
Effect of Mulberry Leaf TMR on Growth Performance, Meat Quality and Expression of Meat Quality Master Genes (ADSL, H-FABP) in Crossbred Black Goats

 ,
, 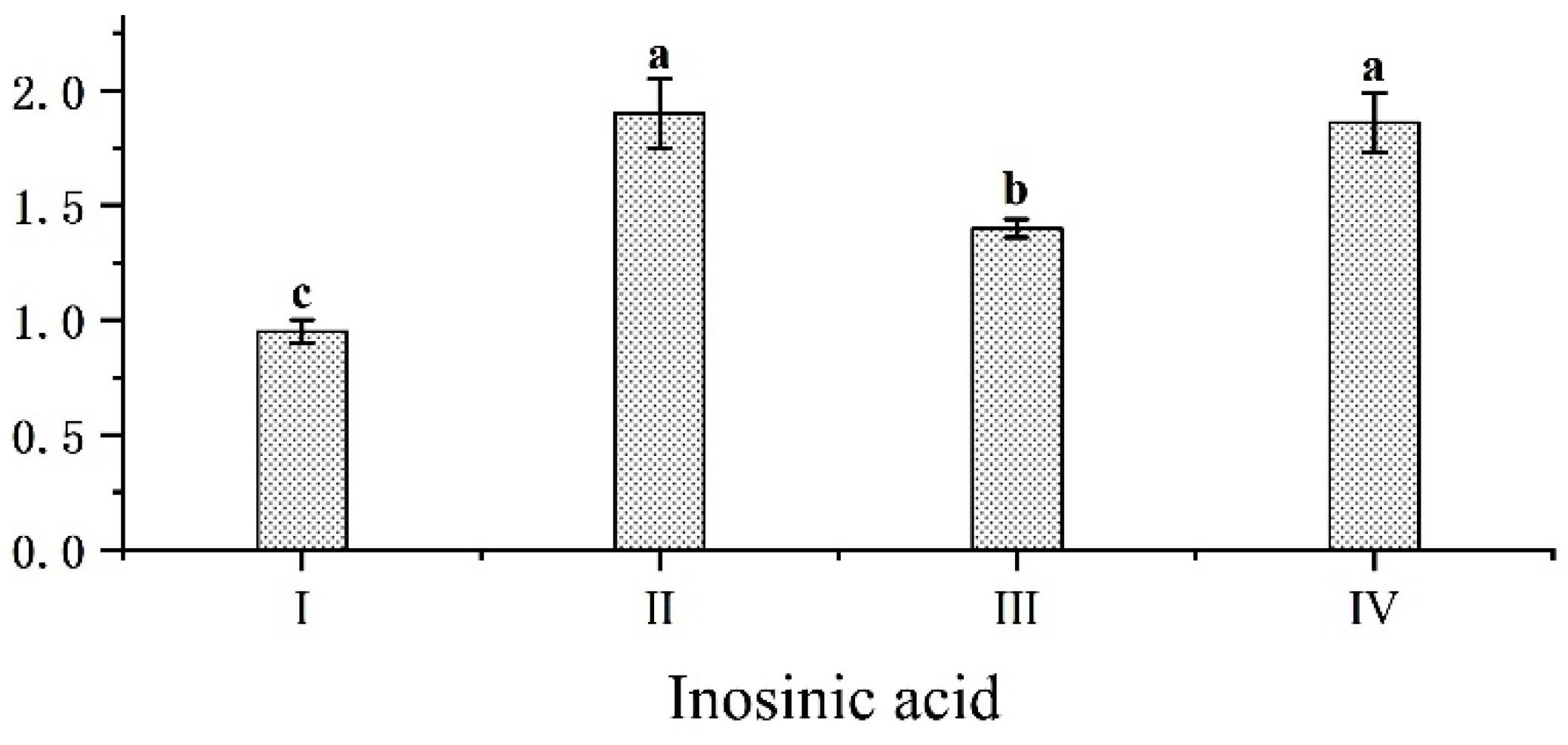{kind=link}
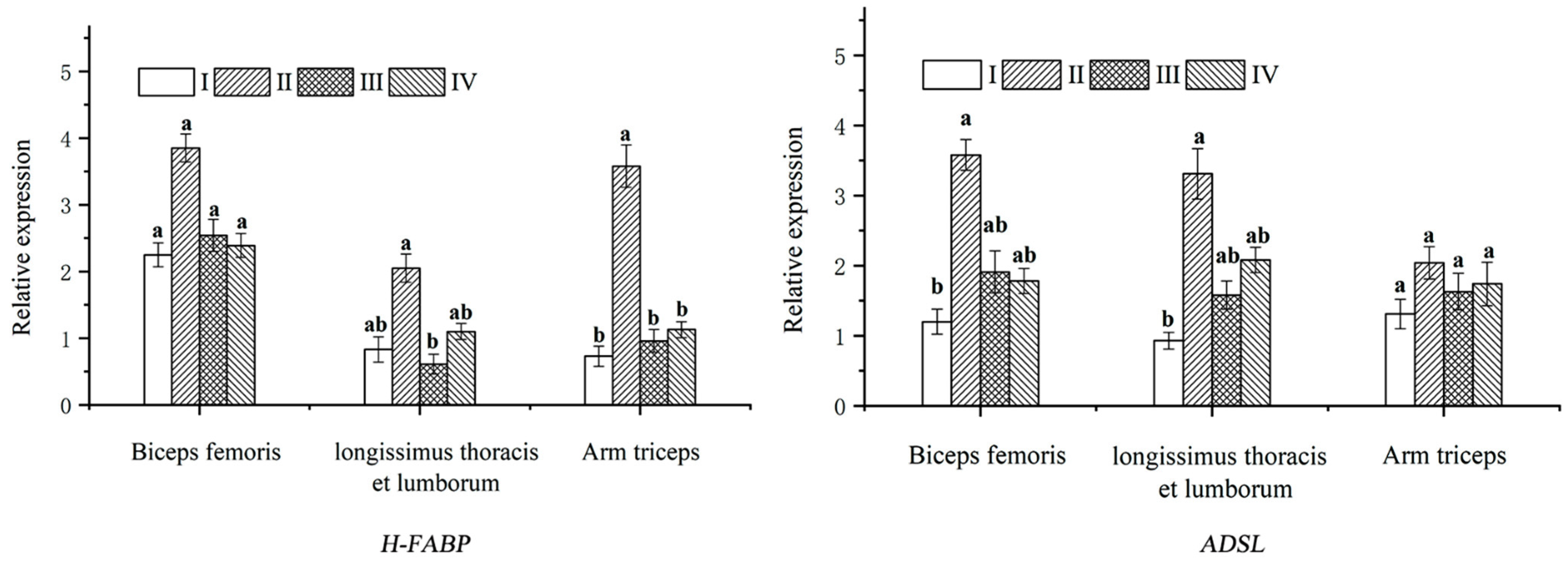{kind=link}
Abstract
Share and Cite
Long, Y.; Han, Y.; Zhao, Y.; Chen, D.; Wang, D.; Yang, Y.; Su, C.; Shen, X. Effect of Mulberry Leaf TMR on Growth Performance, Meat Quality and Expression of Meat Quality Master Genes (ADSL, H-FABP) in Crossbred Black Goats. Foods 2022, 11, 4032. https://doi.org/10.3390/foods11244032
Long Y, Han Y, Zhao Y, Chen D, Wang D, Yang Y, Su C, Shen X. Effect of Mulberry Leaf TMR on Growth Performance, Meat Quality and Expression of Meat Quality Master Genes (ADSL, H-FABP) in Crossbred Black Goats. Foods. 2022; 11(24):4032. https://doi.org/10.3390/foods11244032
Chicago/Turabian StyleLong, Yong, Yong Han, Yuanfeng Zhao, Dianqian Chen, Defeng Wang, Yang Yang, Chaozhi Su, and Xiaoyun Shen. 2022. "Effect of Mulberry Leaf TMR on Growth Performance, Meat Quality and Expression of Meat Quality Master Genes (ADSL, H-FABP) in Crossbred Black Goats" Foods 11, no. 24: 4032. https://doi.org/10.3390/foods11244032
APA StyleLong, Y., Han, Y., Zhao, Y., Chen, D., Wang, D., Yang, Y., Su, C., & Shen, X. (2022). Effect of Mulberry Leaf TMR on Growth Performance, Meat Quality and Expression of Meat Quality Master Genes (ADSL, H-FABP) in Crossbred Black Goats. Foods, 11(24), 4032. https://doi.org/10.3390/foods11244032