
Bioaccessibility and Microencapsulation of Lactobacillus sp. to Enhance Nham Protein Hydrolysates in Thai Fermented Sausage

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microorganism and Culture Condition
2.2. Preparation of Culture Suspensions
2.3. Encapsulation of Probiotics Using the Freeze-Drying Technique
2.4. Estimation of Probiotic-Encapsulation with Probiotic Property
2.4.1. Growth of LAB in Different Lysozyme, pH, and Bile Conditions
2.4.2. Antimicrobial Activity
2.5. Cell Enumeration and Encapsulation Efficiency
2.6. Morphological Characterization
2.7. Nham Fermentation Using an Encapsulated-Probiotic Starter
2.8. Physicochemical Analysis and LAB Count of Nham Products
2.9. Preparation of Nham Protein Hydrolysates
2.10. Antioxidant Activity of NPHs
2.11. Antimicrobial Activity of NPHs
2.12. Statistical Analysis
3. Results and Discussion
3.1. Probiotic Characteristics of the Starter
3.2. Effects of GRF-Inulin Microparticles on Encapsulation Efficiency and Morphological Characteristics
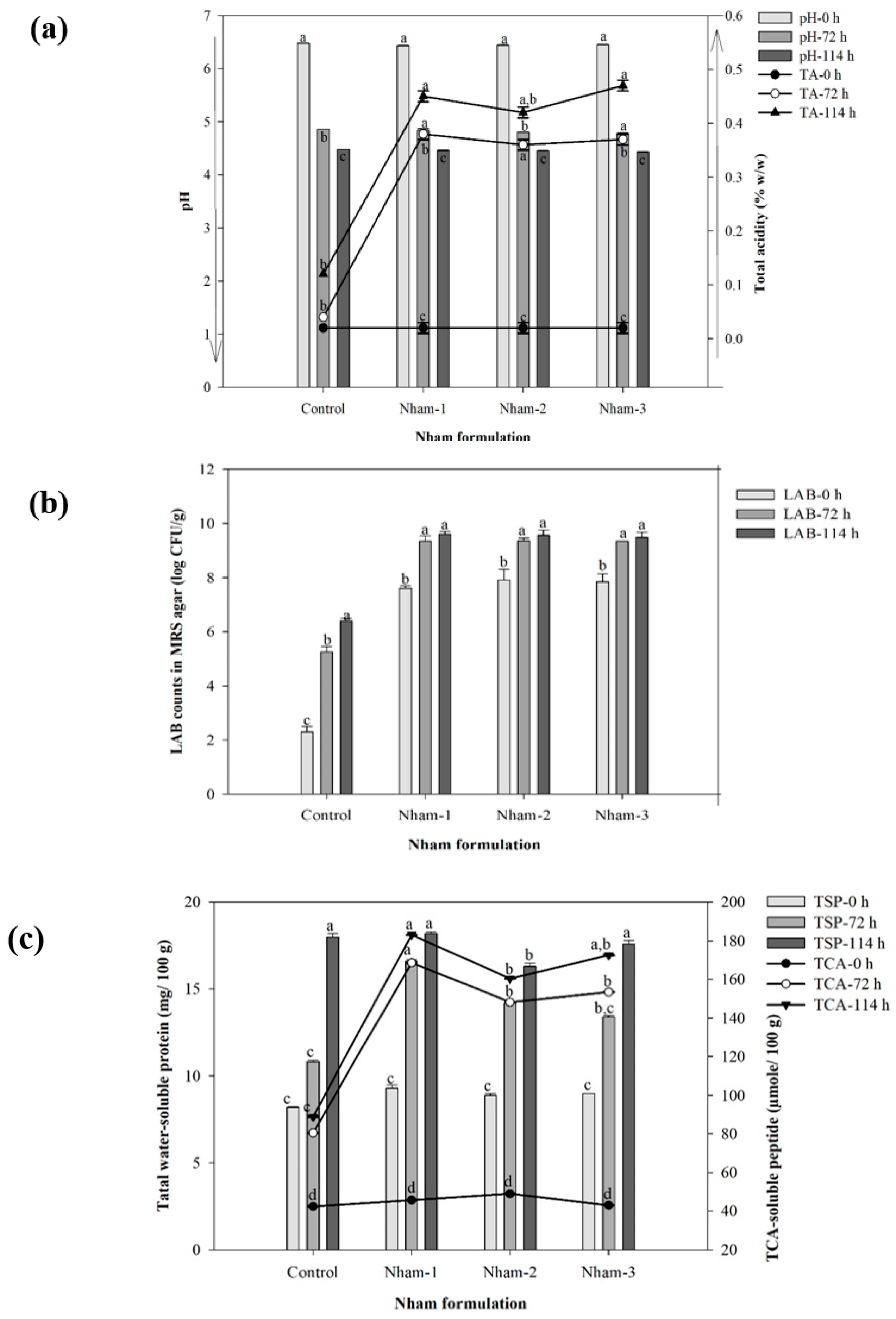
3.3. Physicochemical and Microbiological Characteristics of Nham Probiotic through Encapsulated Lactobacillus Starter Fermentation
3.4. Antioxidant Activity of Lyophilized NPHs
3.4.1. DPPH Radical-Scavenging Activity
3.4.2. FRAP-Reducing Power Capacity
3.5. Antimicrobial Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obradović, N.; Volić, M.; Nedović, V.; Rakin, M.; Bugarski, B. Microencapsulation of Probiotic Starter Culture in Protein–Carbohydrate Carriers Using Spray and Freeze-Drying Processes: Implementation in Whey-Based Beverages. J. Food Eng. 2022, 321, 110948. [Google Scholar] [CrossRef]
- Ann, E.Y.; Kim, Y.; Oh, S.; Imm, J.Y.; Park, D.J.; Han, K.S.; Kim, S.H. Microencapsulation of Lactobacillus acidophilus ATCC 43121 with Prebiotic Substrates Using a Hybridisation System. Int. J. Food Sci. Technol. 2007, 42, 411–419. [Google Scholar] [CrossRef]
- Huq, T.; Khan, A.; Khan, R.A.; Riedl, B.; Lacroix, M. Encapsulation of Probiotic Bacteria in Biopolymeric System. Crit. Rev. Food Sci. Nutr. 2013, 53, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Santivarangkna, C.; Kulozik, U.; Foerst, P. Inactivation Mechanisms of Lactic Acid Starter Cultures Preserved by Drying Processes. J. Appl. Microbiol. 2008, 105, 1–13. [Google Scholar] [CrossRef]
- Jagannath, A.; Raju, P.S.; Bawa, A.S. Comparative Evaluation of Bacterial Cellulose (Nata) as a Cryoprotectant and Carrier Support during the Freeze-Drying Process of Probiotic Lactic Acid Bacteria. LWT Food Sci. Technol. 2010, 43, 1197–1203. [Google Scholar] [CrossRef]
- Martín, M.J.; Lara-Villoslada, F.; Ruiz, M.A.; Morales, M.E. Microencapsulation of Bacteria: A Review of Different Technologies and Their Impact on the Probiotic Effects. Innov. Food Sci. Emerg. Technol. 2015, 27, 15–25. [Google Scholar] [CrossRef]
