
The Effect of Dietary Supplementation with Resveratrol on Growth Performance, Carcass and Meat Quality, Blood Lipid Levels and Ruminal Microbiota in Fattening Goats

Abstract
:1. Introduction
2. Material and Methods
2.1. Animals and Experimental Diets
2.2. Slaughter Surveys and Sampling
2.3. Meat Quality and Blood Chemical Analysis
2.4. Microbiota Analysis by 16S Sequencing
2.5. Statistical Analysis
3. Results
3.1. Growth and Carcass Characteristics
3.2. Meat Quality Characteristics
3.3. Blood Physiological and Biochemical Indexes
3.4. Effects of Dietary RES Supplementation on Rumen Bacterial Microbiota
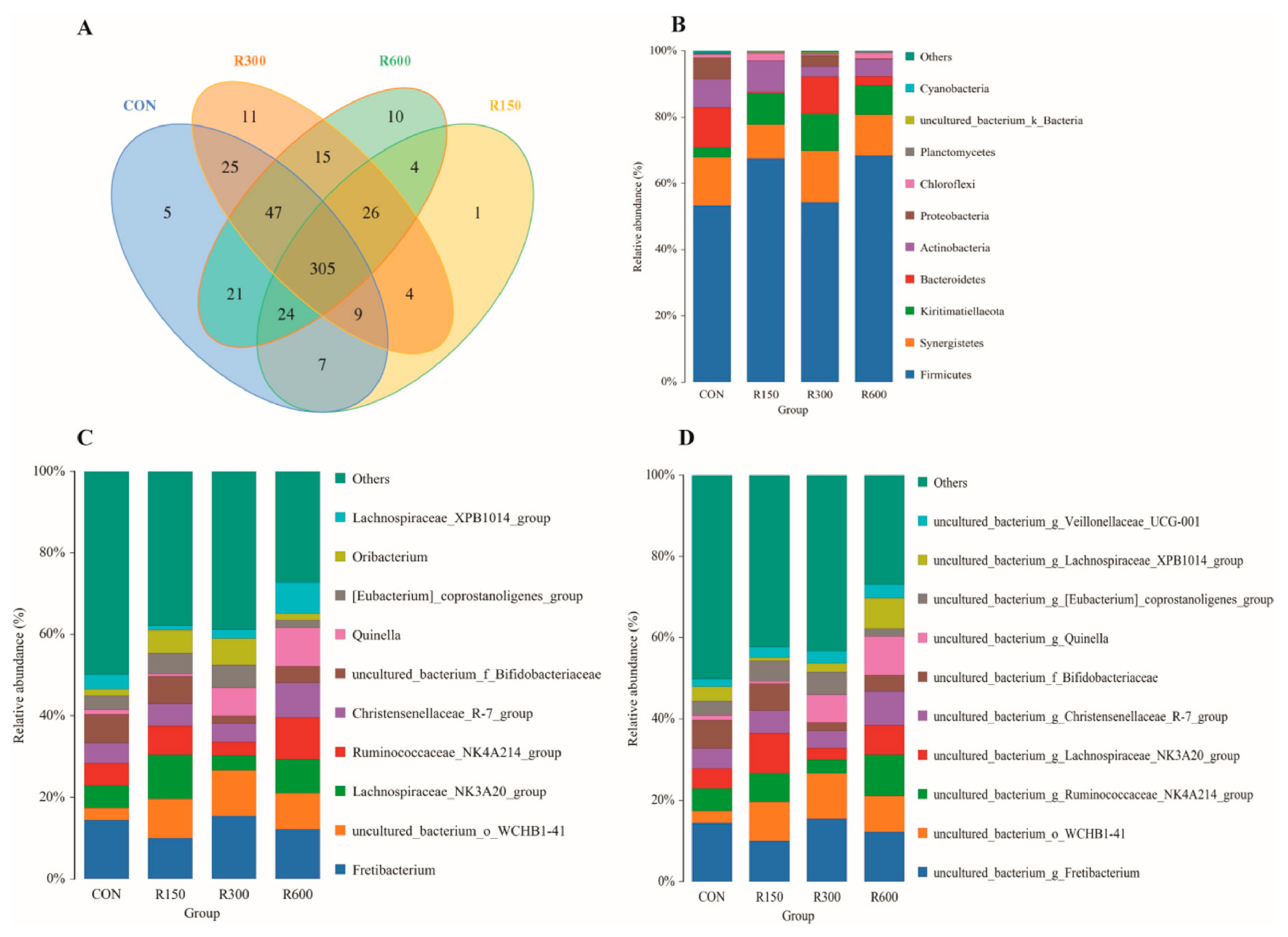
3.4.1. Taxonomic Identification of Rumen Bacteria across Treatments
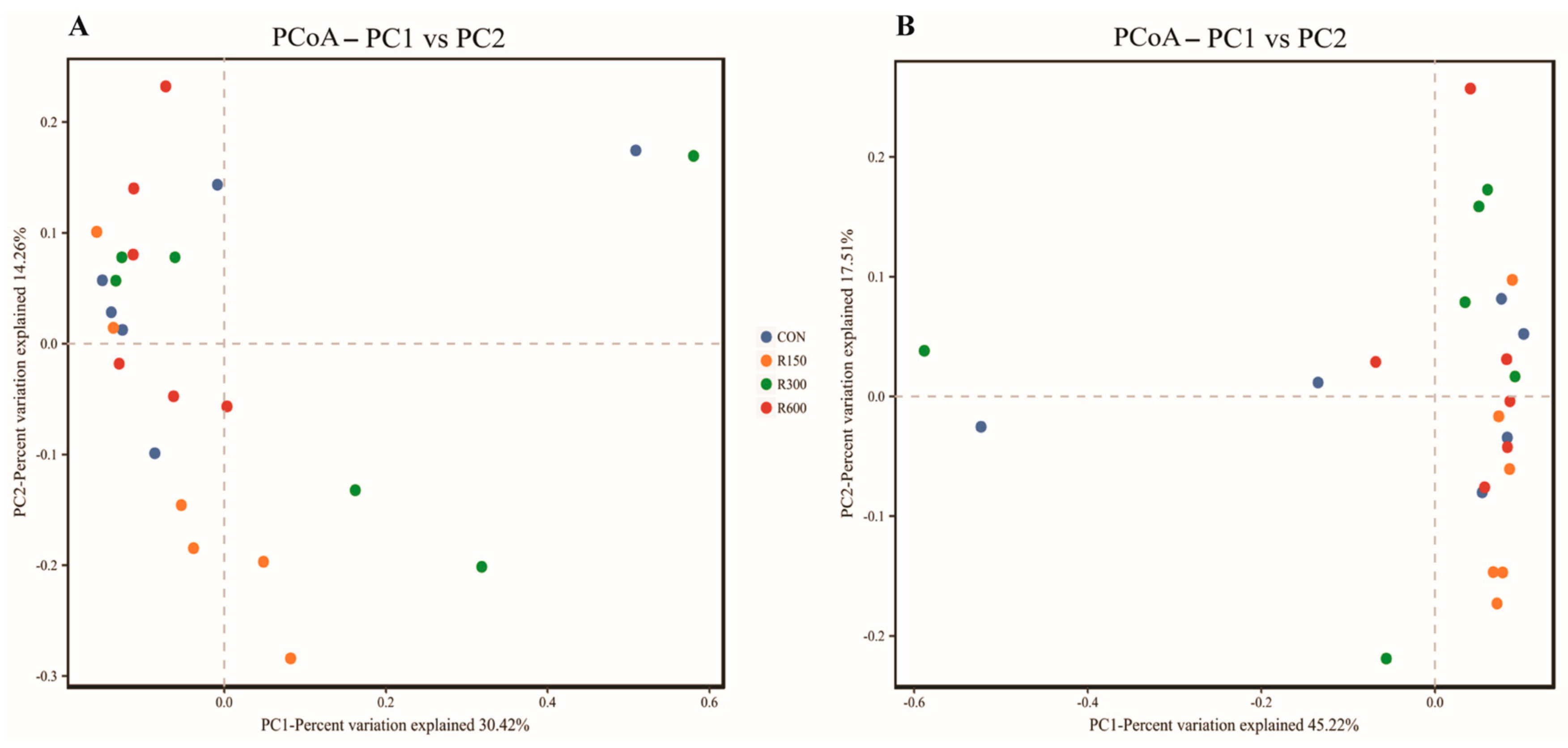
3.4.2. Effects of Dietary RES Supplementation on Microbial Diversity of Rumen Bacteria
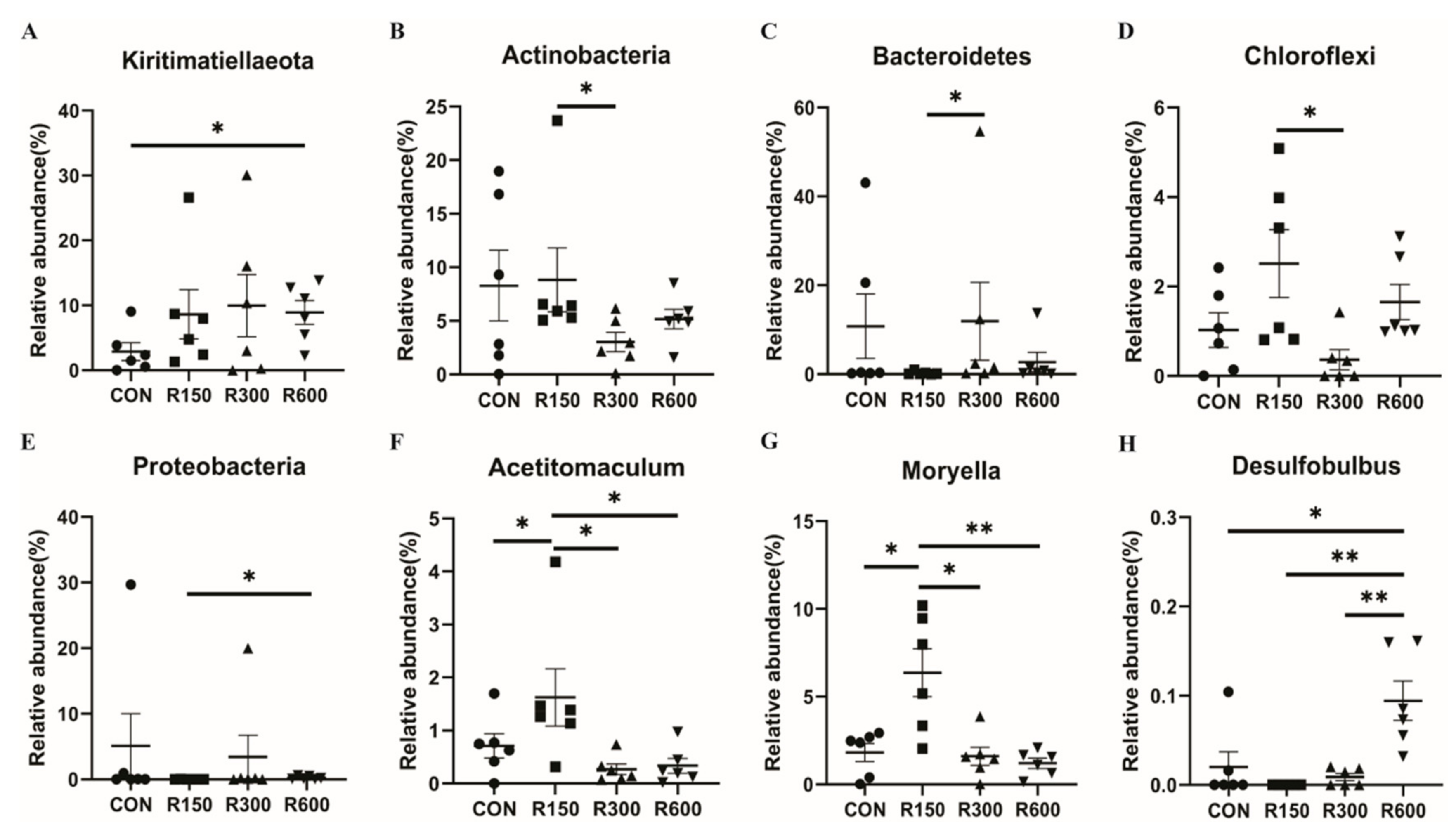
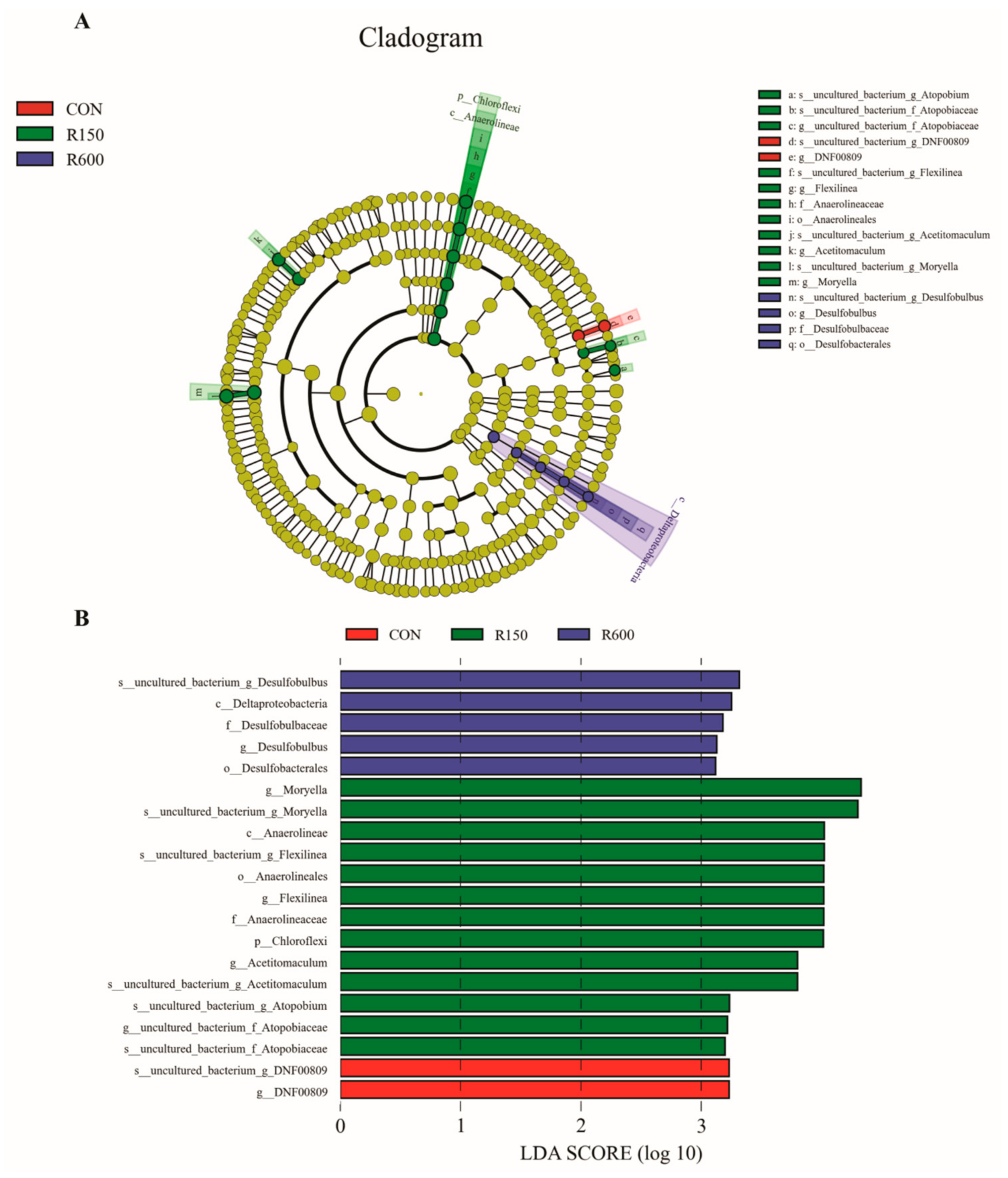
3.4.3. Differential Rumen Bacterial Taxa among Different Treatments
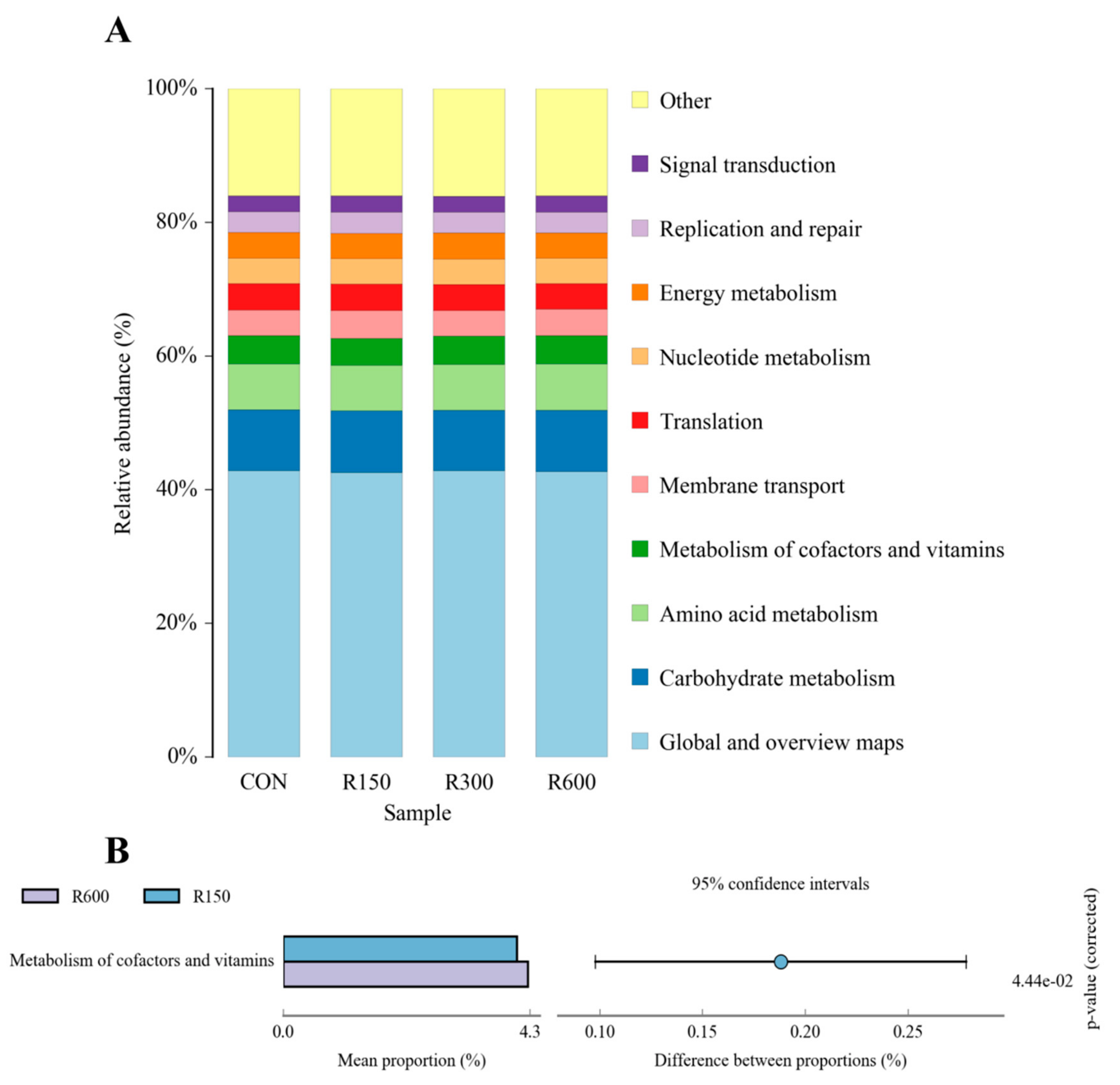
3.4.4. Functional Prediction of Rumen Microbiota among Different Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2589. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.L.; He, J.H.; Xie, H.B.; Yang, Y.S.; Li, J.C.; Zou, Y. Resveratrol induces antioxidant and heat shock protein mRNA expression in response to heat stress in black-boned chickens. Poult. Sci. 2014, 93, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, L.E.; Newton, R.; Kennedy, G.E.; Fenwick, P.S.; Leung, R.H.F.; Ito, K.; Russell, R.E.K.; Barnes, P.J. Anti-inflammatory effects of resveratrol in lung epithelial cells: Molecular mechanisms. Am. J. Physiol. Cell. Mol. Physiol. 2004, 287, L774–L783. [Google Scholar] [CrossRef] [Green Version]
- Xing, T.; Xu, X.L.; Zhou, G.H.; Wang, P.; Jiang, N.N. The effect of transportation of broilers during summer on the expression of heat shock protein 70, postmortem metabolism and meat quality. J. Anim. Sci. 2015, 93, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Xia, Q.; Cui, Y.; Qu, Q.; Wei, Y.; Jiang, Q. Resveratrol increase the proportion of oxidative muscle fiber through the AdipoR1-AMPK-PGC-1α pathway in pigs. J. Funct. Foods 2020, 73, 104090. