


Antibacterial Activities of Agaricus bisporus Extracts and Their Synergistic Effects with the Antistaphylococcal Drug AFN-1252
 ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Extraction
2.3. Total Phenolic Content and Protein Content
2.4. Agar Well Diffusion Method
2.5. HPTLC Analysis
2.6. HPTLC Bioautography Assay
2.7. LC/MS Metabolite Identification
2.8. Bi-Therapy Assay
2.9. Image Processing and Multivariate Analysis
3. Results and Discussion
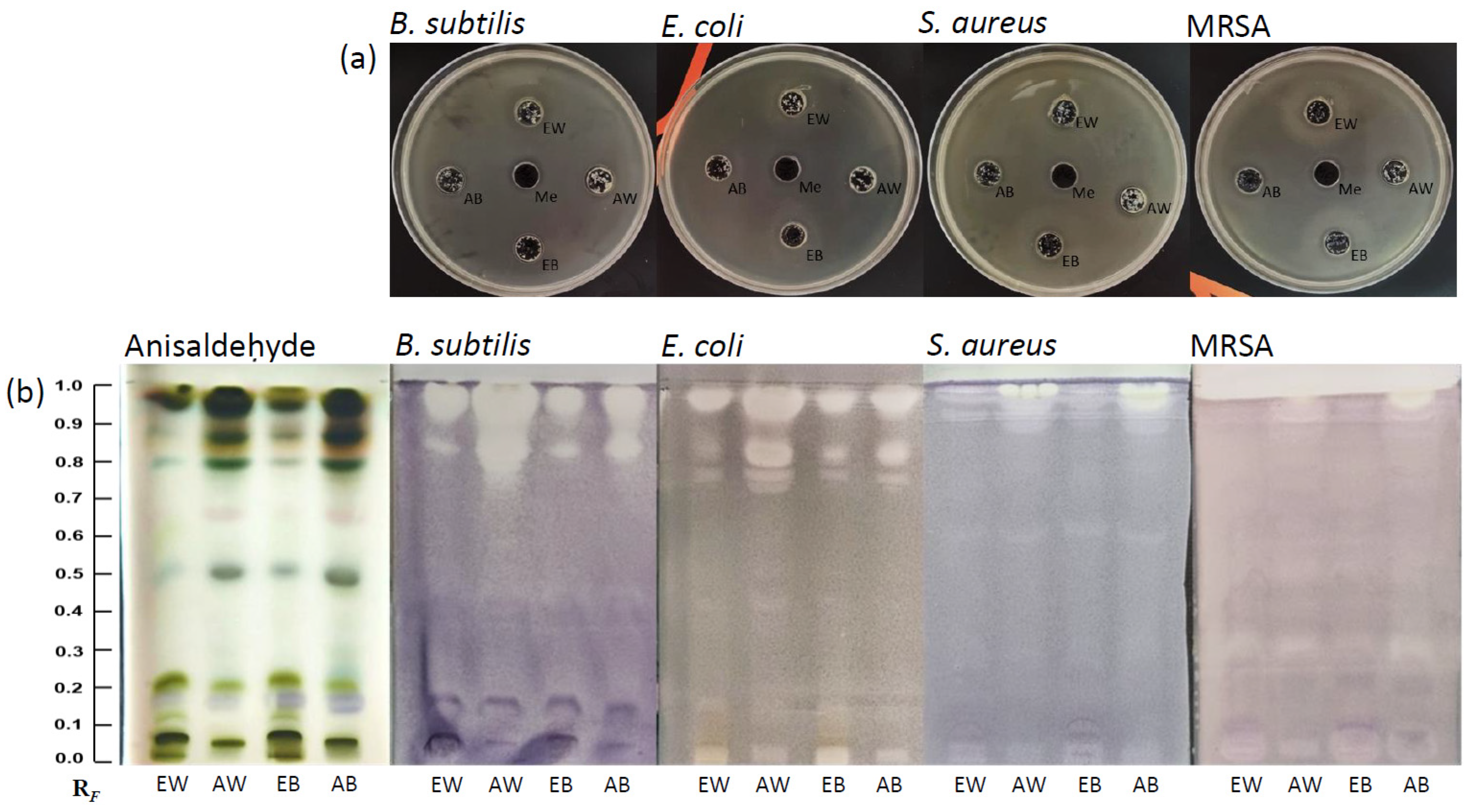
3.1. Extracts of A. bisporus and the Well Diffusion Assay
3.2. HPTLC Fingerprinting and HPTLC Bioassays
3.3. Principal Component Analysis
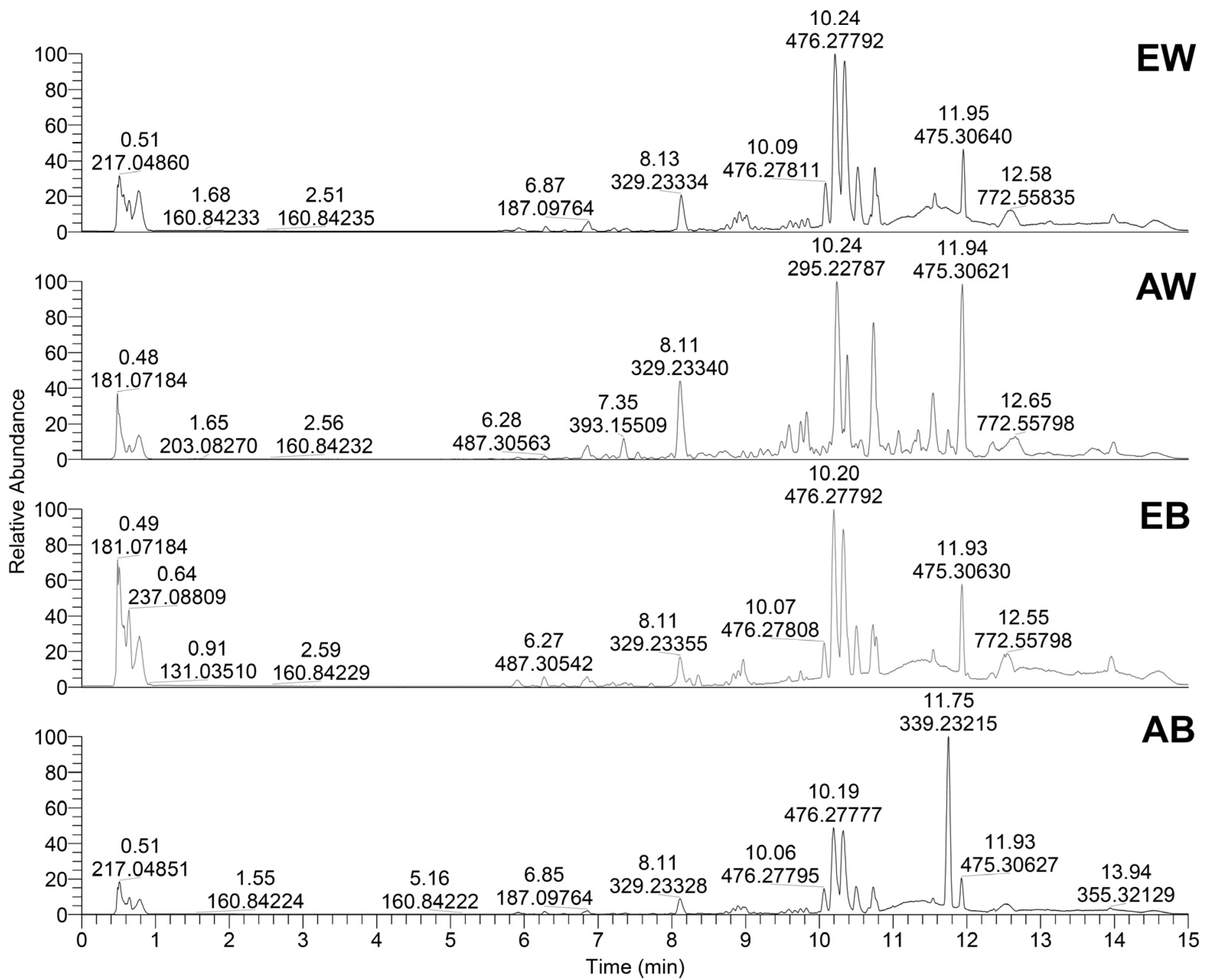
3.4. LC/MS Profiling of Metabolites
3.5. Synergistic Bi-Therapy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular mechanisms of antibiotic resistance revisited. Nat. Rev. Microbiol. 2023, 21, 280–295. [Google Scholar] [CrossRef]
- Mukherjee, R.; Vidic, J.; Auger, S.; Wen, H.-C.; Pandey, R.P.; Chang, C.-M. Exploring Disease Management and Control through Pathogen Diagnostics and One Health Initiative: A Concise Review. Antibiotics 2023, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Gunjan; Vidic, J.; Manzano, M.; Raj, V.S.; Pandey, R.P.; Chang, C.-M. Comparative meta-analysis of antimicrobial resistance from different food sources along with one health approach in Italy and Thailand. One Health 2023, 16, 100477. [Google Scholar] [CrossRef]
- Morvan, C.; Halpern, D.; Kénanian, G.; Pathania, A.; Anba-Mondoloni, J.; Lamberet, G.; Gruss, A.; Gloux, K. The Staphylococcus aureus FASII bypass escape route from FASII inhibitors. Biochimie 2017, 141, 40–46. [Google Scholar] [CrossRef]
- Gunjan; Himanshu; Mukherjee, R.; Vidic, J.; Manzano, M.; Leal, E.; Raj, V.S.; Pandey, R.P.; Chang, C.-M. Comparative meta-analysis of antimicrobial resistance from different food sources along with one health approach in the Egypt and UK. BMC Microbiol. 2023, 23, 291. [Google Scholar] [CrossRef]
- Anusiya, G.; Prabu, U.G.; Yamini, N.; Sivarajasekar, N.; Rambabu, K.; Bharath, G.; Banat, F. A review of the therapeutic and biological effects of edible and wild mushrooms. Bioengineered 2021, 12, 11239–11268. [Google Scholar] [CrossRef]
- Li, H.; Tian, Y.; Menolli, N.; Ye, L.; Karunarathna, S.C.; Perez-Moreno, J.; Rahman, M.M.; Rashid, H.; Phengsintham, P.; Rizal, L.; et al. Reviewing the world’s edible mushroom species: A new evidence-based classification system. Comp. Rev. Food Sci. Food Saf. 2021, 20, 1982–2014. [Google Scholar] [CrossRef]
