
Synthesis of Ganoderic Acids Loaded Zein-Chitosan Nanoparticles and Evaluation of Their Hepatoprotective Effect on Mice Given Excessive Alcohol


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Synthesization of GA-NPs
2.3. Nanoparticle Characteristics
2.3.1. Particle Size, Polydispersity Index (PDI) and ζ-Potential Measurement
2.3.2. Encapsulation Efficiency and Loading Efficiency Determination
2.3.3. Scanning Electron Microscopy (SEM) Analysis
2.3.4. Fourier Transform Infrared (FT-IR) Spectroscopy
2.4. Animal Treatment and Sample Collection
2.5. Biochemical Assays of the Serum and Liver Samples
2.6. Histopathologic Evaluation of Liver and Jejunum Tissues
2.7. Determination of Short-Chain Fatty Acids (SCFAs) in Fecal Samples
2.8. High Throughput Sequencing of 16S rDNA Amplicon
2.9. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effects of CS Concentration on GA-NPs Characteristics
3.2. Morphological Observation of GA-NPs
3.3. FT-IR Characterization of GA-NPs
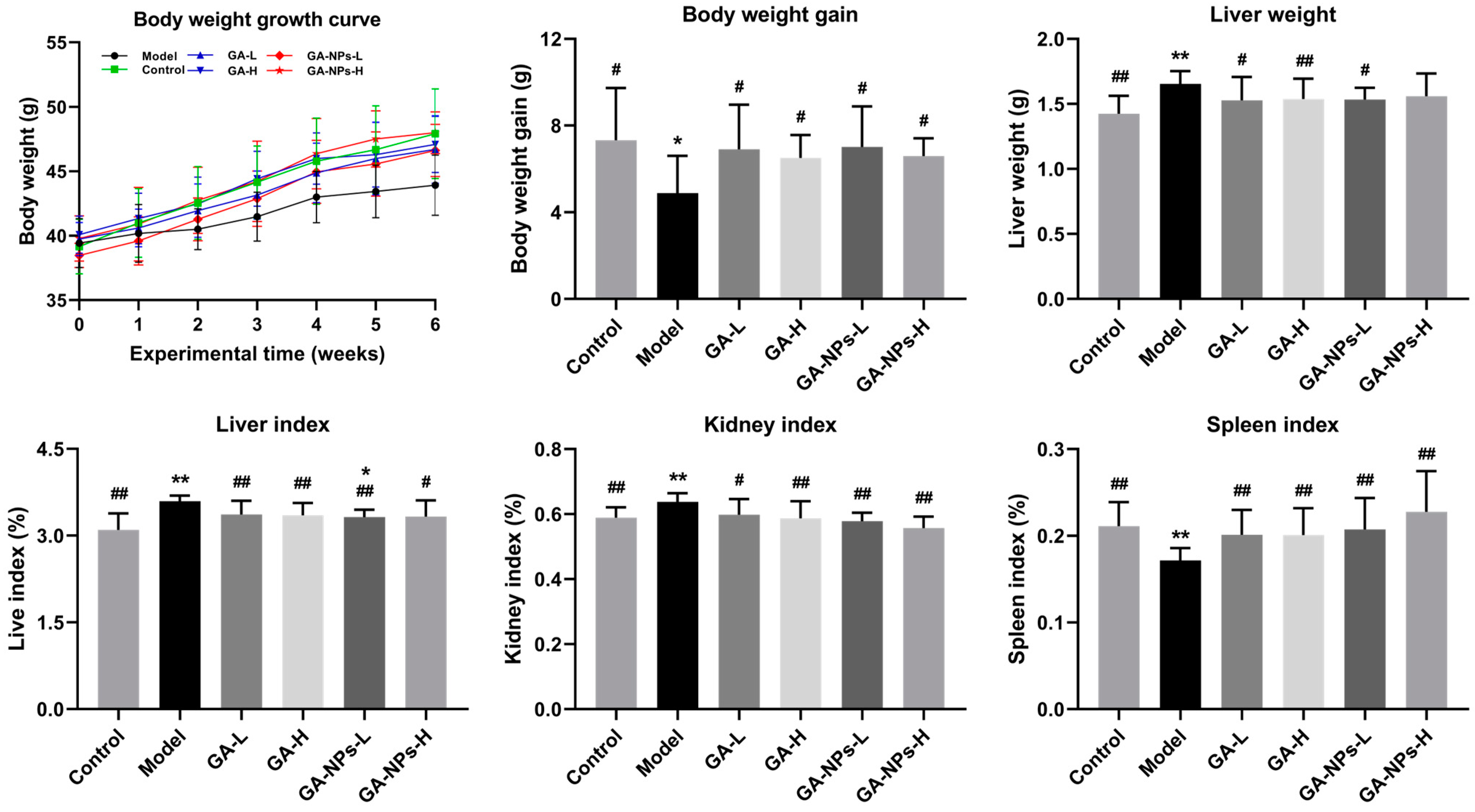
3.4. Effects of GA-NPs Intervention on the Body Weight and Organ Indexes
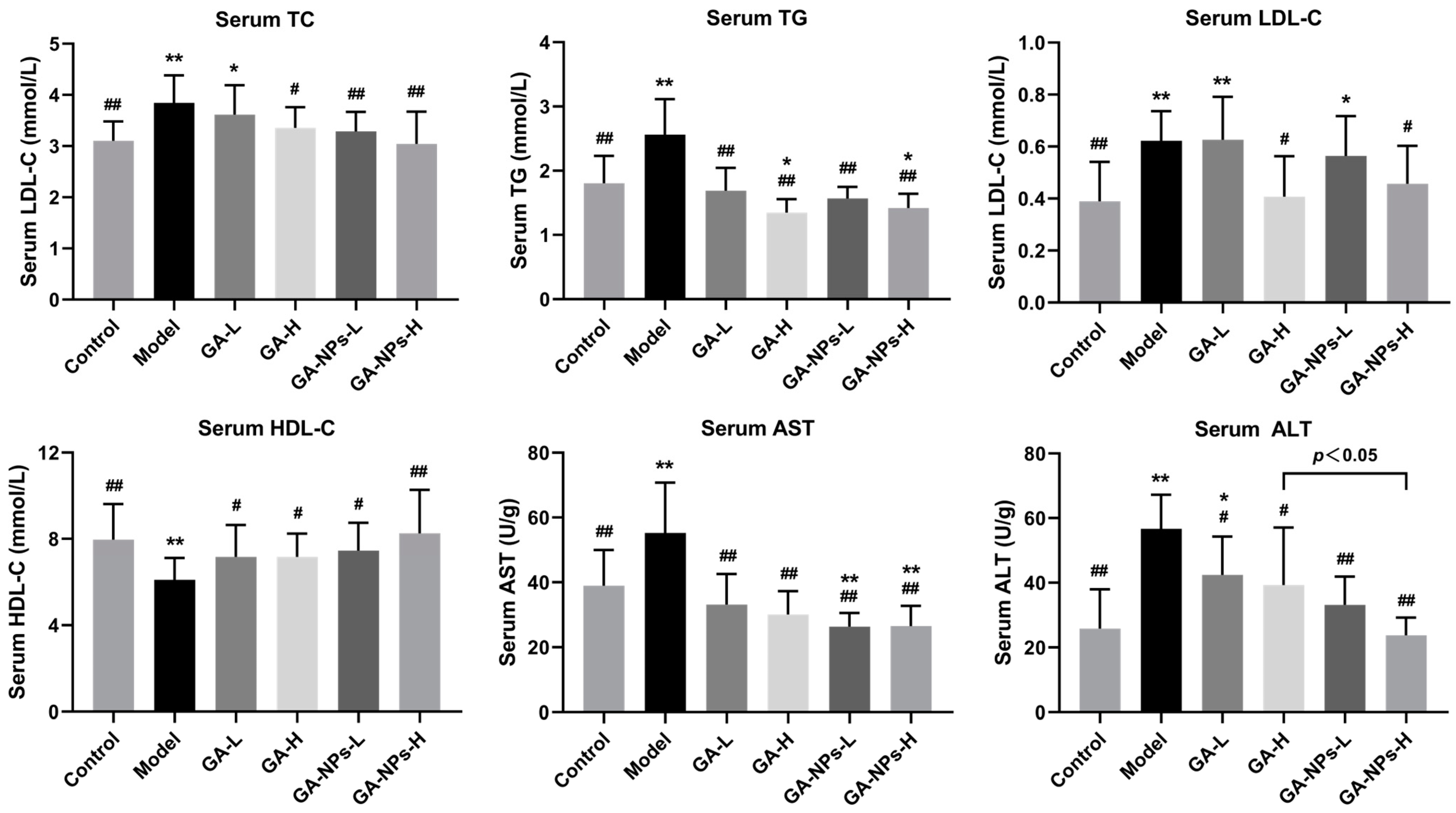
3.5. Effects of GA-NPs Intervention on Serum Biochemcial Parameters
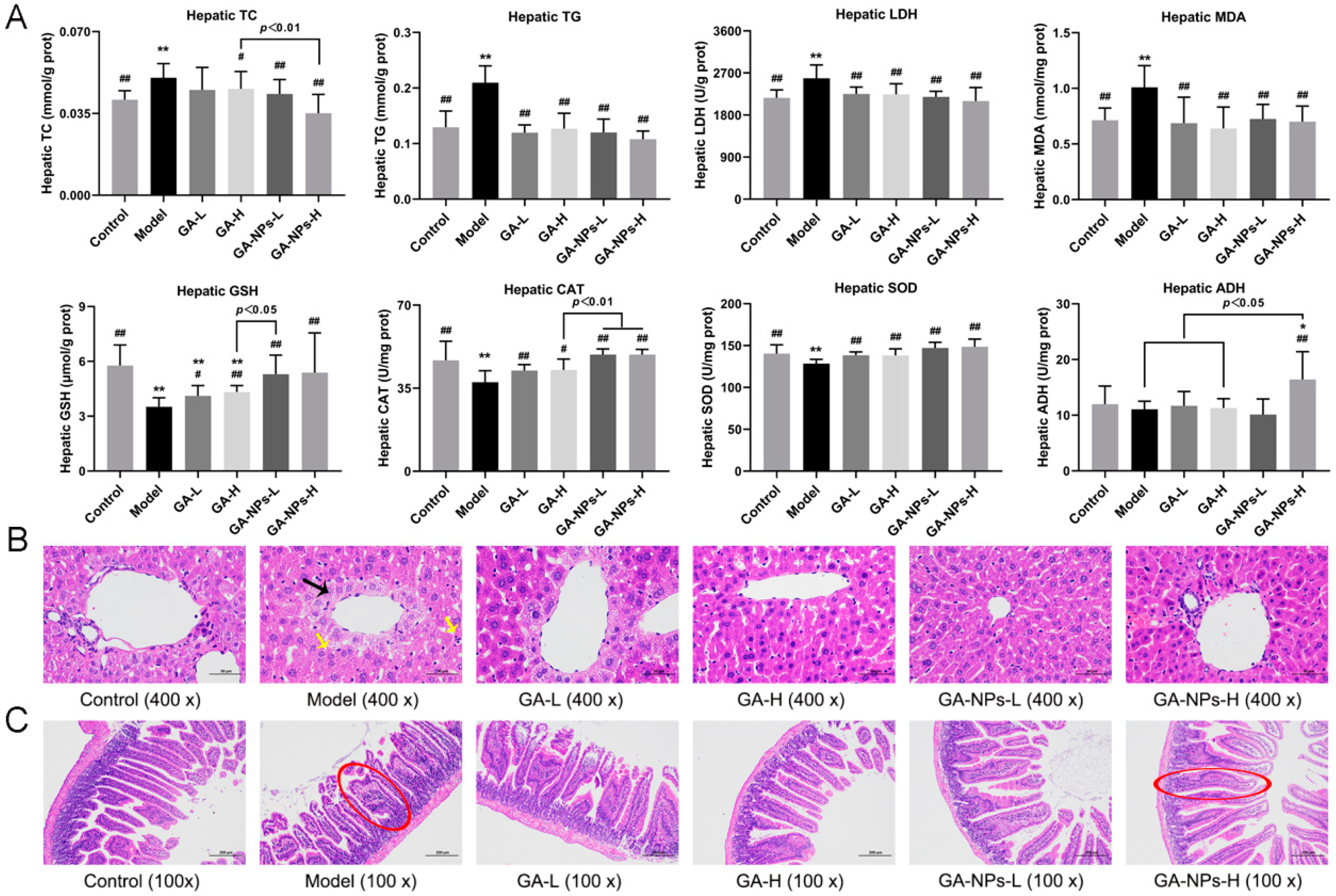
3.6. Effects of GA-NPs Intervention on Liver Biochemical Parameters
3.7. Effects of GA-NPs Intervention on Liver and Jejunum Histopathological Features
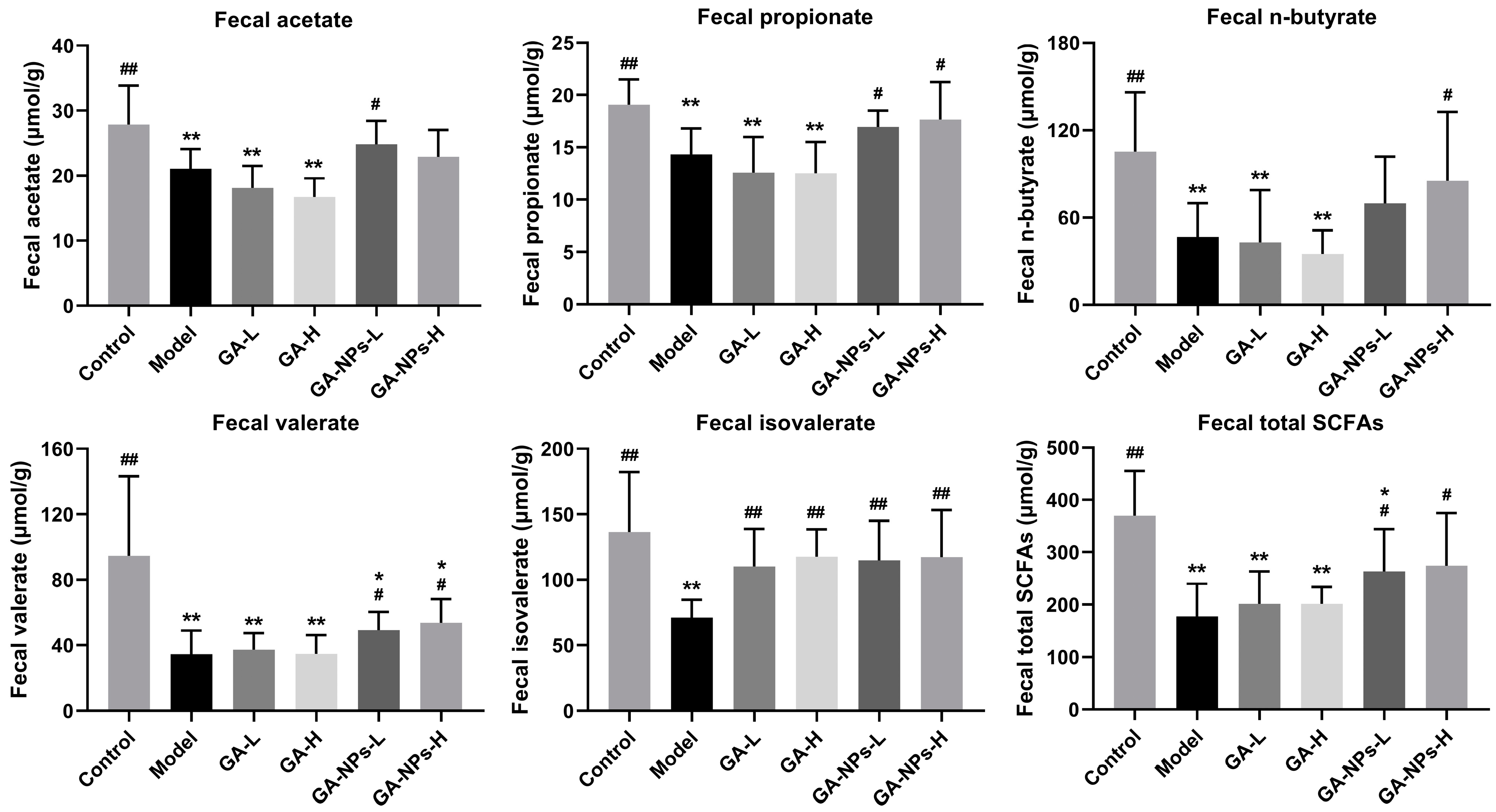
3.8. Effects of GA-NPs Intervention on Fecal SCFAs Levels
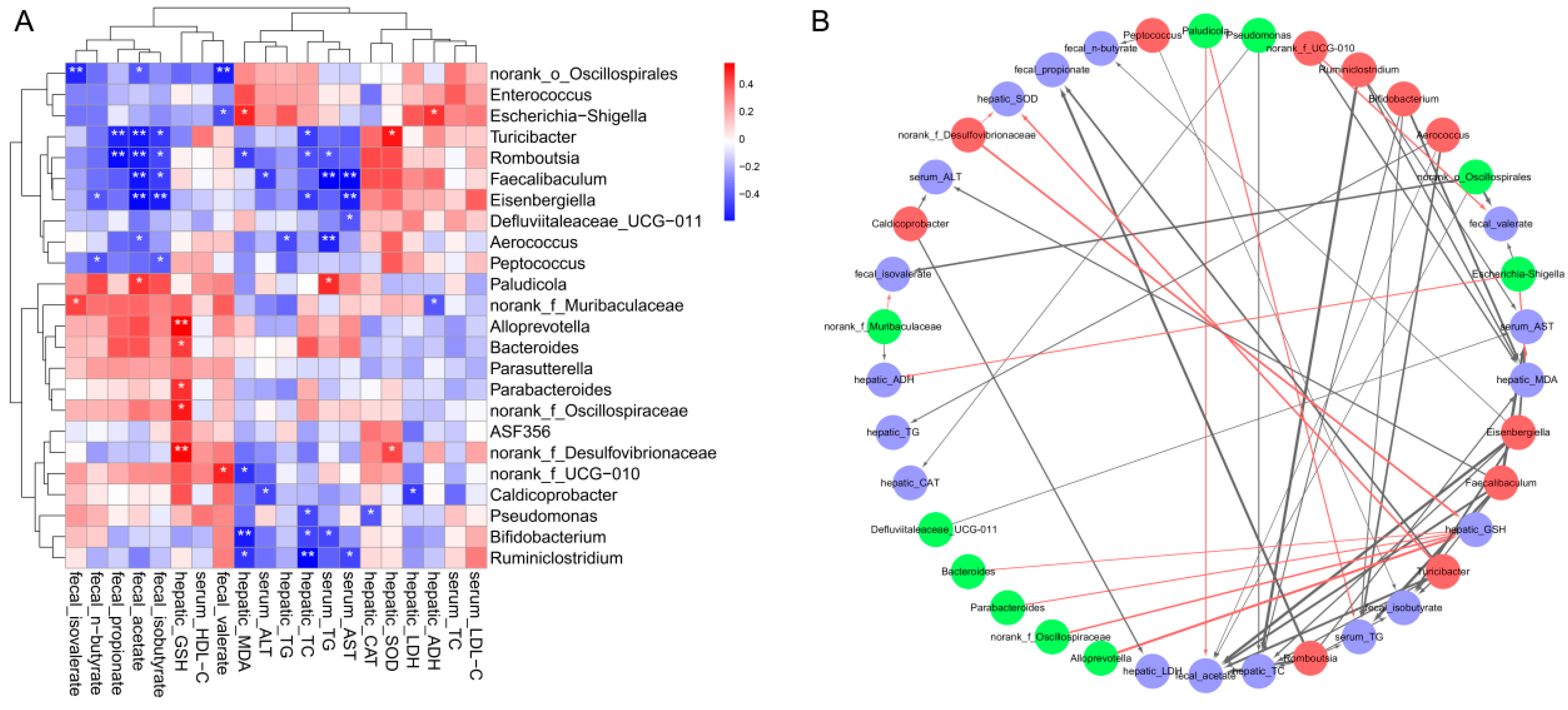
3.9. Effects of GA-NPs Intervention on the Composition of Intestinal Microflora
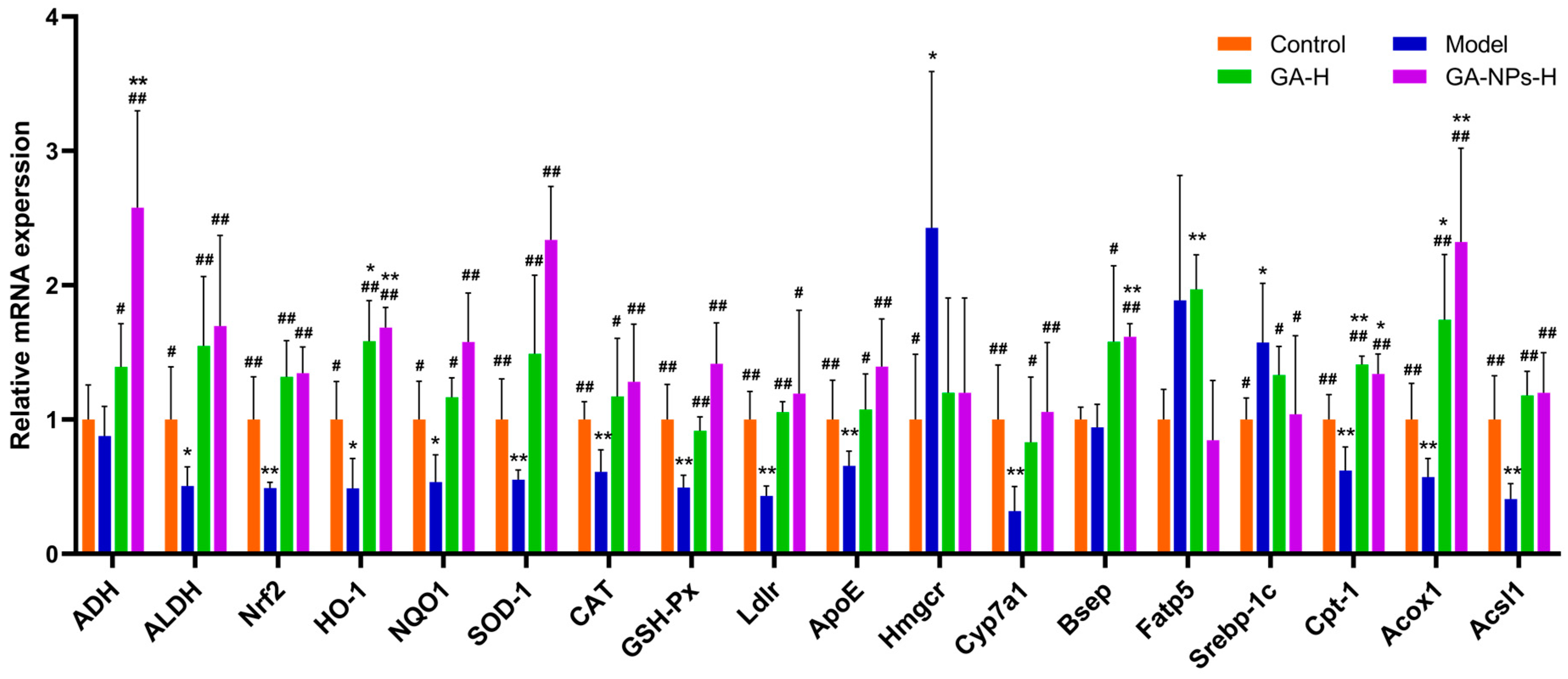
3.10. Effects of GA-NPs Intervention on the mRNA Transcription Levels in Liver
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, X.; Fan, X.; Miyata, T.; Kim, A.; Cajigas-Du, R.C.; Ray, S.; Huang, E.; Taiwo, M.; Arya, R.; Wu, J.; et al. Recent Advances in Understanding of Pathogenesis of Alcohol-Associated Liver Disease. Annu. Rev. Pathol. 2023, 18, 411–438. [Google Scholar] [CrossRef]
