, F.R.; Wikler, M.A.; Alder, J.; Dudley, M.; Eliopoulos, G.; Ferraro, M.; Hardy, D.; Hecht, D.; Hindler, J.; Patel, J. Performance standards for antimicrobial susceptibility testing: Twenty-second informational supplement. Clin. Lab. Stand. Inst. 2012, 32, M100–S122. [Google Scholar]
- Angmo, K.; Kumari, A.; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LW Food Sci. Technol. 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Lin, P.-R.; Ng, C.-C.; Shyu, Y.-T. Probiotic properties of Lactobacillus strains isolated from the feces of breast-fed infants and Taiwanese pickled cabbage. Anaerobe 2010, 16, 578–585. [Google Scholar] [CrossRef]
- Conway, P.; Gorbach, S.; Goldin, B. Survival of lactic acid bacteria in the human stomach and adhesion to intestinal cells. J. Dairy Sci. 1987, 70, 1–12. [Google Scholar] [CrossRef]
- Huang, R.; Tao, X.; Wan, C.; Li, S.; Xu, H.; Xu, F.; Shah, N.P.; Wei, H. In vitro probiotic characteristics of Lactobacillus plantarum ZDY 2013 and its modulatory effect on gut microbiota of mice. J. Dairy Sci. 2015, 98, 5850–5861. [Google Scholar] [CrossRef]
- Hong, S.-I.; Kim, Y.-J.; Pyun, Y.-R. Acid Tolerance ofLactobacillus plantarumfromKimchi. LWT Food Sci. Technol. 1999, 32, 142–148. [Google Scholar] [CrossRef]
- Nannen, N.L.; Hutkins, R.W. Proton-translocating adenosine triphosphatase activity in lactic acid bacterial. J. Dairy Sci. 1991, 74, 747–751. [Google Scholar] [CrossRef]
- Ramirez-Chavarin, M.; Wacher, C.; Eslava-Campos, C.; Perez-Chabela, M. Probiotic potential of thermotolerant lactic acid bacteria strains isolated from cooked meat products. Int. Food Res. J. 2013, 20, 991–1000. [Google Scholar]
- Vataščinová, T.; Pipova, M.; Fraqueza, M.; Maľa, P.; Dudrikova, E.; Drážovská, M.; Laukova, A. Antimicrobial potential of Lactobacillus plantarum strains isolated from Slovak raw sheep milk cheeses. J. Dairy Sci. 2020, 103, 6900–6903. [Google Scholar] [CrossRef]
- Lin, T.-H.; Pan, T.-M. Characterization of an antimicrobial substance produced by Lactobacillus plantarum NTU 102. J. Microbiol. Immunol. Infect. 2019, 52, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Kothari, D.; Lee, W.-D.; Jung, E.S.; Niu, K.-M.; Lee, C.H.; Kim, S.-K. Controlled fermentation using autochthonous Lactobacillus plantarum improves antimicrobial potential of Chinese chives against poultry pathogens. Antibiotics 2020, 9, 386. [Google Scholar] [CrossRef]
- Piard, J.; Desmazeaud, M. Inhibiting factors produced by lactic acid bacteria. 2. Bacteriocins and other antibacterial substances. Lait 1992, 72, 113–142. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Vesterlund, S. Antimicrobial components from lactic acid bacteria. Food Sci. Technol. N. Y. Marcel Dekker 2004, 139, 375–396. [Google Scholar]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant properties of probiotic bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Song, M.W.; Lee, N.-K.; Paik, H.-D. Antioxidant effects of live and heat-killed probiotic Lactobacillus plantarum Ln1 isolated from kimchi. J. Food Sci. Technol. 2018, 55, 3174–3180. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Salminen, S.; Isolauri, E. Probiotics: An overview of beneficial effects. In Lactic Acid Bacteria: Genetics, Metabolism and Applications; Siezen, R.J., Kok, J., Abeem, T., Schaafsma, G., Eds.; Springer: Amsterdam, The Netherlands, 2002; pp. 279–289. [Google Scholar]