- Ndungutse, V.; Mereddy, R.; Sultanbawa, Y. Bioactive Properties of Mushroom (Agaricus bisporus) Stipe Extracts: Functional Properties of Mushroom Stipe Extracts. J. Food Process. Preserv. 2015, 39, 2225–2233. [Google Scholar] [CrossRef]
- Ejikeme, N.; Henrietta, O.U. Antimicrobial activity of some local mushrooms on pathogenic isolates. J. Med. Plants Res. 2010, 4, 2460–2465. [Google Scholar] [CrossRef]
- Risan, M.H.; Taemor, S.H.; Muhsin, A.H.; Hussan, S. Antibacterial activity of Agaricus bisporus and Pleurotus ostreatus extracts against some gram negative and positive bacteria. Eur. J. Biomed. 2017, 4, 9–15. [Google Scholar]
- Huguet, C.; Bourjot, M.; Bellanger, J.-M.; Prévost, G.; Urbain, A. Screening for Antibacterial Activity of French Mushrooms against Pathogenic and Multidrug Resistant Bacteria. Appl. Sci. 2022, 12, 5229. [Google Scholar] [CrossRef]
- Chang, S.T.; Miles, P.G.; Chang, S.T. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Jeong, S.C.; Koyyalamudi, S.R.; Jeong, Y.T.; Song, C.H.; Pang, G. Macrophage Immunomodulating and Antitumor Activities of Polysaccharides Isolated from Agaricus bisporus White Button Mushrooms. J. Med. Food 2012, 15, 58–65. [Google Scholar] [CrossRef]
- Liu, J.; Jia, L.; Kan, J.; Jin, C.-H. In vitro and in vivo antioxidant activity of ethanolic extract of white button mushroom (Agaricus bisporus). Food Chem. Toxicol. 2013, 51, 310–316. [Google Scholar] [CrossRef]
- Moro, C.; Palacios, I.; Lozano, M.; D’Arrigo, M.; Guillamón, E.; Villares, A.; Martínez, J.A.; García-Lafuente, A. Anti-inflammatory activity of methanolic extracts from edible mushrooms in LPS activated RAW 264.7 macrophages. Food Chem. 2012, 130, 350–355. [Google Scholar] [CrossRef]
- Muszynska, B.; Grzywacz, A.; Kala, K.; Gdula-Argasinska, J. Anti-Inflammatory Potential of In Vitro Cultures of the White Button Mushroom, Agaricus bisporus (Agaricomycetes), in Caco-2 Cells. Int. J. Med. Mushrooms 2018, 20, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Natić, M.M.; Dabić, D.Č.; Papetti, A.; Fotirić Akšić, M.M.; Ognjanov, V.; Ljubojević, M.; Tešić, Ž.L. Analysis and characterisation of phytochemicals in mulberry (Morus alba L.) fruits grown in Vojvodina, North Serbia. Food Chem. 2015, 171, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Cvetanovic, A.; Gašić, U.; Dragićević, M.; Stupar, A.; Uysal, A.; Şenkardes, I.; Sinan, K.I.; Picot-Allain, M.C.N.; Ak, G.; et al. UHPLC-LTQ OrbiTrap MS analysis and biological properties of Origanum vulgare subsp. viridulum obtained by different extraction methods. Ind. Crop. Prod. 2020, 154, 112747. [Google Scholar] [CrossRef]