- Morencos, F.C.; De Las Heras Castaño, G.; Ramos, L.M.; Arias, M.J.L.; Ledesma, F.; Romero, F.P. Small bowel bacterial overgrowth in patients with alcoholic cirrhosis. Dig. Dis. Sci. 1996, 41, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Purohit, V.; Bode, J.C.; Bode, C.; Brenner, D.A.; Choudhry, M.A.; Hamilton, F.; Kang, Y.J.; Keshavarzian, A.; Rao, R.; Sartor, R.B.; et al. Alcohol, intestinal bacterial growth, intestinal permeability to endotoxin, and medical consequences: Summary of a symposium. Alcohol 2008, 42, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Llorente, C.; Lang, S.; Brandl, K.; Chu, H.; Jiang, L.; White, R.C.; Clarke, T.H.; Nguyen, K.; Torralba, M.; et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature 2019, 575, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Mackowiak, B.; Fu, Y.; Maccioni, L.; Gao, B. Alcohol-associated liver disease. J. Clin. Investig. 2024, 134, e176345. [Google Scholar] [CrossRef]
- Kong, L.Z.; Chandimali, N.; Han, Y.H.; Lee, D.H.; Kim, J.S.; Kim, S.U.; Kim, T.D.; Jeong, D.K.; Sun, H.N.; Lee, D.S.; et al. Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease. Int. J. Mol. Sci. 2019, 20, 2712. [Google Scholar] [CrossRef]
- Liang, J.; Olsen, R.W. Alcohol use disorders and current pharmacological therapies: The role of GABAA receptors. Acta Pharmacol. Sin. 2014, 35, 981–993. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, S.; Peng, B.; Tan, D.; Wu, M.; Wei, J.; Wang, Y.; Luo, H. Ganoderma lucidum: A comprehensive review of phytochemistry, efficacy, safety and clinical study. Food Sci. Hum. Wellness 2024, 13, 568–596. [Google Scholar] [CrossRef]
- Lu, J.; He, R.; Sun, P.; Zhang, F.; Linhardt, R.J.; Zhang, A. Molecular mechanisms of bioactive polysaccharides from Ganoderma lucidum (Lingzhi), a review. Int. J. Biol. Macromol. 2020, 150, 765–774. [Google Scholar] [CrossRef]