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, J.; Yu, B.; Zheng, P.; Huang, Z.; Mao, X.; He, J.; Yu, J.; Chen, J.; Chen, D. Dietary resveratrol supplementation improves meat quality of finishing pigs through changing muscle fiber characteristics and antioxidative status. Meat Sci. 2015, 102, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, L.; Zhao, X.H.; Chen, X.Y.; Yang, L.; Geng, Z.Y. Dietary resveratrol supplementation prevents transport-stress-impaired meat quality of broilers through maintaining muscle energy metabolism and antioxidant status. Poult. Sci. 2017, 96, 2219–2225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhao, X.; Wang, L.; Yang, L.; Chen, X.; Geng, Z. Resveratrol beneficially affects meat quality of heat-stressed broilers which is associated with changes in muscle antioxidant status. Anim. Sci. J. 2017, 88, 1569–1574. [Google Scholar] [CrossRef]
- Meng, Q.; Sun, S.; Luo, Z.; Shi, B.; Shan, A.; Cheng, B. Maternal dietary resveratrol alleviates weaning-associated diarrhea and intestinal inflammation in pig offspring by changing intestinal gene expression and microbiota. Food Funct. 2019, 10, 5626–5643. [Google Scholar] [CrossRef]
- Jeon, Y.O.; Lee, J.-S.; Lee, H.G. Improving solubility, stability, and cellular uptake of resveratrol by nanoencapsulation with chitosan and γ-poly (glutamic acid). Colloids Surf. B Biointerfaces 2016, 147, 224–233. [Google Scholar] [CrossRef]
- Yin, X.; Liao, W.; Li, Q.; Zhang, H.; Liu, Z.; Zheng, X.; Zheng, L.; Feng, X. Interactions between resveratrol and gut microbiota affect the development of hepatic steatosis: A fecal microbiota transplantation study in high-fat diet mice. J. Funct. Foods 2020, 67, 103883. [Google Scholar] [CrossRef]
- Zhao, Y.N.; Cao, Y.N.; Sun, J.; Liang, Z.; Wu, Q.; Cui, S.H.; Zhi, D.F.; Guo, S.T.; Zhen, Y.H.; Zhang, S.B. Anti-breast cancer activity of resveratrol encapsulated in liposomes. J. Mater. Chem. B 2020, 8, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; Vallejo, F.; Pallarés, F.J.; Larrosa, M.; Lucas, R.; Morales, J.C.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Metabolites and tissue distribution of resveratrol in the pig. Mol. Nutr. Food Res. 2011, 55, 1154–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrosa, M.; Yañéz-Gascón, M.J.; Selma, M.V.; González-Sarrías, A.; Toti, S.; Ceron, J.; Tomas-Barberan, F.; Dolara, P.; Espín, J.C. Effect of a Low Dose of Dietary Resveratrol on Colon Microbiota, Inflammation and Tissue Damage in a DSS-Induced Colitis Rat Model. J. Agric. Food Chem. 2009, 57, 2211–2220. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Luo, T. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef]