- Kumar, V.; Prasher, I.B. Phytochemical Analysis and Antioxidant Activity of Endophytic Fungi Isolated from Dillenia indica Linn. Appl. Biochem. Biotechnol. 2024, 196, 332–349. [Google Scholar] [CrossRef]
- Morosanova, M.A.; Fedorova, T.V.; Polyakova, A.S.; Morosanova, E.I. Agaricus bisporus Crude Extract: Characterization and Analytical Application. Molecules 2020, 25, 5996. [Google Scholar] [CrossRef]
- Ye, L.; Zhang, B.; Yang, X.; Huang, Y.; Luo, J.; Zhang, X.; Tan, W.; Song, C.; Ao, Z.; Shen, C.; et al. Metabolomic profiling reveals biomarkers for diverse flesh colors in jelly fungi (Auricularia cornea). Food Chem. 2024, 446, 138906. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Ren, Y.; Hua, X.; Zhang, J.; Zhang, B.; Dong, J.; Efferth, T.; Ma, P. Phytochemistry and bioactivities of the main constituents of Polyporus umbellatus (Pers.) Fries. Phytomedicine 2022, 103, 154196. [Google Scholar] [CrossRef]
- Iida, A.; Sanekata, M.; Wada, S.-I.; Fujita, T.; Tanaka, H.; Enoki, A.; Fuse, G.; Kanai, M.; Asami, K. Fungal Metabolites. XVIII. New Membrane-Modifying Peptides, Trichorozins I–IV, from the Fungus Trichoderma harzianum. Chem. Pharm. Bull. 1995, 43, 392–397. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Seguin, P.; Ahn, J.-K.; Kim, J.-J.; Chun, S.-C.; Kim, E.-H.; Seo, S.-H.; Kang, E.-Y.; Kim, S.-L.; Park, Y.-J.; et al. Phenolic Compound Concentration and Antioxidant Activities of Edible and Medicinal Mushrooms from Korea. J. Agric. Food Chem. 2008, 56, 7265–7270. [Google Scholar] [CrossRef]
- Palacios, I.; Lozano, M.; Moro, C.; D’arrigo, M.; Rostagno, M.; Martínez, J.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Boonsong, S.; Klaypradit, W.; Wilaipun, P. Antioxidant activities of extracts from five edible mushrooms using different extractants. Agric. Nat. Resour. 2016, 50, 89–97. [Google Scholar] [CrossRef]
- Houshdar Tehrani, M.H.; Fakhrehoseini, E.; Kamali Nejad, M.; Mehregan, H.; Hakemi-Vala, M. Search for Proteins in the Liquid Extract of Edible Mushroom, Agaricus bisporus, and Studying their Antibacterial Effects. Iran. J. Pharm. Res. 2012, 11, 145–150. [Google Scholar]
- Wang, H.; Ng, T. Isolation of a Novel Ubiquitin-like Protein from Pleurotus ostreatus Mushroom with Anti-Human Immunodeficiency Virus, Translation-Inhibitory, and Ribonuclease Activities. Biochem. Biophys. Res. Commun. 2000, 276, 587–593. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Kustrin, E.; Gegechkori, V.; Morton, D.W. High-Performance Thin-Layer Chromatography Hyphenated with Microchemical and Biochemical Derivatizations in Bioactivity Profiling of Marine Species. Mar. Drugs 2019, 17, 148. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Shi, Y.; Zhai, Y.; Ren, J.; Cai, L.; Sun, L.; Liu, L. Penindolacid A, a new indole alkaloid from the marine-derived fungus Penicillium sp. Magn. Reson. Chem. 2023, 61, 554–559. [Google Scholar] [CrossRef]