- Niamah, A.K.; Al-Sahlany, S.T.G.; Ibrahim, S.A.; Verma, D.K.; Thakur, M.; Singh, S.; Patel, A.R.; Aguilar, C.N.; Utama, G.L. Electro-Hydrodynamic Processing for Encapsulation of Probiotics: A Review on Recent Trends, Technological Development, Challenges and Future Prospect. Food Biosci. 2021, 44, 101458. [Google Scholar] [CrossRef]
- Santos Monteiro, S.; Albertina, Y.; Beserra, S.; Miguel, H.; Oliveira, L.; De Bittencourt Pasquali, M.A. Production of Probiotic Passion Fruit (Passiflora edulis Sims f. Flavicarpa Deg.) Drink Using Lactobacillus reuteri and Microencapsulation via Spray Drying. Foods 2020, 9, 335. [Google Scholar] [PubMed] [Green Version]
- Samedi, L.; Linton Charles, A. Viability of 4 Probiotic Bacteria Microencapsulated with Arrowroot Starch in the Simulated Gastrointestinal Tract (GIT) and Yoghurt. Foods 2019, 8, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aswathy, R.G.; Ismail, B.; John, R.P.; Nampoothiri, K.M. Evaluation of the Probiotic Characteristics of Newly Isolated Lactic Acid Bacteria. Appl. Biochem. Biotechnol. 2008, 151, 244–255. [Google Scholar] [CrossRef]
- Fernández De Palencia, P.; López, P.; Corbí, A.L.; Peláez, C.; Requena, T. Probiotic Strains: Survival under Simulated Gastrointestinal Conditions, In Vitro Adhesion to Caco-2 Cells and Effect on Cytokine Secretion. Eur. Food Res. Technol. 2008, 227, 1475–1484. [Google Scholar] [CrossRef] [Green Version]
- Nedelcheva, P.; Denkova, Z.; Denev, P.; Slavchev, A.; Krastanov, A. Probiotic Strain Lactobacillus plantarum NBIMCC 2415 with Antioxidant Activity as a Starter Culture in the Production of Dried Fermented Meat Products. Biotechnol. Biotechnol. Equip. 2010, 24, 1624–1630. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.R.; Sardi, J.d.C.O.; Pitangui, N.d.S.; Roque, S.M.; da Silva, A.C.B.; Rosalen, P.L. Probiotics as an Alternative Antimicrobial Therapy: Current Reality and Future Directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- Visessanguan, W.; Benjakul, S.; Smitinont, T.; Kittikun, C.; Thepkasikul, P.; Panya, A. Changes in Microbiological, Biochemical and Physico-Chemical Properties of Nham Inoculated with Different Inoculum Levels of Lactobacillus curvatus. LWT Food Sci. Technol. 2006, 39, 814–826. [Google Scholar] [CrossRef]
- Paludan-Müller, C.; Madsen, M.; Sophanodora, P.; Gram, L.; Møller, P.L. Fermentation and Microflora of Plaa-Som, a Thai Fermented Fish Product Prepared with Different Salt Concentrations. Int. J. Food Microbiol. 2002, 73, 61–70. [Google Scholar] [CrossRef]
- Mohammadi, R.; Sohrabvandi, S.; Mohammad Mortazavian, A. The Starter Culture Characteristics of Probiotic Microorganisms in Fermented Milks. Eng. Life Sci. 2012, 12, 399–409. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Pateiro, M.; Tomasevic, I.; Domínguez, R.; da Silva Barretto, A.C.; Santos, E.M.; Lorenzo, J.M. Functional Fermented Meat Products with Probiotics—A Review. J. Appl. Microbiol. 2021, 133, 91–103. [Google Scholar] [CrossRef]