- McGovern, E.; Kenny, D.A.; McCabe, M.S.; Fitzsimons, C.; McGee, M.; Kelly, A.K.; Waters, S.M. 16S rRNA Sequencing Reveals Relationship Between Potent Cellulolytic Genera and Feed Efficiency in the Rumen of Bulls. Front. Microbiol. 2018, 9, 1842. [Google Scholar] [CrossRef]
- Lin, L.; Xie, F.; Sun, D.; Liu, J.; Zhu, W.; Mao, S. Ruminal microbiome-host crosstalk stimulates the development of the ruminal epithelium in a lamb model. Microbiome 2019, 7, 83. [Google Scholar] [CrossRef]
- Ma, T.; Chen, D.-D.; Tu, Y.; Zhang, N.-F.; Si, B.-W.; Deng, K.-D.; Diao, Q.-Y. Effect of dietary supplementation with resveratrol on nutrient digestibility, methanogenesis and ruminal microbial flora in sheep. J. Anim. Physiol. Anim. Nutr. 2014, 99, 676–683. [Google Scholar] [CrossRef]
- Ma, T.; Wu, W.; Tu, Y.; Zhang, N.; Diao, Q. Resveratrol affects in vitro rumen fermentation, methane production and prokaryotic community composition in a time- and diet-specific manner. Microb. Biotechnol. 2020, 13, 1118–1131. [Google Scholar] [CrossRef] [Green Version]
- Lee, M. Official and standardized methods of analysis (3rd edn): Edited by C.A. Watson, Royal Society of Chemistry, 1994. £110.00 (xxiv + 778 pages) ISBN 0 85186 441 4. Trends Food Sci. Technol. 1995, 6, 382–383. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Chen, D.W.; He, J.; Zheng, P.; Yu, J.; Mao, X.B.; Huang, Z.Q.; Luo, Y.H.; Luo, J.Q.; Yu, B. Long-term dietary resveratrol supplementation decreased serum lipids levels, improved intramuscular fat content, and changed the expression of several lipid metabolism-related miRNAs and genes in growing-finishing pigs1. J. Anim. Sci. 2019, 97, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Benchaar, C.; Ametaj, B.; Beauchemin, K. Dose response to eugenol supplementation in growing beef cattle: Ruminal fermentation and intestinal digestion. Anim. Feed Sci. Technol. 2010, 158, 57–64. [Google Scholar] [CrossRef]