- Gu, H.; Lin, Y.; Qin, J.; Jia, X.; Wang, W.; Yin, H. Characterization of heteropolysaccharides from Rhizoctonia solani AG1 IA cell wall and comparison of their effect on inducing plant defense. Int. J. Biol. Macromol. 2024, 254, 127747. [Google Scholar] [CrossRef]
- Latif, A.; Issa Khan, M.; Rakha, A.; Ali Khan, J. Evaluating the therapeutic potential of white button mushroom (Agaricus bisporus) against DMBA-induced breast cancer in Sprague Dawley rats. J. Food Biochem. 2021, 45, e13979. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Miao, J.; Huang, L.; Li, B. Microwave freeze-drying of button mushroom (Agaricus bisporus) based on non-volatile taste components by controlling microwave power density. Int. J. Food Sci. Technol. 2022, 57, 379–389. [Google Scholar] [CrossRef]
- Delgado-Povedano, M.D.M.; De Medina, V.S.; Bautista, J.; Priego-Capote, F.; De Castro, M.D.L. Tentative identification of the composition of Agaricus bisporus aqueous enzymatic extracts with antiviral activity against HCV: A study by liquid chromatography–tandem mass spectrometry in high resolution mode. J. Funct. Foods 2016, 24, 403–419. [Google Scholar] [CrossRef]
- Chuluunbaatar, B.; Béni, Z.; Dékány, M.; Kovács, B.; Sárközy, A.; Datki, Z.; Mácsai, L.; Kálmán, J.; Hohmann, J.; Ványolós, A. Triterpenes from the Mushroom Hypholoma lateritium: Isolation, Structure Determination and Investigation in Bdelloid Rotifer Assays. Molecules 2019, 24, 301. [Google Scholar] [CrossRef] [PubMed]
- Yingying, D.; Shicheng, H.; Yingchun, J. Physiological evidence for the relationship between unsaturated fatty acid and cell permeability in extracellular polysaccharide synthesis and secretion Agaricus bitorquis (Quél.) Sacc. Chaidam. Indian J. Biochem. Biophys. (IJBB) 2024, 61, 166–181. [Google Scholar] [CrossRef]
- Wadman, M.W.; van Zadelhoff, G.; Hamberg, M.; Visser, T.; Veldink, G.A.; Vliegenthart, J.F.G. Conversion of linoleic acid into novel oxylipins by the mushroom Agaricus bisporus. Lipids 2005, 40, 1163–1170. [Google Scholar] [CrossRef]
- Liuqing, W.; Qiuhui, H.; Fei, P.; Mugambi, M.A.; Wenjian, Y. Influence of different storage conditions on physical and sensory properties of freeze-dried Agaricus bisporus slices. LWT 2018, 97, 164–171. [Google Scholar] [CrossRef]