- Jemil, I.; Jridi, M.; Nasri, R.; Ktari, N.; ben Slama-Ben Salem, R.; Mehiri, M.; Hajji, M.; Nasri, M. Functional, Antioxidant and Antibacterial Properties of Protein Hydrolysates Prepared from Fish Meat Fermented by Bacillus subtilis A26. Process Biochem. 2014, 49, 963–972. [Google Scholar] [CrossRef]
- Phupaboon, S.; Piyatheerawong, W.; Yunchalard, S. Correlation of Antioxidant Activity and Protein Alteration of Silver BARB Protein Hydrolysates. Asia Pac. J. Sci. Technol. 2020, 25, APST-25-04-05. [Google Scholar]
- Phupaboon, S.; Punyauppa-Path, S.; Kontongdee, P.; Piyatheerawong, W.; Yunchalard, S. Development and Enhancement of Antioxidant Peptides from Spontaneous Plaa-Som Fermentation Co-Stimulated with Chiangrai Phulae Pineapple Enzymatic Reaction. Int. Food Res. J. 2022, 29, 406–415. [Google Scholar] [CrossRef]
- Luan, X.; Feng, M.; Sun, J. Effect of Lactobacillus plantarum on Antioxidant Activity in Fermented Sausage. Int. Food Res. J. 2021, 144, 110351. [Google Scholar] [CrossRef]
- Su, Y. Isolation and Identification of Pelteobagrin, a Novel Antimicrobial Peptide from the Skin Mucus of Yellow Catfish (Pelteobagrus fulvidraco). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.C.; Feng, M.Q.; Sun, J.; Xu, X.L.; Zhou, G.H. Screening of Lactic Acid Bacteria with High Protease Activity from Fermented Sausages and Antioxidant Activity Assessment of Its Fermented Sausages. CYTA J. Food 2019, 17, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Huang, X.; Zhang, H.; Zhang, Y.; Gänzle, M.; Yang, N.; Nisninari, K.; Fang, Y. Probiotic encapsulation in water-in-water emulsion via heteroprotein complex coacervation of type-A gelatin/sodium caseinate. Food Hydrocoll. 2020, 105, 105790. [Google Scholar] [CrossRef]
- Afzaal, M.; Saeed, F.; Ateeq, H.; Imran, A.; Yasmin, I.; Shahid, A.; Javed, A.; Sahid, Y.A.; Islan, F.; Ofoedu, C.E.; et al. Survivability of probiotics under hostile conditions as affected by prebiotic-based encapsulating materials. Int. J. Food Prop. 2022, 25, 2044–2054. [Google Scholar] [CrossRef]
- Kimoto-Nira, H.; Kobayashi, M.; Nomura, M.; Sasaki, K.; Suzuki, C. Bile Resistance in Lactococcus lactis Strains Varies with Cellular Fatty Acid Composition: Analysis by Using Different Growth Media. Int. J. Food Microbiol. 2009, 131, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Blaszyk, M.; Holley, R.A. Interaction of Monolaurin, Eugenol and Sodium Citrate on Growth of Common Meat Spoilage and Pathogenic Organisms. Int. J. Food Microbiol. 1998, 39, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Antimicrobial Activity of Basil (Ocimum basilicum) Oil against Salmonella enteritidis In Vitro and in Food. Biosci. Biotechnol. Biochem. 2010, 74, 1200–1204. [Google Scholar] [CrossRef] [Green Version]