- De Palencia, P.F.; López, P.; Corbí, A.L.; Peláez, C.; Requena, T. Probiotic strains: Survival under simulated gastrointestinal conditions, in vitro adhesion to Caco-2 cells and effect on cytokine secretion. Eur. Food Res. Technol. 2008, 227, 1475–1484. [Google Scholar] [CrossRef] [Green Version]
- Sohn, H.; Chang, Y.H.; Yune, J.H.; Jeong, C.H.; Shin, D.-M.; Kwon, H.C.; Kim, D.H.; Hong, S.W.; Hwang, H.; Jeong, J.Y. Probiotic Properties of Lactiplantibacillus plantarum LB5 Isolated from Kimchi Based on Nitrate Reducing Capability. Foods 2020, 9, 1777. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Provencio, D.; Llopis, M.; Antolín, M.; De Torres, I.; Guarner, F.; Pérez-Martínez, G.; Monedero, V. Adhesion properties of Lactobacillus casei strains to resected intestinal fragments and components of the extracellular matrix. Arch. Microbiol. 2009, 191, 153–161. [Google Scholar] [CrossRef]
- Pretzer, G.; Snel, J.; Molenaar, D.; Wiersma, A.; Bron, P.A.; Lambert, J.; de Vos, W.M.; van der Meer, R.; Smits, M.A.; Kleerebezem, M. Biodiversity-based identification and functional characterization of the mannose-specific adhesin of Lactobacillus plantarum. J. Bacteriol. 2005, 187, 6128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahfoufi, N.; Mallet, J.; Graham, E.; Matar, C. Role of probiotics and prebiotics in immunomodulation. Curr. Opin. Food Sci. 2018, 20, 82–91. [Google Scholar] [CrossRef]
- Park, J.-S.; Joe, I.; Rhee, P.D.; Jeong, C.-S.; Jeong, G. A lactic acid bacterium isolated from kimchi ameliorates intestinal inflammation in DSS-induced colitis. J. Microbiol. 2017, 55, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Pillidge, C.; Gopal, P.; Gill, H. Antibiotic susceptibility profiles of new probiotic Lactobacillus and Bifidobacterium strains. Int. J. Food Microbiol. 2005, 98, 211–217. [Google Scholar] [CrossRef]
- Mathur, S.; Singh, R. Antibiotic resistance in food lactic acid bacteria—A review. Int. J. Food Microbiol. 2005, 105, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Sánchez, B.; de Los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′–3′) |
|---|---|
| TNF-α a | (F) AAG CCC TGG TAT GAG CCC ATC TAT(R) AGG GCA ATG ATC CCA AAG TAG ACC |
| IL-6 | (F) ACA GCC ACT CAC CTC TTC AGA AC(R) TTT TCT GCC AGT GCC TCT TTG C |
| IL-1β | (F) TGT ACC TGT CCT GCG TGT TGA AAG(R) CTG GGC AGA CTC AAA TTC CAG CTT |
| IL-4 | (F) TCA TTT TCC CTC GGT TTC AG(R) AGA ACA GAG GGG GAA GCA GT |
| IL-10 | (F) TCA GGG TGG CGA CTC TAT(R) TGG GCT TCT TCT AAA TCG TTC |
| IFN-γ | (F) ATA TCT TGG CTT TTC AGC TC(R) CTC CTT TTT CGC TTC CCT GT |
| GAPDH | (F) GAC CCC TTC ATT GAC CTC AAC TAC(R) ATG ACA AGC TTC CCG TTC TCA G |
| Pathogens | Inhibition Zone (mm) ± SD a | ||