- Pandohee, J.; Stevenson, P.G.; Conlan, X.A.; Zhou, X.-R.; Jones, O.A.H. Off-line two-dimensional liquid chromatography for metabolomics: An example using Agaricus bisporus mushrooms exposed to UV irradiation. Metabolomics 2015, 11, 939–951. [Google Scholar] [CrossRef]
- Rilda, Y.; Khairu Ummah, K.; Septiani, U.; Syukri, S.; Agustien, A.; Pardi, H.; Sofyan, N. Biosynthesis of Zinc oxide nanorods using Agaricus bisporus and its antibacterial capability enhancement with dodeciltriethoxyl on cotton textiles. Mater. Sci. Eng. B 2023, 298, 116910. [Google Scholar] [CrossRef]
- Wickramasinghe, M.A.; Nadeeshani, H.; Sewwandi, S.M.; Rathnayake, I.; Kananke, T.C.; Liyanage, R. Comparison of nutritional composition, bioactivities, and FTIR- ATR microstructural properties of commercially grown four mushroom species in Sri Lanka; Agaricus bisporus, Pleurotus ostreatus, Calocybe sp. (MK-white), Ganoderma lucidum. Food Prod. Process Nutr. 2023, 5, 43. [Google Scholar] [CrossRef]
- Câmara Neto, J.F.; Campelo, M.d.S.; Cerqueira, G.S.; De Miranda, J.A.L.; Guedes, J.A.C.; De Almeida, R.R.; Soares, S.D.A.; Gramosa, N.V.; Zocolo, G.J.; Vieira, G.P.; et al. Gastroprotective effect of hydroalcoholic extract from Agaricus blazei Murill against ethanol-induced gastric ulcer in mice. J. Ethnopharmacol. 2022, 292, 115191. [Google Scholar] [CrossRef] [PubMed]
- Zhai, F.-H.; Chen, Y.-F.; Zhang, Y.; Zhao, W.-J.; Han, J.-R. Phenolic compounds and antioxidant properties of wheat fermented with Agaricus brasiliensis and Agaricus bisporus. FEMS Microbiol. Lett. 2021, 368, fnaa213. [Google Scholar] [CrossRef] [PubMed]
- Nagy, M.; Socaci, S.A.; Dulf, F.V.; Diaconeasa, Z.; Farcas, A.C.; Tofana, M.; Semeniuc, C.A. Bioactive Compounds and Volatile Profiles of Five Transylvanian Wild Edible Mushrooms. Molecules 2018, 23, 3272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.; Atlagić, K.; Novaković, A.; Šibul, F.; Živić, M.; Stevanović, K.; Pejin, B. Fatty Acids Predominantly Affect Anti-Hydroxyl Radical Activity and FRAP Value: The Case Study of Two Edible Mushrooms. Antioxidants 2019, 8, 480. [Google Scholar] [CrossRef] [PubMed]
- Casillas-Vargas, G.; Ocasio-Malavé, C.; Medina, S.; Morales-Guzmán, C.; Del Valle, R.G.; Carballeira, N.M.; Sanabria-Ríos, D.J. Antibacterial fatty acids: An update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Progress. Lipid Res. 2021, 82, 101093. [Google Scholar] [CrossRef]