- AOAC International. AOAC Guidelines for Single Laboratory Validation of Chemical Methods for Dietary Supplements and Botanicals; AOAC: Rockville, MD, USA, 2002; pp. 1–38. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin-Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Ketnawa, S.; Wickramathilaka, M.; Liceaga, A.M. Changes on Antioxidant Activity of Microwave-Treated Protein Hydrolysates after Simulated Gastrointestinal Digestion: Purification and Identification. Food Chem. 2018, 254, 36–46. [Google Scholar] [CrossRef]
- Phupaboon, S.; Matra, M.; Prommachart, R.; Totakul, P.; Supapong, C.; Wanapat, M. Extraction, Characterization, and Chitosan Microencapsulation of Bioactive Compounds from Cannabis sativa L., Cannabis indica L., and Mitragyna speiosa K. Antioxidants 2022, 11, 2103. [Google Scholar] [CrossRef]
- Binsan, W.; Benjakul, S.; Visessanguan, W.; Roytrakul, S.; Tanaka, M.; Kishimura, H. Antioxidative Activity of Mungoong, an Extract Paste, from the Cephalothorax of White Shrimp (Litopenaeus vannamei). Food Chem. 2008, 106, 185–193. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to Select a Probiotic? A Review and Update of Methods and Criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Heller, K.J. Probiotic Bacteria in Fermented Foods: Product Characteristics and Starter Organisms. Am. J. Clin. Nutr. 2001, 73, 374S–379S. [Google Scholar] [CrossRef] [Green Version]
- Kimoto-Nira, H.; Suzuki, C.; Sasaki, K.; Kobayashi, M.; Mizumachi, K. Survival of a Lactococcus lactis Strain Varies with Its Carbohydrate Preference under In Vitro Conditions Simulated Gastrointestinal Tract. Int. J. Food Microbiol. 2010, 143, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Ogunremi, O.R.; Sanni, A.I.; Agrawal, R. Probiotic Potentials of Yeasts Isolated from Some Cereal-Based Nigerian Traditional Fermented Food Products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Isolation and Preliminary Characterization of a Bacteriocin Produced by Lactobacillus plantarum N014 Isolated from Nham, a Traditional Thai Fermented Pork. J. Food Prot. 2006, 69, 1937–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, R.V.D.B.; Borges, S.V.; Botrel, D.A. Gum Arabic/Starch/Maltodextrin/Inulin as Wall Materials on the Microencapsulation of Rosemary Essential Oil. Carbohydr. Polym. 2014, 101, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Chadong, K.; Yunchalard, S.; Piyatheerawong, W. Physicochemical Characteristics and Protein Degradation during Fermentation of Plaa-Som, A Traditional Fermented Fish Product of North-Eastern Thailand. Indian J. Tradit. Knowl. 2015, 14, 220–225. [Google Scholar]
