
Citrus Extract High in Flavonoids Beneficially Alters Intestinal Metabolic Responses in Subjects with Features of Metabolic Syndrome †

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro TIM-2 Study
2.1.1. Collection and Preparation of Fecal Samples
2.1.2. Study Product
2.1.3. Experimental Set-Up
2.1.4. Measurement of Short-Chain Fatty Acids and Branched-Chain Fatty Acids
2.2. Clinical Study
2.2.1. Participants
2.2.2. Study Design and Protocol
2.2.3. Measurement of Fecal Calprotectin
2.2.4. Measurement of Short-Chain Fatty Acids and Branched-Chain Fatty Acids
2.2.5. Analysis of Microbial Composition
2.2.6. Study Product
2.2.7. Statistical Analyses
3. Results
3.1. In Vitro TIM-2 Study
Production of Short-Chain Fatty Acids and Branched-Chain Fatty Acids
3.2. Clinical Study
3.2.1. Anthropometric Outcomes
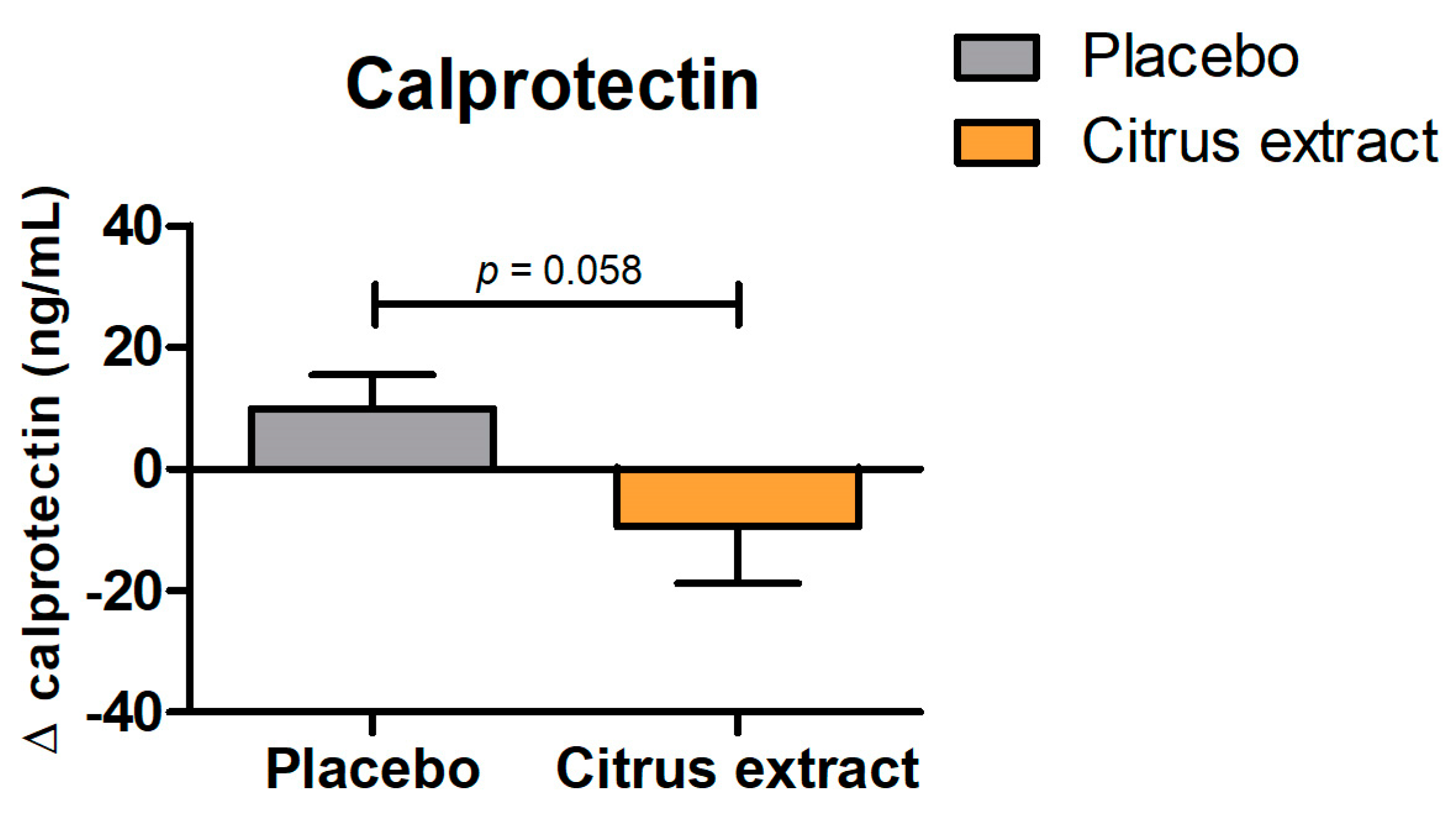
3.2.2. Calprotectin
3.2.3. Short-Chain Fatty Acids and Branched-Chain Fatty Acids
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Carrera-Bastos, P.; Fontes-Villalba, M.; O’Keefe, J.; Lindeberg, S.; Cordain, L. The western diet and lifestyle and diseases of civilization. Res. Rep. Clin. Cardiol. 2011, 2, 15–35. [Google Scholar] [CrossRef]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F. Enteric flora in health and disease. Digestion 2006, 73 (Suppl. 1), 5–12. [Google Scholar] [CrossRef] [PubMed]
- Mai, V.; Draganov, P.V. Recent advances and remaining gaps in our knowledge of associations between gut microbiota and human health. World J. Gastroenterol. 2009, 15, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: Major fermentation by-products and their impact on host health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- Dabke, K.; Hendrick, G.; Devkota, S. The gut microbiome and metabolic syndrome. J. Clin. Investig. 2019, 129, 4050–4057. [Google Scholar] [CrossRef] [PubMed]
- Sost, M.M.; Ahles, S.; Verhoeven, J.; Verbruggen, S.; Stevens, Y.; Venema, K. A Citrus Fruit Extract High in Polyphenols Beneficially Modulates the Gut Microbiota of Healthy Human Volunteers in a Validated In Vitro Model of the Colon. Nutrients 2021, 13, 3915. [Google Scholar] [CrossRef] [PubMed]
- Van Rymenant, E.; Salden, B.; Voorspoels, S.; Jacobs, G.; Noten, B.; Pitart, J.; Possemiers, S.; Smagghe, G.; Grootaert, C.; Van Camp, J. A Critical Evaluation of In Vitro Hesperidin 2S Bioavailability in a Model Combining Luminal (Microbial) Digestion and Caco-2 Cell Absorption in Comparison to a Randomized Controlled Human Trial. Mol. Nutr. Food Res. 2018, 62, e1700881. [Google Scholar] [CrossRef]