- Santra, A.; Karim, S.A. Rumen Manipulation to Improve Animal Productivity. Asian-Australas. J. Anim. Sci. 2003, 16, 748–763. [Google Scholar] [CrossRef]
- Valenzuela-Grijalva, N.V.; Pinelli-Saavedra, A.; Muhlia-Almazan, A.; Domínguez-Díaz, D.; González-Ríos, H. Dietary inclusion effects of phytochemicals as growth promoters in animal production. J. Anim. Sci. Technol. 2017, 59, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanapat, M.; Kang, S.; Polyorach, S. Development of feeding systems and strategies of supplementation to enhance rumen fermentation and ruminant production in the tropics. J. Anim. Sci. Biotechnol. 2013, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Fang, C.; Ma, Y.; He, S.; Ajuwon, K.M.; He, J. Dietary resveratrol supplement improves carcass traits and meat quality of Pekin ducks. Poult. Sci. 2021, 100, 100802. [Google Scholar] [CrossRef] [PubMed]
- Griñán-Ferré, C.; Bellver-Sanchis, A.; Izquierdo, V.; Corpas, R.; Roig-Soriano, J.; Chillón, M.; Andres-Lacueva, C.; Somogyvári, M.; Sőti, C.; Sanfeliu, C.; et al. The pleiotropic neuroprotective effects of resveratrol in cognitive decline and Alzheimer’s disease pathology: From antioxidant to epigenetic therapy. Ageing Res. Rev. 2021, 67, 101271. [Google Scholar] [CrossRef]
- Musial, C.; Siedlecka-Kroplewska, K.; Kmiec, Z.; Gorska-Ponikowska, M. Modulation of Autophagy in Cancer Cells by Dietary Polyphenols. Antioxidants 2021, 10, 123. [Google Scholar] [CrossRef]
- Chartrin, P.; Méteau, K.; Juin, H.; Bernadet, M.D.; Guy, G.; Larzul, C.; Rémignon, H.; Mourot, J.; Duclos, M.; Baéza, E. Effects of Intramuscular Fat Levels on Sensory Characteristics of Duck Breast Meat. Poult. Sci. 2006, 85, 914–922. [Google Scholar] [CrossRef]
- Zhao, J.X.; Li, Q.; Zhang, R.X.; Liu, W.Z.; Ren, Y.S.; Zhang, C.X.; Zhang, J.X. Effect of dietary grape pomace on growth performance, meat quality and antioxidant activity in ram lambs. Anim. Feed Sci. Technol. 2018, 236, 76–85. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Baas, T.J.; Malek, M.; Dekkers, J.C.M.; Prusa, K.; Rothschild, M.F. Correlations among selected pork quality traits. J. Anim. Sci. 2002, 80, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoseini, A.; Namazi, G.; Farrokhian, A.; Reiner, Ž.; Aghadavod, E.; Bahmani, F.; Asemi, Z. The effects of resveratrol on metabolic status in patients with type 2 diabetes mellitus and coronary heart disease. Food Funct. 