

Antibacterial Activity and Mechanism of Linalool against Shigella sonnei and Its Application in Lettuce
 and
and
Abstract
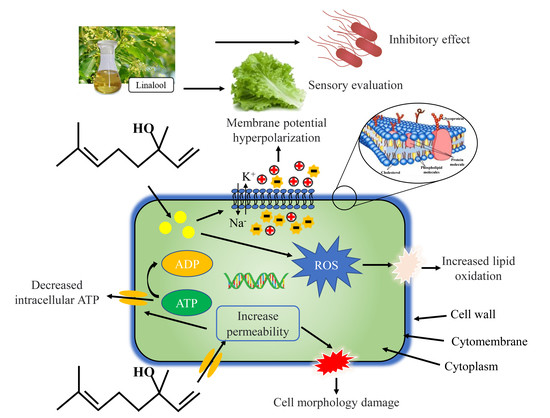
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Bacterial Strains and Culture Conditions
2.3. Effects of Linalool on the Inhibitory Effect of S. sonnei
2.3.1. Determination of MIC and Minimum Bactericidal Concentration (MBC)
2.3.2. Growth Curves
2.3.3. Inactivation Effect of Linalool on S. sonnei in LB Broth and Phosphate-Buffered Sa-Line (PBS)
2.4. Antibacterial Mechanism of Linalool on S. sonnei
2.4.1. Intracellular ROS Levels
2.4.2. Intracellular ATP Concentrations
2.4.3. Extracellular MDA Content
2.4.4. Field-Emission Scanning Electron Microscopy (FESEM)-Based Observations
2.4.5. Membrane Potential
2.5. Effects of Linalool on Lettuce Leaves
2.5.1. Inactivation Effect of Linalool on S. sonnei on Leaf Surface of Lettuce
2.5.2. Superficial Color
2.5.3. Organoleptic Quality: Overall Visual Quality (OVQ)
2.6. Statistical Analysis
3. Results
3.1. Effects of Linalool on the Inhibitory Effect of S. sonnei
3.1.1. MIC and MBC
3.1.2. Growth Curve
3.1.3. Antibacterial Curve Assay
3.2. Antibacterial Mechanism of Linalool on S. sonnei
3.2.1. Intracellular ROS Level
3.2.2. Intracellular ATP Content
3.2.3. Extracellular Malondialdehyde (MDA) Content
3.2.4. FESEM Observations
3.2.5. Membrane Potential
3.3. Effects of Linalool on Lettuce Leaves
3.3.1. Inactivation Effect of Linalool on S. sonnei on the Leaf Surface of Lettuce
3.3.2. Surface Color
3.3.3. Organoleptic Quality: Overall Visual Quality (OVQ)
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lee, J.-Y.; Kang, D.-H. Development of an improved selective medium for the detection of Shigella spp. LWT-Food Sci Technol. 2016, 65, 311–317. [Google Scholar] [CrossRef]
- Khalil, I.A.; Troeger, T.; Blacker, B.F. Morbidity and mortality due to shigella and enterotoxigenic Escherichia coli diarrhoea: The global burden of disease study. Lancet Infect. Dis. 2018, 18, 1305. [Google Scholar] [CrossRef]
- Naidoo, S.; Olaniran, A. Treated wastewater effluent as a source of microbial pollution of surface water resources. Int. J. Environ. Res. Public Health 2014, 11, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Torraca, V.; Holt, K.; Mostowy, S. Shigella sonnei. Trends Microbiol. 2020, 28, 696–697. [Google Scholar] [CrossRef]
- Wang, J.; Qiu, S.; Xu, X.; Su, W.; Liang, B.; Ma, Q. Emergence of ONPG-negative Shigella sonnei in Shanghai, China. Diagn. Microbiol. Infect. Dis. 2015, 83, 338–340. [Google Scholar] [CrossRef]
- Chourabi, K.; Campoy, S.; Rodriguez, J.; Kloula, S.; Landoulsi, A.; Chatti, A. UV-C adaptation of Shigella: Morphological, outer membrane proteins, secreted proteins, and lipopolysaccharides effects. Curr. Microbiol. 2017, 74, 1261–1269. [Google Scholar] [CrossRef]
- Mecha, A.C.; Onyango, M.S.; Ochieng, A.; Momba, M.N.B. Modelling inactivation kinetics of waterborne pathogens in municipal wastewater using ozone. Environ. Eng. Res. 2020, 25, 890–897. [Google Scholar] [CrossRef]
- Rangel-Vargas, E.; Luna-Rojo, A.M.; Cadena-Ramirez, A.; Torres-Vitela, M.R.; Gomez-Aldapa, C.A. Behavior of 11 foodborne bacteria on whole and cut mangoes var. ataulfo and kent and antibacterial activities of hibiscus sabdariffa extracts and chemical sanitizers directly onto mangoes contaminated with foodborne bacteria. J. Food Protect. 2018, 81, 743–753. [Google Scholar] [CrossRef]