- Lima, A.C.D.; Cecatti, C.; Fidelix, M.P.; Adorno, M.A.T.; Sakamoto, I.K.; Cesar, T.B.; Sivieri, K. Effect of Daily Consumption of Orange Juice on the Levels of Blood Glucose, Lipids, and Gut Microbiota Metabolites: Controlled Clinical Trials. J. Med. Food 2019, 22, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Stevens, Y.; Rymenant, E.V.; Grootaert, C.; Camp, J.V.; Possemiers, S.; Masclee, A.; Jonkers, D. The Intestinal Fate of Citrus Flavanones and Their Effects on Gastrointestinal Health. Nutrients 2019, 11, 1464. [Google Scholar] [CrossRef]
- Venema, K. The TNO In Vitro Model of the Colon (TIM-2). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 293–304. [Google Scholar]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C., Jr.; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute scientific statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef]
- Aguirre, M.; Ramiro-Garcia, J.; Koenen, M.E.; Venema, K. To pool or not to pool? Impact of the use of individual and pooled fecal samples for in vitro fermentation studies. J. Microbiol. Methods 2014, 107, 1–7. [Google Scholar] [CrossRef]
- Yi, L.; Ma, S.; Ren, D. Phytochemistry and bioactivity of Citrus flavonoids: A focus on antioxidant, anti-inflammatory, anticancer and cardiovascular protection activities. Phytochem. Rev. 2017, 16, 479–511. [Google Scholar] [CrossRef]
- Cuevas-Tena, M.; Alegria, A.; Lagarda, M.J.; Venema, K. Impact of plant sterols enrichment dose on gut microbiota from lean and obese subjects using TIM-2 in vitro fermentation model. J. Funct. Foods 2019, 54, 164–174. [Google Scholar] [CrossRef]
- van Nuenen, M.H.M.C.; Diederick Meyer, P.; Venema, K. The Effect of Various Inulins andClostridium difficileon the Metabolic Activity of the Human Colonic Microbiotain vitro. Microb. Ecol. Health Dis. 2009, 15, 137–144. [Google Scholar] [CrossRef]
- van Nuenen, M.H.; Venema, K.; van der Woude, J.C.; Kuipers, E.J. The metabolic activity of fecal microbiota from healthy individuals and patients with inflammatory bowel disease. Dig. Dis. Sci. 2004, 49, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Salden, B.N.H. Nutritional Interventions Focusing on Gastrointestinal and Metabolic Health. Ph.D. Thesis, Maastricht University, Maastricht, The Netherlands, 2017. [Google Scholar]
- de Bock, M.; Derraik, J.G.; Brennan, C.M.; Biggs, J.B.; Morgan, P.E.; Hodgkinson, S.C.; Hofman, P.L.; Cutfield, W.S. Olive (Olea europaea L.) leaf polyphenols improve insulin sensitivity in middle-aged overweight men: A randomized, placebo-controlled, crossover trial. PLoS ONE 2013, 8, e57622. [Google Scholar] [CrossRef] [PubMed]
- De Boever, P.; Deplancke, B.; Verstraete, W. Fermentation by gut microbiota cultured in a simulator of the human intestinal microbial ecosystem is improved by supplementing a soygerm powder. J. Nutr. 2000, 130, 2599–2606. [Google Scholar] [CrossRef]
- Possemiers, S.; Verthe, K.; Uyttendaele, S.; Verstraete, W. PCR-DGGE-based quantification of stability of the microbial community in a simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 2004, 49, 495–507. [Google Scholar] [CrossRef]