- Huang, C.-H.; Lin, W.-K.; Chang, S.; Tsai, G.-J. Evaluation of the hypoglycaemic and antioxidant effects of submerged Ganoderma lucidum cultures in type 2 diabetic rats. Mycology 2020, 12, 82–93. [Google Scholar] [CrossRef]
- Guo, W.-L.; Deng, J.-C.; Pan, Y.-Y.; Xu, J.-X.; Hong, J.-L.; Shi, F.-F.; Liu, G.-L.; Qian, M.; Bai, W.-D.; Zhang, W.; et al. Hypoglycemic and hypolipidemic activities of Grifola frondosa polysaccharides and their relationships with the modulation of intestinal microflora in diabetic mice induced by high-fat diet and streptozotocin. Int. J. Biol. Macromol. 2020, 153, 1231–1240. [Google Scholar] [CrossRef]
- Guo, W.-L.; Cao, Y.-J.; You, S.-Z.; Wu, Q.; Zhang, F.; Han, J.-Z.; Lv, X.-C.; Rao, P.-F.; Ai, L.-Z.; Ni, L. Ganoderic acids-rich ethanol extract from Ganoderma lucidum protects against alcoholic liver injury and modulates intestinal microbiota in mice with excessive alcohol intake. Curr. Res. Food Sci. 2022, 5, 515–530. [Google Scholar] [CrossRef]
- Cao, F.-R.; Xiao, B.-X.; Wang, L.-S.; Tao, X.; Yan, M.-Z.; Pan, R.-L.; Liao, Y.-H.; Liu, X.-M.; Chang, Q. Plasma and brain pharmacokinetics of ganoderic acid A in rats determined by a developed UFLC–MS/MS method. J. Chromatogr. B 2017, 1052, 19–26. [Google Scholar] [CrossRef]
- Li, K.; Zhang, Y.; Hao, X.; Xie, D.; Wang, C.; Zhang, H.; Jin, P.; Du, Q. Improved Stability and In Vitro Anti-Arthritis Bioactivity of Curcumin-Casein Nanoparticles by Ultrasound-Driven Encapsulation. Nutrients 2022, 14, 5192. [Google Scholar] [CrossRef]
- Witika, B.A.; Makoni, P.A.; Matafwali, S.K.; Chabalenge, B.; Mwila, C.; Kalungia, A.C.; Nkanga, C.I.; Bapolisi, A.M.; Walker, R.B. Biocompatibility of biomaterials for nanoencapsulation: Current approaches. Nanomaterials 2020, 10, 1649. [Google Scholar] [CrossRef]