2019, 10, 6042–6051. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.M.; Hegele, R.A. Functional foods and dietary supplements for the management of dyslipidaemia. Nat. Rev. Endocrinol. 2017, 13, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Xu, Y.; Mei, X.; Meng, Q.; Gao, Y.; Li, B.; Tu, Y. Antiobesity and lipid lowering effects of theaflavins on high-fat diet induced obese rats. J. Funct. Foods 2013, 5, 1142–1150. [Google Scholar] [CrossRef]
- Guo, X.-F.; Li, J.-M.; Tang, J.; Li, D. Effects of resveratrol supplementation on risk factors of non-communicable diseases: A meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2018, 58, 3016–3029. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, J.; Li, L.; Zheng, J.; Shi, Y.; Le, G. Regulatory effects of resveratrol on glucose metabolism and T-lymphocyte subsets in the development of high-fat diet-induced obesity in C57BL/6 mice. Food Funct. 2014, 5, 1452–1463. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, J.; Yu, B.; Chen, J.; Chen, D. Effects of resveratrol on lipid metabolism in muscle and adipose tissues: A reevaluation in a pig model. J. Funct. Foods 2015, 14, 590–595. [Google Scholar] [CrossRef]
- Kutbi, E.H.; Sohouli, M.H.; Fatahi, S.; Lari, A.; Shidfar, F.; Aljhdali, M.M.; Alhoshan, F.M.; Elahi, S.S.; Almusa, H.A.; Abu-Zaid, A. The beneficial effects of cinnamon among patients with metabolic diseases: A systematic review and dose-response meta-analysis of randomized-controlled trials. Crit. Rev. Food Sci. Nutr. 2021, 4, 1–19. [Google Scholar] [CrossRef]
- Le Lay, J.E.; Du, Q.; Mehta, M.B.; Bhagroo, N.; Hummer, B.T.; Falloon, J.; Carlson, G.; Rosenbaum, A.I.; Jin, C.; Kimko, H.; et al. Blocking endothelial lipase with monoclonal antibody MEDI5884 durably increases high density lipoprotein in nonhuman primates and in a phase 1 trial. Sci. Transl. Med. 2021, 13, eabb0602. [Google Scholar] [CrossRef]
- Lin, L.; Trabi, E.B.; Xie, F.; Mao, S. Comparison of the fermentation and bacterial community in the colon of Hu sheep fed a low-grain, non-pelleted, or pelleted high-grain diet. Appl. Microbiol. Biotechnol. 2021, 105, 2071–2080. [Google Scholar] [CrossRef]