- Boon, N.; Top, E.M.; Verstraete, W.; Siciliano, S.D. Bioaugmentation as a tool to protect the structure and function of an activated-sludge microbial community against a 3-chloroaniline shock load. Appl. Env. Microbiol. 2003, 69, 1511–1520. [Google Scholar] [CrossRef]
- Rinttila, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef]
- Xiong, R.-G.; Zhou, D.-D.; Wu, S.-X.; Huang, S.-Y.; Saimaiti, A.; Yang, Z.-J.; Shang, A.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Health Benefits and Side Effects of Short-Chain Fatty Acids. Foods 2022, 11, 2863. [Google Scholar] [CrossRef]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; van Harsselaar, J.; et al. Short chain fatty acids in human gut and metabolic health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Sahuri-Arisoylu, M.; Brody, L.P.; Parkinson, J.R.; Parkes, H.; Navaratnam, N.; Miller, A.D.; Thomas, E.L.; Frost, G.; Bell, J.D. Reprogramming of hepatic fat accumulation and ‘browning’ of adipose tissue by the short-chain fatty acid acetate. Int. J. Obes. 2016, 40, 955–963. [Google Scholar] [CrossRef]
- McNabney, S.M.; Henagan, T.M. Short Chain Fatty Acids in the Colon and Peripheral Tissues: A Focus on Butyrate, Colon Cancer, Obesity and Insulin Resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Feinle-Bisset, C.; Asarian, L.; Horowitz, M.; Beglinger, C.; Geary, N. Ghrelin, CCK, GLP-1, and PYY(3–36): Secretory Controls and Physiological Roles in Eating and Glycemia in Health, Obesity, and After RYGB. Physiol. Rev. 2017, 97, 411–463. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Zhou, J.; Wang, H.; Ding, Y.; Zhou, J.; Chong, P.H.; Zhu, L.; Ke, L.; Wang, X.; Rao, P.; et al. Effects of valerate on intestinal barrier function in cultured Caco-2 epithelial cell monolayers. Mol. Biol. Rep. 2022, 49, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Lent-Schochet, D.; Ramakrishnan, N.; McLaughlin, M.; Jialal, I. Metabolic syndrome is an inflammatory disorder: A conspiracy between adipose tissue and phagocytes. Clin. Chim. Acta 2019, 496, 35–44. [Google Scholar] [CrossRef]
- Charles-Messance, H.; Mitchelson, K.A.J.; De Marco Castro, E.; Sheedy, F.J.; Roche, H.M. Regulating metabolic inflammation by nutritional modulation. J. Allergy Clin. Immunol. 2020, 146, 706–720. [Google Scholar] [CrossRef]
- Stevens, Y.; de Bie, T.; Pinheiro, I.; Elizalde, M.; Masclee, A.; Jonkers, D. The effects of citrus flavonoids and their metabolites on immune-mediated intestinal barrier disruption using an in vitro co-culture model. Br. J. Nutr. 2022, 128, 1917–1926. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Control (SIEM) | Citrus Extract | |
|---|---|---|
| Total SCFAs (mmol) | 193.3 | 201.7 |
| Acetate (mmol) | 74.5 | 75.3 |
| Propionate (mmol) | 60.5 | 60.0 |
| Butyrate (mmol) | 53.4 | 61.2 |
| Valerate (mmol) | 2.8 | 4.2 |