- Liang, H.; Zhou, B.; He, L.; An, Y.; Lin, L.; Li, Y.; Liu, S.; Chen, Y.; Li, B. Fabrication of zein/quaternized chitosan nanoparticles for the encapsulation and protection of curcumin. RSC Adv. 2015, 5, 13891–13900. [Google Scholar] [CrossRef]
- Wei, Z.; Huang, Q. Assembly of Protein–Polysaccharide complexes for delivery of bioactive Ingredients: A Perspective paper. J. Agric. Food Chem. 2019, 67, 1344–1352. [Google Scholar] [CrossRef]
- Yu, S.; Xu, X.; Feng, J.; Liu, M.; Hu, K. Chitosan and chitosan coating nanoparticles for the treatment of brain disease. Int. J. Pharm. 2019, 560, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Shariatinia, Z. Pharmaceutical applications of chitosan. Adv. Colloid Interface Sci. 2019, 263, 131–194. [Google Scholar] [CrossRef]
- Li, M.-F.; Chen, L.; Xu, M.-Z.; Zhang, J.-L.; Wang, Q.; Zeng, Q.-Z.; Wei, X.-C.; Yuan, Y. The formation of zein-chitosan complex coacervated particles: Relationship to encapsulation and controlled release properties. Int. J. Biol. Macromol. 2018, 116, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Yan, H.; Wang, X.; Zhou, Y.; Gao, X.; Puligundla, P.; Wan, X. Encapsulation of epigallocatechin gallate in zein/chitosan nanoparticles for controlled applications in food systems. Food Chem. 2017, 231, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, L.; Huang, S.; Zhang, Y.; Xu, J.; Zhang, Z. Anti-cancer Effects of a Neutral Triterpene Fraction from Ganoderma lucidum and its Active Constituents on SW620 Human Colorectal Cancer Cells. Anti-Cancer Agents Med. Chem. 2020, 20, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Hernández, J.A.; Del-Toro-Sánchez, C.L.; Cinco-Moroyoqui, F.J.; Ruiz-Cruz, S.; Juárez, J.; Castro-Enríquez, D.D.; Barreras-Urbina, C.G.; López-Ahumada, G.A.; Rodríguez-Félix, F. Gallic Acid-Loaded zein nanoparticles by electrospraying process. J. Food Sci. 2019, 84, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhang, B.; Whent, M.; Yu, L.; Wang, Q. Preparation and characterization of zein/chitosan complex for encapsulation of α-tocopherol, and its in vitro controlled release study. Colloids Surf. B Biointerfaces 2011, 85, 145–152. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Arteel, G.E. Effect of ethanol on lipid metabolism. J. Hepatol. 2019, 70, 237–248. [Google Scholar] [CrossRef]
- Tan, J.K.; Macia, L.; Mackay, C.R. Dietary fiber and SCFAs in the regulation of mucosal immunity. J. Allergy Clin. Immunol. 2023, 151, 361–370. [Google Scholar] [CrossRef]
- Fairfield, B.; Schnabl, B. Gut dysbiosis as a driver in alcohol-induced liver injury. JHEP Rep. 2021, 3, 100220. [Google Scholar] [CrossRef]
- Gao, X.; Chang, S.; Liu, S.; Peng, L.; Xie, J.; Dong, W.; Tian, Y.; Sheng, J. Correlations between α-Linolenic Acid-Improved Multitissue Homeostasis and Gut Microbiota in Mice Fed a High-Fat Diet. MSystems 2020, 5, 10–128. [Google Scholar] [CrossRef]
- Inagaki, A.; Sakata, T. Dose-Dependent stimulatory and inhibitory effects of luminal and serosal n-Butyric acid on epithelial cell proliferation of pig distal colonic mucosa. J. Nutr. Sci. Vitaminol. 2005, 51, 156–160. [Google Scholar] [CrossRef]
- Raje, V.; Ahern, K.W.; Martinez, B.A.; Howell, N.L.; Oenarto, V.; Granade, M.E.; Kim, J.W.; Tundup, S.; Bottermann, K.; Gödecke, A.; et al. Adipocyte lipolysis drives acute stress-induced insulin resistance. Sci. Rep. 2020, 10, 18166. [Google Scholar] [CrossRef]
- Liu, J.; Yue, S.; Yang, Z.; Feng, W.; Meng, X.; Wang, A.; Peng, C.; Wang, C.; Yan, D. Oral hydroxysafflor yellow A reduces obesity in mice by modulating the gut microbiota and serum metabolism. Pharmacol. Res. 2018, 134, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, K.; Wu, W.; Lv, L.; Bian, X.; Yang, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; et al. Administration of Bifidobacterium bifidum CGMCC 15068 modulates gut microbiota and metabolome in azoxymethane (AOM)/dextran sulphate sodium (DSS)-induced colitis-associated colon cancer (CAC) in mice. Appl. Microbiol. Biotechnol. 