- Barichella, M.; Severgnini, M.; Cilia, R.; Cassani, E.; Bolliri, C.; Caronni, S.; Ferri, V.; Cancello, R.; Ceccarani, C.; Faierman, S.; et al. Unraveling gut microbiota in Parkinson’s disease and atypical parkinsonism. Mov. Disord. 2019, 34, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Laffin, M.; Fedorak, R.; Zalasky, A.; Park, H.; Gill, A.; Agrawal, A.; Keshteli, A.; Hotte, N.; Madsen, K.L. A high-sugar diet rapidly enhances susceptibility to colitis via depletion of luminal short-chain fatty acids in mice. Sci. Rep. 2019, 9, 12294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truax, A.D.; Chen, L.; Tam, J.W.; Cheng, N.; Guo, H.; Koblansky, A.A.; Chou, W.-C.; Wilson, J.E.; Brickey, W.J.; Petrucelli, A.; et al. The Inhibitory Innate Immune Sensor NLRP12 Maintains a Threshold against Obesity by Regulating Gut Microbiota Homeostasis. Cell Host Microbe 2018, 24, 364–378.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vascellari, S.; Palmas, V.; Melis, M.; Pisanu, S.; Cusano, R.; Uva, P.; Perra, D.; Madau, V.; Sarchioto, M.; Oppo, V.; et al. Gut Microbiota and Metabolome Alterations Associated with Parkinson’s Disease. mSystems 2020, 5, e00561–e00520. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Chou, W.-C.; Lai, Y.; Liang, K.; Tam, J.W.; Brickey, W.J.; Chen, L.; Montgomery, N.D.; Li, X.; Bohannon, L.M.; et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites. Science 2020, 370, eaay9097. [Google Scholar] [CrossRef]
- Anderson, C.J.; Koester, L.R.; Schmitz-Esser, S. Rumen Epithelial Communities Share a Core Bacterial Microbiota: A Meta-Analysis of 16S rRNA Gene Illumina MiSeq Sequencing Datasets. Front. Microbiol. 2021, 12, 625400. [Google Scholar] [CrossRef]
- Fernández-Calleja, J.; Konstanti, P.; Swarts, H.J.M.; Bouwman, L.; Garcia-Campayo, V.; Billecke, N.; Oosting, A.; Smidt, H.; Keijer, J.; Van Schothorst, E.M. Non-invasive continuous real-time in vivo analysis of microbial hydrogen production shows adaptation to fermentable carbohydrates in mice. Sci. Rep. 2018, 8, 15351. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Li, F.; Li, C.; Li, G.; Zhang, D.; Song, Q.; Li, X.; Zhao, Y.; Wang, W. Characterization of the rumen microbiota and its relationship with residual feed intake in sheep. Animal 2021, 15, 100161. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, % | Content | Nutrition Levels, % | Content |