- Kumar, P.; Lee, J.-H.; Beyenal, H.; Lee, J. Fatty Acids as Antibiofilm and Antivirulence Agents. Trends Microbiol. 2020, 28, 753–768. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Yield of Extraction (mg/g) | Total Phenolic Content (mg/mL) | Total Protein Content (mg/mL) |
|---|---|---|---|
| EW | 51.40 ± 5.5 | 0.51 ± 0.05 | 1.02 ± 0.3 |
| EB | 60.80 ± 5.2 | 0.49 ± 0.04 | 1.33 ± 0.12 |
| AW | 8.46 ± 0.65 | 0.54 ± 0.06 | 2.38 ± 0.15 |
| AB | 28.78 ± 5.33 | 0.29 ± 0.09 | 1.00 ± 0.23 |
| No | Compound Name | tR, min | Molecular Formula, [M–H]– | Calculated Mass, m/z | Exact Mass, m/z | Δ ppm | MS2 Fragments, (% Base Peak) | Ref |
|---|---|---|---|---|---|---|---|---|
| Phenolic acids | ||||||||
| 1 | Gallic acid | 0.68 | C7H5O5– | 169.01425 | 169.01429 | −0.28 | 125.02459 (100), 169.01451 (42) | [19] |
| 2 | Dihydroxybenzoic acid | 1.12 | C7H5O4– | 153.01933 | 153.01941 | −0.53 | 109.02964 (100), 153.01941 (48) | [30] |
| 3 | Hydroxybenzoic acid 1 | 2.61 | C7H5O3– | 137.02442 | 137.02459 | −1.25 | 93.03468 (100), 137.02461 (29) | [31] |
| 4 | Benzoic acid | 4.93 | C7H5O2– | 121.02950 | 121.02966 | −1.31 | 121.02971 (100) | [32] |
| 5 | Caffeic acid | 5.25 | C9H7O4– | 179.03498 | 179.03513 | −0.80 | 135.04564 (100), 179.03488 (7) | [20] |
| 6 | p-Coumaric acid | 6.04 | C9H7O3– | 163.04007 | 163.04021 | −0.89 | 119.05042 (100) | [19] |
| 7 | Hydroxybenzoic acid 2 | 6.77 | C7H5O3– | 137.02442 | 137.02457 | −1.08 | 93.03466 (100), 137.02438 (68) | [31] |
| Amino acids | ||||||||
| 8 | L-Threonine | 0.49 | C4H8NO3– | 118.05100 | 118.05109 | −0.76 | 74.02480 (100), 118.05112 (34) | [33] |
| 9 | D-Asparagine | 0.50 | C4H7N2O3– | 131.04622 | 131.04629 | −0.53 | 69.03466 (4), 87.04523 (100), 113.02460 (9) | [33] |
| 10 | L-Glutamic acid | 0.52 | C5H8NO4– | 146.04588 | 146.04602 | −0.94 | 102.05613 (100), 128.03555 (55), 146.04623 (51) | [33] |
| 11 | L-Aspartic acid | 0.53 | C4H6NO4– | 132.03023 | 132.03039 | −1.18 | 71.01395 (41), 115.00387 (100 | [33] |
| 12 | D-Valine | 0.53 | C5H10NO2– | 116.07170 | 116.07186 | −1.35 | 73.02954 (57), 99.00883 (6), 116.07183 (100) | [33] |
| 13 | L-Pyroglutamic acid | 0.64 | C5H6NO3– | 128.03532 | 128.03542 | −0.78 | 128.03543 (100) | [33] |
| 14 | L-Glutamyl-L-leucine | 0.65 | C11H19N2O5– | 259.12995 | 259.13007 | −0.46 | 127.05146 (11), 128.03622 (50), 130.08778 (100), 197.13062 (22), 241.11896 (17) | [33] |
| 15 | Tyrosine | 0.65 | C9H10NO3– | 180.06662 | 180.06673 | −0.62 | 101.02451 (28), 119.05041 (51), 136.07693 (8), 163.04037 (92), 180.06688 (100) | [33] |
| 16 | D-α-Aminoadipic acid | 0.65 | C6H10NO4– | 160.06153 | 160.06173 | −1.25 | 99.04529 (100), 101.02456 (40), 116.03542 (23), 143.03522 (10) | [21] |