2020, 104, 5915–5928. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, Y.; Wan, P.; Dong, W.; Huang, K.; Ran, L.; Mi, J.; Lu, L.; Zeng, X.; Cao, Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 108952. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-W.; Li, Y.-T.; Ye, J.-Z.; Lv, L.-X.; Yang, L.-Y.; Bian, X.-Y.; Wu, W.-R.; Wu, J.-J.; Shi, D.; Wang, Q.; et al. New strain of Pediococcus pentosaceus alleviates ethanol-induced liver injury by modulating the gut microbiota and short-chain fatty acid metabolism. World J. Gastroenterol. 2020, 26, 6224–6240. [Google Scholar] [CrossRef]
- Da Silva Duarte, V.; Cruz, B.C.D.S.; Tarrah, A.; Dias, R.S.; De Paula Dias Moreira, L.; Lemos, W.J.F., Jr.; Silva, L.C.F.; Santana, G.R.; De Oliveira, L.L.; Peluzio, M.D.C.G.; et al. Chemoprevention of DMH-Induced Early Colon Carcinogenesis in Male BALB/c Mice by Administration of Lactobacillus Paracasei DTA81. Microorganisms 2020, 8, 1994. [Google Scholar] [CrossRef]
- Xue, G.; Lai, S.; Li, X.; Zhang, W.; You, J.; Chen, H.; Qian, Y.; Gao, P.; Liu, Z.; Liu, Y. Efficient bioconversion of organic wastes to high optical activity of l-lactic acid stimulated by cathode in mixed microbial consortium. Water Res. 2018, 131, 1–10. [Google Scholar] [CrossRef]
- Jung, S.-J.; Hwang, J.-H.; Park, E.-O.; Lee, S.-O.; Chung, Y.-J.; Chung, M.-J.; Lim, S.; Lim, T.-J.; Ha, Y.; Park, B.-H.; et al. Regulation of alcohol and acetaldehyde metabolism by a mixture of lactobacillus and bifidobacterium species in human. Nutrients 2021, 13, 1875. [Google Scholar] [CrossRef]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.-X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 2020, 70, 761–774. [Google Scholar] [CrossRef]