|---|---|---|---|
| Corn | 16.30 | Crude protein | 17.00 |
| Palm Kernel Expeller | 21.00 | Neutral detergent fiber | 43.52 |
| Peanut vine | 12.00 | Crude fat | 2.72 |
| Soybean meal | 3.00 | Calcium,Ca | 0.65 |
| Molasses | 2.00 | Total Phosphorus,TP | 0.43 |
| Manioc waste | 12.00 | Crude ash | 7.76 |
| Bagasse | 6.00 | Nitrogen free exteact | 51.2 |
| Corn skin | 4.00 | Acid detergent fiber | 22.65 |
| Cassava alcohol residue | 9.00 | ||
| rice mill by-product | 5.00 | ||
| Urea | 1.20 | ||
| Dicalcium phosphate | 0.40 | ||
| Limestone | 1.00 | ||
| Bentonite | 4.00 | ||
| Rumen protected fat | 1.00 | ||
| Premix feed | 2.10 | ||
| Total | 100.00 |
| Item | Control | 150 mg/kg | 300 mg/kg | 600 mg/kg |
|---|---|---|---|---|
| Final body weight (kg) | 45.09 ± 0.63 b | 48.90 ± 1.01 a | 44.67 ± 0.47 b | 38.04 ± 0.76 c |
| Initial body weight (kg) | 28.32 ± 0.50 | 28.43 ± 0.49 | 28.05 ± 0.45 | 28.20 ± 0.68 |
| ADG (g/d) | 139.75 ± 4.08 b | 170.59 ± 5.24 a | 138.44 ± 4.10 b | 82.06 ± 4.86 c |
| ADFI (kg/d) | 1.10 ± 0.09 a | 1.17 ± 0.11 a | 1.11 ± 0.14 a | 0.89 ± 0.07 b |
| FCR (kg/kg) | 7.87 ± 0.33 b | 6.99 ± 0.71 b | 7.48 ± 0.45 b | 9.20 ± 0.37 a |
| Hot carcass weight (kg) | 20.24 ± 0.43 b | 22.72 ± 1.00 a | 20.46 ± 0.53 b | 17.69 ± 0.51 c |
| Net meat weight (kg) | 7.44 ± 0.13 b | 8.62 ± 0.41 a | 7.32 ± 0.11 b | 5.71 ± 0.24 c |
| Dressing percentage (%) | 45.32 ± 0.98 | 45.77 ± 0.93 | 45.11 ± 1.36 | 46.50 ± 1.15 |
| Neat percentage (%) | 36.38 ± 0.43 b | 37.91 ± 0.45 a | 36.21 ± 0.32 b | 34.20 ± 0.72 c |
| LA (cm2) | 13.33 ± 0.36 b | 15.21 ± 0.34 a | 13.83 ± 0.37 b | 11.83 ± 0.54 c |
| Item | Control | 150 mg/kg | 300 mg/kg | 600 mg/kg |
|---|---|---|---|---|
| pH45min | 6.46 ± 0.18 | 6.49 ± 0.07 | 6.46 ± 0.08 | 6.70 ± 0.07 |
| pH24h | 6.00 ± 0.06 | 5.80 ± 0.06 | 5.96 ± 0.17 | 5.86 ± 0.15 |
| conductivity | 3.04 ± 0.07 | 3.28 ± 0.11 | 3.04 ± 0.06 | 3.17 ± 0.06 |
| Meat color: | ||||
| L* | 33.42 ± 0.43 | 33.00 ± 0.74 | 32.30 ± 0.85 | 32.80 ± 0.52 |
| a* | 7.32 ± 0.22b | 8.40 ± 0.27 a | 9.39 ± 0.49a | 8.92 ± 0.28 a |
| b* | 6.20 ± 0.21 | 6.66 ± 0.46 | 6.60 ± 0.35 | 6.75 ± 0.19 |
| Shear force(N) | 86.00 ± 4.10 a | 70.16 ± 2.55 b | 66.50 ± 2.01b | 63.23 ± 3.41 b |
| IMF(%) | 2.49 ± 0.10 b | 3.31 ± 0.34 a | 3.48 ± 0.17a | 4.13 ± 0.46 a |