- Corzo, N.; Lopezfandino, R.; Delgado, T.; Ramos, M.; Olano, A. Changes in furosine and proteins of uht-treated milks stored at high ambient-temperatures. Zeitschrift für Lebensmittel-Untersuchung und Forschung 1994, 198, 302–306. [Google Scholar] [CrossRef]
- Su, G.W.; Zheng, L.; Cui, C.; Yang, B.; Ren, J.; Zhao, M. Characterization of antioxidant activity and volatile compounds of Maillard reaction products derived from different peptide fractions of peanut hydrolysate. Food Res. Int. 2011, 44, 3250–3258. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.; Luo, W.; Wang, L.; Water Quality Center. Detection of disinfectant residual in liquid chlorine/chlorine dioxide combined disinfection process. China Water Wastewater 2018, 34, 58–61. [Google Scholar] [CrossRef]
- Ashkenazi, S.; Mayzahav, M.; Dinari, G.; Gabbay, U.; Zilberberg, R.; Samra, Z. Recent trends in the epidemiology of Shigella species in Israel. Clin. Infect. Dis. 1993, 17, 897–899. [Google Scholar] [CrossRef]
- Lee, G.M.; Jung-Kue, S. Flavor and taste characteristics of black pepper by combining nonthermal sterilization treatments. Food Eng. Prog. 2020, 24, 126–132. [Google Scholar] [CrossRef]
- Perni, S.; Kong, M.G.; Prokopovich, P. Cold atmospheric pressure gas plasma enhances the wear performance of ultra-high molecular weight polyethylene. Acta Biomater. 2012, 8, 1357–1365. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V. Essential oils: Sources of antimicrobials and food preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef]
- Raguso, R.A. More lessons from linalool: Insights gained from a ubiquitous floral volatile. Curr. Opin. Plant Biol. 2016, 32, 31–36. [Google Scholar] [CrossRef]
- Aprotosoaie, A.C.; Hancianu, M.; Costache, I.; Miron, A. Linalool: A review on a key odorant molecule with valuable biological properties. Flavour Fragr. J. 2014, 29, 193–219. [Google Scholar] [CrossRef]
- Cheng, B.-H.; Sheen, L.-Y.; Chang, S.-T. Hypolipidemic effects of S-(+)-linalool and essential oil from Cinnamomum osmophloeum ct. linalool leaves in mice. J. Tradit. Complementary Med. 2018, 8, 46–52. [Google Scholar] [CrossRef]
- Herman, A.; Tambor, K.; Herman, A. Linalool affects the antimicrobial efficacy of essential oils. Curr. Microbiol. 2016, 72, 165–172. [Google Scholar] [CrossRef]
- Zheng, X.Y.; Guo, J.L.; Rao, H.S.; Guo, D.; Huang, Y.X.; Shi, C. Antibacterial and antibiofilm activity of coenzyme Q0 against Vibrio parahaemolyticus. Food Control 2019, 109, 106955. [Google Scholar] [CrossRef]
- Akhtar, F.; Khan, A.U.; Misba, L.; Akhtar, K.; Ali, A. Antimicrobial and antibiofilm photodynamic therapy against vancomycin resistant Staphylococcus aureus (VRSA) induced infection in vitro and in vivo. Eur. J. Pharm. Biopharm. 2021, 160, 65–76. [Google Scholar] [CrossRef]
- Yu, C.; Liu, J.; Zhu, W.; Qin, N.; Ren, X.; Zhu, B.; Xia, X. Impact of dietary components on enteric infectious disease. Crit. Rev. Food Sci. 2021, 18, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Alminderej, F.; Bakari, S.; Almundarij, T.I.; Snoussi, M.; Aouadi, K.; Kadri, A. Antimicrobial and wound healing potential of a new chemotype from Piper cubeba L. essential oil and in silico study on S. aureus tyrosyl-tRNA synthetase protein. Plants 2021, 20, 205. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Wang, S.; Li, J.H.; Bai, F.T.; Yang, Y.P.; Xu, Y.F.; Shi, C. The antimicrobial activity of coenzyme Q(0) against planktonic and biofilm forms of Cronobacter sakazakii. Food Microbiol. 2019, 86, 103337. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cai, J.X.; Chen, H.X.; Zhong, Q.P.; Hou, Y.Q.; Chen, W.J. Antibacterial activity and mechanism of linalool against Pseudomonas aeruginosa. Microb. Pathogenesis 2020, 141, 103980. [Google Scholar] [CrossRef]