- Massey, V.; Beier, J.; Ritzenthaler, J.; Roman, J.; Arteel, G. Potential role of the Gut/Liver/Lung axis in Alcohol-Induced tissue pathology. Biomolecules 2015, 5, 2477–2503. [Google Scholar] [CrossRef]
- Cederbaum, A.I. Alcohol metabolism. Clin. Liver Dis. 2012, 16, 667–685. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, T.; Song, Z. Chronic alcohol consumption disrupted cholesterol homeostasis in rats: Down-Regulation of Low-Density lipoprotein receptor and enhancement of cholesterol biosynthesis pathway in the liver. Alcohol/Alcohol Clin. Exp. Res. 2010, 34, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Sun, W.; Gao, L.; Liu, S. Therapeutic targets of hypercholesterolemia: HMGCR and LDLR. Diabetes Metab. Syndr. Obes. 2019, 12, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, K.L.; Ruan, X.Z.; Liu, B.C. Dysregulation of the Low-Density lipoprotein receptor pathway is involved in Lipid Disorder-Mediated organ injury. Int. J. Biol. Sci. 2016, 12, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S. Effects of ethanol upon lipid metabolism. Lipids 1974, 9, 103–116. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef]
- Kumari, A.; Pathak, D.P.; Asthana, S. Bile acids mediated potential functional interaction between FXR and FATP5 in the regulation of Lipid Metabolism. Int. J. Biol. Sci. 2020, 16, 2308–2322. [Google Scholar] [CrossRef]
- Quan, J.; Bode, A.M.; Luo, X. ACSL family: The regulatory mechanisms and therapeutic implications in cancer. Eur. J. Pharmacol. 2021, 909, 174397. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′−3′) | Reverse Primer (5′−3′) |
|---|---|---|
| ADH2 | AACGGTGAAKGTTCCCAAAA | ACGACCCCCAGCCTAATACA |
| ALDH2 | ATCCTCGGCTACATCAAATCG | GTCTTTTACGTCCCCGAACAC |
| Nrf2 | CCGGGAKCAAKCAGAKA | ACGTTGTCCCCATTTTTGCG |
| HO-1 | AKCCCAGTCTATGCCCCAC | GGCGTGCAAGGGATGATTTC |
| NQO1 | AACAAGCAAKCCCAGTCTATGC | AGGTAGCGGGTATATGCGTGGGCC |