| 17 | D-Phenylalanine | 0.77 | C9H10NO2– | 164.07170 | 164.07182 | −0.73 | 72.00916 (35), 91.05540 (6), 103.05539 (6), 147.04532 (100) | [21] |
| Fatty acids | ||||||||
| 18 | 9,10,13-Trihydroxy-11-octadecenoic acid | 8.13 | C18H33O5– | 329.23335 | 329.23352 | −0.52 | 139.11314 (24), 171.10291 (67), 211.13429 (57), 229.14474 (42), 329.23380 (100) | [34] |
| 19 | 8-Hydroxy-13-oxo-9,11-octadecadienoic acid | 9.06 | C18H29O4– | 309.20713 | 309.20733 | −0.63 | 113.09734 (70), 171.10291 (63), 195.10283 (100), 291.19696 (6) | [35] |
| 20 | 5,8-Dihydroxy-9,12-octadecadienoic acid | 9.58 | C18H31O4– | 311.22278 | 311.22298 | −0.64 | 171.10292 (100), 197.11871 (40), 211.13425 (86), 275.20178 (14), 293.21259 (80) | [34] |
| 21 | Linolenic acid | 10.06 | C18H29O2– | 277.21730 | 277.21741 | −0.37 | 134.03749 (34), 233.15533 (5), 277.21829 (100) | [36] |
| 22 | 8-Hydroxy-9,12-octadecadienoic acid | 10.25 | C18H31O3– | 295.22787 | 295.22793 | −0.21 | 171.10284 (54), 195.1387 (9), 277.21747 (100), 295.22815 (70) | [37] |
| 23 | 16-Hydroxyhexadecanoic acid | 11.56 | C16H31O3– | 271.22787 | 271.22802 | −0.56 | 225.22264 (83), 271.22809 (100) | [36] |
| 24 | Linoleic acid | 11.93 | C18H31O2– | 279.23295 | 279.23309 | −0.47 | 279.23315 (100) | [36] |
| 25 | 2-Hydroxystearic acid | 12.32 | C18H35O3– | 299.25917 | 299.25930 | −0.44 | 253.25391 (69), 281.24927 (2), 299.25934 (100) | [21] |
| Steroids | ||||||||
| 26 | Unknown steroid 1 | 9.80 | C29H47O7– | 507.33273 | 507.33273 | −0.01 | 387.29028 (100), 428.29233 (10) | NA |
| 27 | Polyporusterone G | 10.73 | C28H43O5– | 459.31160 | 459.31166 | −0.13 | 325.25391 (100) | [22] |
| 28 | Unknown steroid 2 | 11.94 | C28H43O6– | 475.30651 | 475.30667 | −0.33 | 97.02962 (11), 315.26965 (19), 369.31674 (13), 431.31708 (100) | NA |
| Peptides | ||||||||
| 29 | Benzyl-2-[(1-hydroxy-4-methylpentan-2-yl)-carbamoyl]-pyrrolidine-1-carboxylate | 9.62 | C19H27N2O4– | 347.19763 | 347.19843 | −2.31 | 166.01128 (13), 171.10306 (41), 211.13458 (19), 293.21323 (45), 311.22311 (100) | [23] |
| 30 | 2-Methyl-N-[N-[N-[(phenylmethoxy)-carbonyl]-isoleucyl]-leucyl]-alanine methyl ester | 10.20 | C25H38N3O6– | 476.27660 | 476.27778 | −2.47 | 196.03815 (11), 279.23309 (100) | [23] |
| Other metabolites | ||||||||
| 31 | Succinic acid | 0.50 | C4H5O4– | 117.01930 | 117.01948 | −1.56 | 73.02953 (100), 99.00886 (11), 117.01947 (45) | [38] |
| 32 | Maleic acid | 0.52 | C4H3O4– | 115.00368 | 115.00381 | −1.15 | 71.01392 (100), 115.00381 (18) | [38] |