- Miceli, A.; Vetrano, F.; Sabatino, L.; D’Anna, F.; Moncada, A. Influence of preharvest gibberellic acid treatments on postharvest quality of minimally processed leaf lettuce and rocket. Horticulturae 2019, 5, 63. [Google Scholar] [CrossRef]
- Sadekuzzaman, M.; Yang, S.; Kim, H.S.; Mizan, M.F.R.; Ha, S.D. Evaluation of a novel antimicrobial (lauric arginate ester) substance against biofilm of Escherichia coli O157: H7, Listeria monocytogenes, and Salmonella spp. Int. J. Food Sci. Technol. 2017, 52, 2058–2067. [Google Scholar] [CrossRef]
- Bagamboula, C.F.; Uyttendaele, M.; Debevere, J. Inhibitory effect of thyme and basil essential oils, carvacrol, thymol, estragol, linalool and p-cymene towards Shigella sonnei and S. flexneri. Food Microbiol. 2004, 21, 33–42. [Google Scholar] [CrossRef]
- Kang, S.M.; Li, X.J.; Xing, Z.Y.; Liu, X.; Bai, X.Y.; Yang, Y.P.; Shi, C. Antibacterial effect of citral on yersinia enterocolitica and its mechanism. Food Control 2022, 135, 108775. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, K.; Zhang, K.; Zhang, J.; Fu, J.; Li, J. Antibacterial Activity of Cinnamomum camphora essential oil on Escherichia coli during planktonic growth and biofilm formation. Front. Microbiol. 2020, 11, 561002. [Google Scholar] [CrossRef]
- Kang, J.H.; Song, K.B. Combined washing effect of noni extract and oregano essential oil on the decontamination of Listeria monocytogenes on romaine lettuce. Int. J. Food Sci. Technol. 2020, 55, 3515–3523. [Google Scholar] [CrossRef]
- Krasniewska, K.; Kosakowska, O.; Pobiega, K.; Gniewosz, M. The Influence of Two-Component mixtures from Spanish origanum oil with Spanish marjoram oil or Coriander oil on antilisterial activity and sensory quality of a fresh cut vegetable mixture. Foods 2021, 9, 1740. [Google Scholar] [CrossRef]
- Viacava, G.E.; Ayala-Zavala, J.F.; Gonzalez-Aguilar, G.A.; Ansorena, M.R. Effect of free and microencapsulated thyme essential oil on quality attributes of minimally processed lettuce. Postharvest Biol. Technol. 2018, 145, 125–133. [Google Scholar] [CrossRef]
- Rhen, M. Salmonella and reactive oxygen species: A love-hate relationship. J. Innate Immun. 2019, 11, 216–226. [Google Scholar] [CrossRef]
- Ma, M.M.; Zhao, J.X.; Yan, X.H.; Zeng, Z.L.; Wan, D.M.; Yu, P.; Xia, J.H.; Zhang, G.H.; Gong, D.M. Synergistic effects of monocaprin and carvacrol against Escherichia coli O157:H7 and Salmonella Typhimurium in chicken meat preservation. Food Control 2021, 132, 108480. [Google Scholar] [CrossRef]
- Peralta, D.R.; Adler, C.; Corbalan, N.S.; Garcia, E.C.P.; Pomares, M.F.; Vincent, P.A. Enterobactin as part of the oxidative stress response repertoire. PLoS ONE 2016, 11, e0157799. [Google Scholar] [CrossRef]
- Dalecki, A.G.; Crawford, C.L.; Wolschendorf, F. Copper and antibiotics: Discovery, modes of action, and opportunities for medicinal applications. Microbiol. Met. Ions 2017, 10, 193–260. [Google Scholar]
- Han, Y.J.; Sun, Z.C.; Chen, W.W. Antimicrobial susceptibility and antibacterial mechanism of limonene against Listeria monocytogenes. Molecules 2020, 25, 33. [Google Scholar] [CrossRef]
- Cui, H.Y.; Zhang, X.J.; Zhou, H.; Zhao, C.T.; Lin, L. Antimicrobial activity and mechanisms of Salvia sclarea essential oil. Bot. Stud. 2015, 56, 16. [Google Scholar] [CrossRef]
- Stratakos, A.C.; Sima, F.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Stef, L.; Pet, I.; Corcionivoschi, N. The in vitro effect of carvacrol, a food additive, on the pathogenicity of O157 and non-O157 Shiga-toxin producing Escherichia coli. Food Control 2018, 84, 290–296. [Google Scholar] [CrossRef]