- Paludan-Müller, C.; Henrik Huss, H.; Gram, L. Characterization of Lactic Acid Bacteria Isolated from a Thai Low-Salt Fermented Fish Product and the Role of Garlic as Substrate for Fermentation. Int. J. Food Microbiol. 1999, 46, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Visessanguan, W.; Benjakul, S.; Riebroy, S.; Thepkasikul, P. Changes in Composition and Functional Properties of Proteins and Their Contributions to Nham Characteristics. Meat Sci. 2004, 66, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Martínez-Alvarez, O.; Benjakul, S.; Rawdkuen, S. Gelatin Hydrolysates from Farmed Giant Catfish Skin Using Alkaline Proteases and Its Antioxidative Function of Simulated Gastro-Intestinal Digestion. Food Chem. 2016, 192, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ktari, N.; Jridi, M.; Bkhairia, I.; Sayari, N.; ben Salah, R.; Nasri, M. Functionalities and Antioxidant Properties of Protein Hydrolysates from Muscle of Zebra Blenny (Salaria basilisca) Obtained with Different Crude Protease Extracts. Int. Food Res. J. 2012, 49, 747–756. [Google Scholar] [CrossRef]
- Khantaphant, S.; Benjakul, S. Comparative Study on the Proteases from Fish Pyloric Caeca and the Use for Production of Gelatin Hydrolysate with Antioxidative Activity. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Feng, M.-q.; Sun, J.; Xu, X.-l.; Zhou, G.-h. Protein Degradation and Peptide Formation with Antioxidant Activity in Pork Protein Extracts Inoculated with Lactobacillus plantarum and Staphylococcus simulans. Meat Sci. 2020, 160, 107958. [Google Scholar] [CrossRef] [PubMed]
- Bougatef, A.; Nedjar-Arroume, N.; Manni, L.; Ravallec, R.; Barkia, A.; Guillochon, D.; Nasri, M. Purification, and Identification of Novel Antioxidant Peptides from Enzymatic Hydrolysates of Sardinelle (Sardinella aurita) By-Products Proteins. Food Chem. 2010, 118, 559–565. [Google Scholar] [CrossRef]
- Ranathunga, S.; Rajapakse, N.; Kim, S.K. Purification and Characterization of Antioxidative Peptide Derived from Muscle of Conger Eel (Conger myriaster). Eur. Food Res. Technol. 2006, 222, 310–315. [Google Scholar] [CrossRef]
- Jun, S.Y.; Park, P.J.; Jung, W.K.; Kim, S.K. Purification and Characterization of an Antioxidative Peptide from Enzymatic Hydrolysate of Yellowfin Sole (Limanda aspera) Frame Protein. Eur. Food Res. Technol. 2004, 219, 20–26. [Google Scholar]
- Cheung, I.W.Y.; Cheung, L.K.Y.; Tan, N.Y.; Li-Chan, E.C.Y. The Role of Molecular Size in Antioxidant Activity of Peptide Fractions from Pacific Hake (Merluccius productus) Hydrolysates. Food Chem. 2012, 134, 1297–1306. [Google Scholar] [CrossRef]
- Doan, C.H.; Davidson, P.M. Growth and Production of Enterotoxin A by Staphylococcus aureus on “Home-Style” French Fries. J. Food Sci. 1999, 64, 913–917. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Mixing Ratio of Probiotic Starter Encapsulated Powder | Amount Ratio [% (w/v)] |
|---|---|---|
| Control | Spontaneous fermentation | no fill |
| Nham-1 | L. casei KKU-KK1: L. pentosus KKU-KK2: L. acidophilus KKU-KK3 | 1:1:2 |
| Nham-2 | L. casei KKU-KK1: L. pentosus KKU-KK2: L. acidophilus KKU-KK3 | 2:1:2 |
| Nham-3 | L. casei KKU-KK1: L. pentosus KKU-KK2: L. acidophilus KKU-KK3 | 1:2:2 |