| 33 | Oxaceprol | 0.73 | C7H10NO4– | 172.06153 | 172.06166 | −0.77 | 96.04548 (26), 140.03525 (8), 172.06171 (100) | [38] |
| 34 | Glutaric acid | 0.90 | C5H7O4– | 131.03498 | 131.03515 | −1.30 | 87.04521 (100), 113.02464 (7), 131.03514 (41) | [39] |
| 35 | Adipic acid | 1.37 | C6H9O4– | 145.05063 | 145.05081 | −1.20 | 83.05041 (6), 101.06094 (100), 145.05069 (29) | [40] |
| 36 | Hexanoic acid | 6.44 | C6H11O2– | 115.07645 | 115.07658 | −1.13 | 71.01392 (54), 115.07677 (100) | [41] |
| 37 | Agaritine | 6.53 | C12H16N3O4– | 266.11463 | 266.11481 | −0.68 | 74.05275 (19), 83.10571 (19), 128.03564 (100), 248.10542 (21) | [42] |
| 38 | Indole-2-carboxylic acid | 6.56 | C9H6NO2– | 160.04040 | 160.04055 | −0.93 | 116.05077 (67), 160.03983 (100) | [43] |
| 39 | Azelaic acid | 7.14 | C9H15O4– | 187.09758 | 187.09772 | −0.72 | 59.01388 (20), 87.00887 (31), 99.08166 (12), 125.09732 (100) | [44] |
| 40 | Penipacid C | 7.34 | C10H9N2O4– | 221.05678 | 221.05707 | −1.29 | 92.05076 (36), 136.04056 (100) | [30] |
| 41 | Strobilactone A | 9.04 | C15H21O4– | 265.14453 | 265.14485 | −1.21 | 203.14365 (6), 221.15497 (100), 247.13522 (23), 265.14548 (82) | [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankov, M.; Léguillier, V.; Gašić, U.; Anba-Mondoloni, J.; Ristivojević, M.K.; Radoičić, A.; Dimkić, I.; Ristivojević, P.; Vidic, J. Antibacterial Activities of Agaricus bisporus Extracts and Their Synergistic Effects with the Antistaphylococcal Drug AFN-1252. Foods 2024, 13, 1715. https://doi.org/10.3390/foods13111715
Jankov M, Léguillier V, Gašić U, Anba-Mondoloni J, Ristivojević MK, Radoičić A, Dimkić I, Ristivojević P, Vidic J. Antibacterial Activities of Agaricus bisporus Extracts and Their Synergistic Effects with the Antistaphylococcal Drug AFN-1252. Foods. 2024; 13(11):1715. https://doi.org/10.3390/foods13111715
Chicago/Turabian StyleJankov, Milica, Vincent Léguillier, Uroš Gašić, Jamila Anba-Mondoloni, Maja Krstić Ristivojević, Aleksandra Radoičić, Ivica Dimkić, Petar Ristivojević, and Jasmina Vidic. 2024. "Antibacterial Activities of Agaricus bisporus Extracts and Their Synergistic Effects with the Antistaphylococcal Drug AFN-1252" Foods 13, no. 11: 1715. https://doi.org/10.3390/foods13111715
APA StyleJankov, M., Léguillier, V., Gašić, U., Anba-Mondoloni, J., Ristivojević, M. K., Radoičić, A., Dimkić, I., Ristivojević, P., & Vidic, J. (2024). Antibacterial Activities of Agaricus bisporus Extracts and Their Synergistic Effects with the Antistaphylococcal Drug AFN-1252. Foods, 13(11), 1715. https://doi.org/10.3390/foods13111715