- Ju, J.; Xie, Y.F.; Yu, H.; Guo, Y.H.; Cheng, Y.L.; Zhang, R.R.; Yao, W.R. Synergistic inhibition effect of citral and eugenol against Aspergillus niger and their application in bread preservation. Food Chem. 2020, 310, 125974. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Lee, D.G. Resveratrol induces membrane and DNA disruption via pro-oxidant activity against Salmonella typhimurium. Biochem. Biophys. Res. Commun. 2017, 489, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Gao, Y.; Fu, X.; Chen, W.; Yang, J.H.; Chen, Z.Y.; Wang, Z.X.; Zhuansun, X.X.; Feng, J.G.; Chen, Y. Preparation of peppermint oil nanoemulsions: Investigation of stability, antibacterial mechanism and apoptosis effects. Colloid Surf. B 2021, 201, 111626. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.K.; Das, G.; Baek, K.H. Antibacterial mechanism of the action of Enteromorpha linza L. essential oil against Escherichia coli and Salmonella Typhimurium. Bot. Stud. 2015, 56, 13. [Google Scholar] [CrossRef]
- Lin, Y.L.; Tang, X.; Xu, L.Z.; Wang, S.Y. Antibacterial properties and possible action mechanism of chelating peptides-zinc nanocomposite against Escherichia coli. Food Control 2019, 106, 106675. [Google Scholar] [CrossRef]
- Luo, K.Y.; Zhao, P.Y.; He, Y.F.; Kang, S.N.; Shen, C.Y.; Shi, C. Antibacterial effect of oregano essential oil against Vibrio vulnificus and its mechanism. Foods 2022, 11, 403. [Google Scholar] [CrossRef]
- Jha, R.; Regmi, R.; Simon, L.S. Efficacy of Eucalyptus essential oil against leaf spot (Alternaria solani) of Solanum melongena L. J. Agric. Biol. Sci. 2014, 9, 320–322. [Google Scholar]
- Gunduz, G.T.; Gonul, S.A.; Karapinar, M. Efficacy of oregano oil in the inactivation of Salmonella typhimurium on lettuce. Food Control 2010, 21, 513–517. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Color Parameter | Treatment | ||||
|---|---|---|---|---|---|
| Control | 1/2× MIC | 1× MIC | 3/2× MIC | 2× MIC | |
| L* | 59.68 ± 1.80 a | 56.86 ± 1.29 a | 60.08 ± 0.77 a | 57.34 ± 3.12 a | 56.99 ± 3.04 a |
| a* | −6.60 ± 0.67 a | −7.65 ± 0.32 a | −6.99 ± 0.19 a | −6.81 ± 0.96 a | −6.88 ± 1.09 a |
| b* | 19.76 ± 2.05 a | 22.48 ± 1.34 a | 22.18 ± 0.50 a | 20.37 ± 2.18 a | 21.33 ± 3.13 a |
| ΔE | 4.81 ± 0.53 a | 4.46 ± 0.52 a | 5.21 ± 0.38 a | 4.00 ± 0.76 a | 4.89 ± 0.23 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, R.; Guo, P.; Zhang, Z.; Wang, J.; Guo, X.; Guo, D.; Wang, Y.; Lü, X.; Shi, C. Antibacterial Activity and Mechanism of Linalool against Shigella sonnei and Its Application in Lettuce. Foods 2022, 11, 3160. https://doi.org/10.3390/foods11203160
Su R, Guo P, Zhang Z, Wang J, Guo X, Guo D, Wang Y, Lü X, Shi C. Antibacterial Activity and Mechanism of Linalool against Shigella sonnei and Its Application in Lettuce. Foods. 2022; 11(20):3160. https://doi.org/10.3390/foods11203160
Chicago/Turabian StyleSu, Ruiying, Peng Guo, Ziruo Zhang, Jingzi Wang, Xinyi Guo, Du Guo, Yutang Wang, Xin Lü, and Chao Shi. 2022. "Antibacterial Activity and Mechanism of Linalool against Shigella sonnei and Its Application in Lettuce" Foods 11, no. 20: 3160. https://doi.org/10.3390/foods11203160
APA StyleSu, R., Guo, P., Zhang, Z., Wang, J., Guo, X., Guo, D., Wang, Y., Lü, X., & Shi, C. (2022). Antibacterial Activity and Mechanism of Linalool against Shigella sonnei and Its Application in Lettuce. Foods, 11(20), 3160. https://doi.org/10.3390/foods11203160