| SOD-1 | TTGGCCGTACAA GGTGG | CGCAATCCCAATCACTCCAC |
| CAT | TCACCCACGATATCACCAGA | AGCTGAGCCTGACTCTCCAG |
| GSH-Px | GGGACCCTGAGACTTAGAGC | AATCCGTACTAGCGCTCACA |
| Ldlr | ATGCTGGAGATAGAGTGGAGTT | CCGCCAAGATCAAAKAG |
| ApoE | AKCCGCTTCTGGGATTACCT | TCAGTGCCGTCAGTTCTTGTG |
| Cyp7a1 | CCTTGGGACGTTTTCCTGCT | GCGCTCTTTGATTTAGAKG |
| Bsep | TCTGACTCAGTGATTCTTCGCA | CCCATAAACATCAGCCAGTTGT |
| Fatp5 | AKTCGGGAGGCAGAAKCT | AGCGGGTCATACAAGTGAGC |
| Srebp-1c | GCCGGCGCCATGGACGAGCTGG | CAGAKGGCTTCCAGAGAGGAG |
| Cpt-1 | TCCATGCATACCAAAGTGGA | TGGTAGGAGAGCAGCACCTT |
| Acox | GCCTGCTGTGTGGGTATGTCATT | GTCATGGGCGGGTGCAT |
| Acsl1 | CACTTCTTGCCTCGTTCCAC | GTCGTCCCGCTCTATGACAC |
| Mouse 18S | AGTCCCTGCCCTTTGTACACA | CGATCCCAGGGCCTCACTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Yang, Y.; Liang, Z.; Guo, W.; Lv, X.; Ni, L.; Chen, Y. Synthesis of Ganoderic Acids Loaded Zein-Chitosan Nanoparticles and Evaluation of Their Hepatoprotective Effect on Mice Given Excessive Alcohol. Foods 2024, 13, 2760. https://doi.org/10.3390/foods13172760
Cao Y, Yang Y, Liang Z, Guo W, Lv X, Ni L, Chen Y. Synthesis of Ganoderic Acids Loaded Zein-Chitosan Nanoparticles and Evaluation of Their Hepatoprotective Effect on Mice Given Excessive Alcohol. Foods. 2024; 13(17):2760. https://doi.org/10.3390/foods13172760
Chicago/Turabian StyleCao, Yingjia, Yuheng Yang, Zihua Liang, Weiling Guo, Xucong Lv, Li Ni, and Youting Chen. 2024. "Synthesis of Ganoderic Acids Loaded Zein-Chitosan Nanoparticles and Evaluation of Their Hepatoprotective Effect on Mice Given Excessive Alcohol" Foods 13, no. 17: 2760. https://doi.org/10.3390/foods13172760
APA StyleCao, Y., Yang, Y., Liang, Z., Guo, W., Lv, X., Ni, L., & Chen, Y. (2024). Synthesis of Ganoderic Acids Loaded Zein-Chitosan Nanoparticles and Evaluation of Their Hepatoprotective Effect on Mice Given Excessive Alcohol. Foods, 13(17), 2760. https://doi.org/10.3390/foods13172760