| Probiotic Strains | (%) Survival Rate of Resistive Difference Conditions | Antimicrobial Activity (mm) | ||||
|---|---|---|---|---|---|---|
| Lysozyme | pH (3.0) | Bile salt | E. coli | S. aureus | Ent. aerogenes | |
| The probiotic potential before encapsulation | ||||||
| KKU-KK1 | 76.6 | 85.3 | 75.2 | 17.0 | 19.8 | 19.4 |
| KKU-KK2 | 70.2 | 65.0 | 60.2 | 9.7 | 12.3 | 18.0 |
| KKU-KK3 | 75.8 | 65.2 | 68.0 | 16.2 | 11.5 | 7.7 |
| The probiotic potential after encapsulation | ||||||
| KKU-KK1 | 70.4 | 62.8 | 65.6 | 18.9 | 20.8 | 20.6 |
| KKU-KK2 | 58.5 | 50.3 | 50.1 | 12.4 | 16.4 | 18.5 |
| KKU-KK3 | 62.4 | 61.8 | 60.4 | 16.6 | 12.2 | 11.3 |
| Probiotic Strains | Enumeration of Cell Viability (log CFU/g) | Encapsulation Efficiency (%) | Size (µm) | Zeta (mV) | |
|---|---|---|---|---|---|
| Initial Cell Suspension | After Encapsulated | ||||
| KKU-KK1 | 12.5 ± 0.0 a | 12.5 ± 0.0 a | 100.0 ± 0.0 a | 10.8 ± 0.1 a | −44.4 ± 0.3 a |
| KKU-KK2 | 12.7 ± 0.0 a | 12.7 ± 0.0 a | 100.0 ± 0.0 a | 10.3 ± 0.3 a | −43.8 ± 0.2 a |
| KKU-KK3 | 12.8 ± 0.0 a | 12.8 ± 0.1 a | 100.0 ± 0.0 a | 10.5 ± 0.2 a | −44.6 ± 0.0 a |
| NPH | Inhibition Diameter Zone of Antimicrobial Activity (mm) | |||
|---|---|---|---|---|
| E. coli | S. aureus | Ent. aerogenes | Sal. typhimurium | |
| NPH-control | 8.0 ± 1.0 c | 8.0 ± 0.0 c | 7.0 ± 2.0 c | - |
| NPH-nham1 | 22.0 ± 0.0 a | 16.0 ± 1.0 a | 18.0 ± 2.0 a | 17.0 ± 1.0 a |
| NPH-nham2 | 16.0 ± 1.0 b | 11.0 ± 3.0 b | 14.0 ± 1.0 b | 15.0 ± 2.0 c |
| NPH-nham3 | 20.0 ± 2.0 a,b | 13.0 ± 1.0 b | 15.0 ± 3.0 b | 16.0 ± 0.0 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phupaboon, S.; Kontongdee, P.; Hashim, F.J.; Kanpipit, N.; Matra, M.; Totakul, P.; Prommachart, R.; Phesatcha, B.; Wanapat, M. Bioaccessibility and Microencapsulation of Lactobacillus sp. to Enhance Nham Protein Hydrolysates in Thai Fermented Sausage. Foods 2022, 11, 3846. https://doi.org/10.3390/foods11233846
Phupaboon S, Kontongdee P, Hashim FJ, Kanpipit N, Matra M, Totakul P, Prommachart R, Phesatcha B, Wanapat M. Bioaccessibility and Microencapsulation of Lactobacillus sp. to Enhance Nham Protein Hydrolysates in Thai Fermented Sausage. Foods. 2022; 11(23):3846. https://doi.org/10.3390/foods11233846
Chicago/Turabian StylePhupaboon, Srisan, Papatchaya Kontongdee, Farah J. Hashim, Nattawadee Kanpipit, Maharach Matra, Pajaree Totakul, Ronnachai Prommachart, Burarat Phesatcha, and Metha Wanapat. 2022. "Bioaccessibility and Microencapsulation of Lactobacillus sp. to Enhance Nham Protein Hydrolysates in Thai Fermented Sausage" Foods 11, no. 23: 3846. https://doi.org/10.3390/foods11233846
APA StylePhupaboon, S., Kontongdee, P., Hashim, F. J., Kanpipit, N., Matra, M., Totakul, P., Prommachart, R., Phesatcha, B., & Wanapat, M. (2022). Bioaccessibility and Microencapsulation of Lactobacillus sp. to Enhance Nham Protein Hydrolysates in Thai Fermented Sausage. Foods, 11(23), 3846. https://doi.org/10.3390/foods11233846