| Item | Control | 150 mg/kg | 300 mg/kg | 600 mg/kg |
|---|---|---|---|---|
| TG (mmol/l) | 0.31 ± 0.01 a | 0.27 ± 0.01 b | 0.26 ± 0.02 b | 0.20 ± 0.02 c |
| TC (mmol/l) | 2.55 ± 0.14 | 2.52 ± 0.14 | 2.58 ± 0.13 | 2.50 ± 0.09 |
| HDL-C (mmol/l) | 0.96 ± 0.06 b | 1.36 ± 0.08 a | 1.49 ± 0.11 a | 1.45 ± 0.09 a |
| LDL-C (mmol/l) | 0.63 ± 0.13 a | 0.21 ± 0.08 b | 0.22 ± 0.07 b | 0.25 ± 0.06 b |
| HDL-C/TC (%) | 38.33 ± 4.00 b | 54.29 ± 3.34 a | 57.89 ± 3.38 a | 58.28 ± 3.51 a |
| Item | Control | 150 mg/kg | 300 mg/kg | 600 mg/kg |
|---|---|---|---|---|
| Chao1 | 263.31 ± 38.20 ab | 230.17 ± 12.65 b | 218.45 ± 40.32 ab | 280.51 ± 15.24 a |
| ACE | 249.24 ± 27.79 | 228.30 ± 14.09 | 209.50 ± 40.54 | 274.90 ± 16.12 |
| Shannon | 5.08 ± 0.41 | 5.27 ± 0.19 | 4.84 ± 0.39 | 5.25 ± 0.37 |
| Simpson | 0.92 ± 0.023 | 0.93 ± 0.020 | 0.92 ± 0.019 | 0.92 ± 0.011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Jiang, Y.; Zhang, S.; Zou, J.; Gao, X.; Song, Y.; Zhang, Y.; Hu, Y.; Huang, Y.; Jiang, Q. The Effect of Dietary Supplementation with Resveratrol on Growth Performance, Carcass and Meat Quality, Blood Lipid Levels and Ruminal Microbiota in Fattening Goats. Foods 2022, 11, 598. https://doi.org/10.3390/foods11040598
Shen Y, Jiang Y, Zhang S, Zou J, Gao X, Song Y, Zhang Y, Hu Y, Huang Y, Jiang Q. The Effect of Dietary Supplementation with Resveratrol on Growth Performance, Carcass and Meat Quality, Blood Lipid Levels and Ruminal Microbiota in Fattening Goats. Foods. 2022; 11(4):598. https://doi.org/10.3390/foods11040598
Chicago/Turabian StyleShen, Yujian, Yuhang Jiang, Sanbao Zhang, Juhong Zou, Xiaotong Gao, Ying Song, Yu Zhang, Yan Hu, Yanna Huang, and Qinyang Jiang. 2022. "The Effect of Dietary Supplementation with Resveratrol on Growth Performance, Carcass and Meat Quality, Blood Lipid Levels and Ruminal Microbiota in Fattening Goats" Foods 11, no. 4: 598. https://doi.org/10.3390/foods11040598
APA StyleShen, Y., Jiang, Y., Zhang, S., Zou, J., Gao, X., Song, Y., Zhang, Y., Hu, Y., Huang, Y., & Jiang, Q. (2022). The Effect of Dietary Supplementation with Resveratrol on Growth Performance, Carcass and Meat Quality, Blood Lipid Levels and Ruminal Microbiota in Fattening Goats. Foods, 11(4), 598. https://doi.org/10.3390/foods11040598