|---|---|---|---|
| WiKim0112 | WCFS1 | KACC11451 | |
| E. coli O157:H7 (ATCC 35150) | 18.0 ± 0.5 | 19.0 ± 0.5 | 16.0 ± 0.1 |
| Listeria monocytogenes (ATCC 15313) | 20.0 ± 0.4 | 18.0 ± 0.2 | 15.0 ± 0.5 |
| Salmonella choleraesuis (KCCM40763) | 20.0 ± 0.3 | 20.0 ± 0.1 | 16.0 ± 0.2 |
| Shigella boydii (KCCM41649) | 23.0 ± 0.4 | 24.0 ± 0.2 | 21.0 ± 0.2 |
| Staphylococcus aureus (ATCC25923) | 23.0 ± 0.3 | 23.0 ± 0.1 | 20.0 ± 0.3 |
| Yersinia enterocolitica (KCCM41657) | 23.0 ± 0.2 | 24.0 ± 0.5 | 22.0 ± 0.5 |
| Antibiotics | Wikim0112 | WCFS1 | KACC11451 | |||
|---|---|---|---|---|---|---|
| Zone Diameter (mm) ± SD a | Sensitivity | Zone Diameter (mm) ± SD | Sensitivity | Zone Diameter (mm) ± SD | Sensitivity | |
| Ampicillin (AMP) | 29.0 ± 1.4 | S | 39.3 ± 1.3 | S | 29.7 ± 1.3 | S |
| Chloramphenicol (C) | 31.0 ± 0.8 | S | 30.7 ± 0.9 | S | 30.7 ± 0.9 | S |
| Clindamycin (CD) | 26.3 ± 1.3 | S | 31.7 ± 1.3 | S | 12.0 ± 1.6 | R |
| Ciprofloxacin (CIP) | 14.3 ± 0.9 | I | 13.0 ± 1.4 | R | 13.7 ± 0.5 | R |
| Gentamicin (CN) | 18.7 ± 1.3 | I | 28.7 ± 0.9 | S | 28.3 ± 1.3 | S |
| Doxycycline (DXT) | 28.7 ± 0.9 | S | 16.3 ± 1.3 | I | 18.7 ± 0.9 | I |
| Erythromycin (E) | 37.0 ± 0.8 | S | 38.0 ± 1.6 | S | 33.7 ± 1.3 | S |
| Kanamycin (K) | 12.3 ± 1.3 | S | 31.3 ± 0.5 | S | 28.3 ± 0.5 | S |
| Penicillin G (P) | 29.3 ± 0.9 | S | 31.3 ± 1.3 | S | 24.7 ± 0.5 | S |
| Streptomycin (S) | 13.3 ± 1.3 | R | 25.3 ± 1.3 | S | 23.0 ± 1.4 | S |
| Trimethoprim-sulfamethoxazole (SXT) | 29.3 ± 1.6 | S | 26.7 ± 0.5 | S | 29.7 ± 1.4 | S |
| Tetracycline (TE) | 18.7 ± 1.3 | I | 12.0 ± 1.3 | R | - | R |
| Vancomycin (VA) | - b | R | - | R | - | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, C.-H.; Sohn, H.; Hwang, H.; Lee, H.-J.; Kim, T.-W.; Kim, D.-S.; Kim, C.-S.; Han, S.-G.; Hong, S.-W. Comparison of the Probiotic Potential between Lactiplantibacillus plantarum Isolated from Kimchi and Standard Probiotic Strains Isolated from Different Sources. Foods 2021, 10, 2125. https://doi.org/10.3390/foods10092125
Jeong C-H, Sohn H, Hwang H, Lee H-J, Kim T-W, Kim D-S, Kim C-S, Han S-G, Hong S-W. Comparison of the Probiotic Potential between Lactiplantibacillus plantarum Isolated from Kimchi and Standard Probiotic Strains Isolated from Different Sources. Foods. 2021; 10(9):2125. https://doi.org/10.3390/foods10092125
Chicago/Turabian StyleJeong, Chang-Hee, Hyejin Sohn, Hyelyeon Hwang, Ho-Jae Lee, Tae-Woon Kim, Dong-Sub Kim, Chun-Sung Kim, Sung-Gu Han, and Sung-Wook Hong. 2021. "Comparison of the Probiotic Potential between Lactiplantibacillus plantarum Isolated from Kimchi and Standard Probiotic Strains Isolated from Different Sources" Foods 10, no. 9: 2125. https://doi.org/10.3390/foods10092125
APA StyleJeong, C.-H., Sohn, H., Hwang, H., Lee, H.-J., Kim, T.-W., Kim, D.-S., Kim, C.-S., Han, S.-G., & Hong, S.-W. (2021). Comparison of the Probiotic Potential between Lactiplantibacillus plantarum Isolated from Kimchi and Standard Probiotic Strains Isolated from Different Sources. Foods, 10(9), 2125. https://doi.org/10.3390/foods10092125