| Caproate (mmol) | 2.1 | 1.0 |
| BCFAs (mmol) | 7.3 | 4.0 |
| Butyrate/total SCFA ratio | 0.28 | 0.30 |
| Total Population (n = 50) | Placebo (n = 27) | Citrus Extract (n = 23) | |
|---|---|---|---|
| Age, years (mean ± SD) | 51 ± 13 | 50 ± 14 | 52 ± 11 |
| Sex, Male/Female (n) | 18/32 | 9/18 | 9/14 |
| WHR (mean ± SD) | 0.93 ± 0.07 | 0.91 ± 0.07 | 0.94 ± 0.07 |
| BMI, kg/m2 (mean ± SD) | 30.8 ± 3.8 | 31.4 ± 4.2 | 30.0 ± 3.2 |
| BP systolic (mean ± SD) | 131 ± 13 | 131 ± 17 | 131 ± 7 |
| BP diastolic (mean ± SD) | 82 ± 9 | 82 ± 10 | 83 ± 8 |
| Placebo (n = 27) | Citrus Extract (n = 23) | P1 | P2 | |||||
|---|---|---|---|---|---|---|---|---|
| Baseline | 6 Weeks | 12 Weeks | Baseline | 6 Weeks | 12 Weeks | |||
| BMI (kg/m2) | 31.4 ± 0.73 | 31.4 ± 0.74 | 31.5 ± 0.77 | 30.0 ± 0.79 | 30.0 ± 0.80 | 30.0 ± 0.83 | 0.616 | 0.514 |
| WHR | 0.91 ± 0.014 | 0.91 ± 0.013 | 0.92 ± 0.013 | 0.94 ± 0.015 | 0.94 ± 0.013 | 0.93 ± 0.014 | 0.617 | 0.087 |
| Placebo (n = 27) | Citrus Extract (n = 23) | p Value | |||
|---|---|---|---|---|---|
| Baseline | 12 Weeks | Baseline | 12 Weeks | ||
| SCFAs and BCFAs | |||||
| Total SCFAs (µmol/g feces) | 56.7 ± 5.0 | 65.0 ± 5.9 | 69.3 ± 5.5 | 75.4 ± 6.4 | 0.792 |
| Acetate (µmol/g feces) | 32.9 ± 3.0 | 38.4 ± 3.4 | 39.8 ± 3.3 | 42.9 ± 3.6 | 0.625 |
| Propionate (µmol/g feces) | 11.1 ± 1.3 | 12.9 ± 1.6 | 13.9 ± 1.4 | 14.1 ± 1.7 | 0.465 |
| Butyrate (µmol/g feces) | 10.4 ± 1.1 | 10.9 ± 1.3 | 12.4 ± 1.2 | 15.0 ± 1.4 | 0.249 |
| Valerate (µmol/g feces) | 1.8 ± 0.2 | 2.1 ± 0.2 | 2.3 ± 0.2 | 2.4 ± 0.2 | 0.602 |
| Caproate (µmol/g feces) | 0.47 ± 0.2 | 0.63 ± 0.2 | 0.89 ± 0.2 | 0.92 ± 0.2 | 0.651 |
| BCFA (µmol/g feces) | 3.1 ± 0.3 | 3.2 ± 0.4 | 3.6 ± 0.4 | 3.9 ± 0.4 | 0.807 |
| Butyrate/ total SCFA ratio | 0.18 ± 0.01 | 0.17 ± 0.01 | 0.18 ± 0.01 | 0.19 ± 0.01 | 0.022 |
| Butyrate-producing bacteria | |||||
| E. rectale/C. coccoides (copies/g) × 1010 | 8.6 ± 1.3 | 9.5 ± 1.4 | 6.6 ± 1.4 | 9.5 ± 1.5 | 0.383 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurer Sost, M.; Stevens, Y.; Salden, B.; Troost, F.; Masclee, A.; Venema, K. Citrus Extract High in Flavonoids Beneficially Alters Intestinal Metabolic Responses in Subjects with Features of Metabolic Syndrome. Foods 2023, 12, 3413. https://doi.org/10.3390/foods12183413
Maurer Sost M, Stevens Y, Salden B, Troost F, Masclee A, Venema K. Citrus Extract High in Flavonoids Beneficially Alters Intestinal Metabolic Responses in Subjects with Features of Metabolic Syndrome. Foods. 2023; 12(18):3413. https://doi.org/10.3390/foods12183413
Chicago/Turabian StyleMaurer Sost, Mônica, Yala Stevens, Bouke Salden, Freddy Troost, Ad Masclee, and Koen Venema. 2023. "Citrus Extract High in Flavonoids Beneficially Alters Intestinal Metabolic Responses in Subjects with Features of Metabolic Syndrome" Foods 12, no. 18: 3413. https://doi.org/10.3390/foods12183413