

Technological and Enzymatic Characterization of Autochthonous Lactic Acid Bacteria Isolated from Viili Natural Starters



Abstract
1. Introduction
2. Materials and Methods
2.1. Activation of Viili Starters and Monitoring of Fermentation
2.2. Isolation and Characterization of Lactic Acid Bacteria
2.3. Enzymatic Activities
2.4. Acidification Performance
2.5. In Vitro EPS Production
2.6. Detection of the hdcA Gene of Gram-Positive Bacteria
2.7. Assessment of Antimicrobial Activity
2.8. Statistical Analysis
3. Results and Discussion
3.1. Physico-Chemical and Microbiological Parameters
3.2. Identification of the Lactic Acid Bacteria Isolates
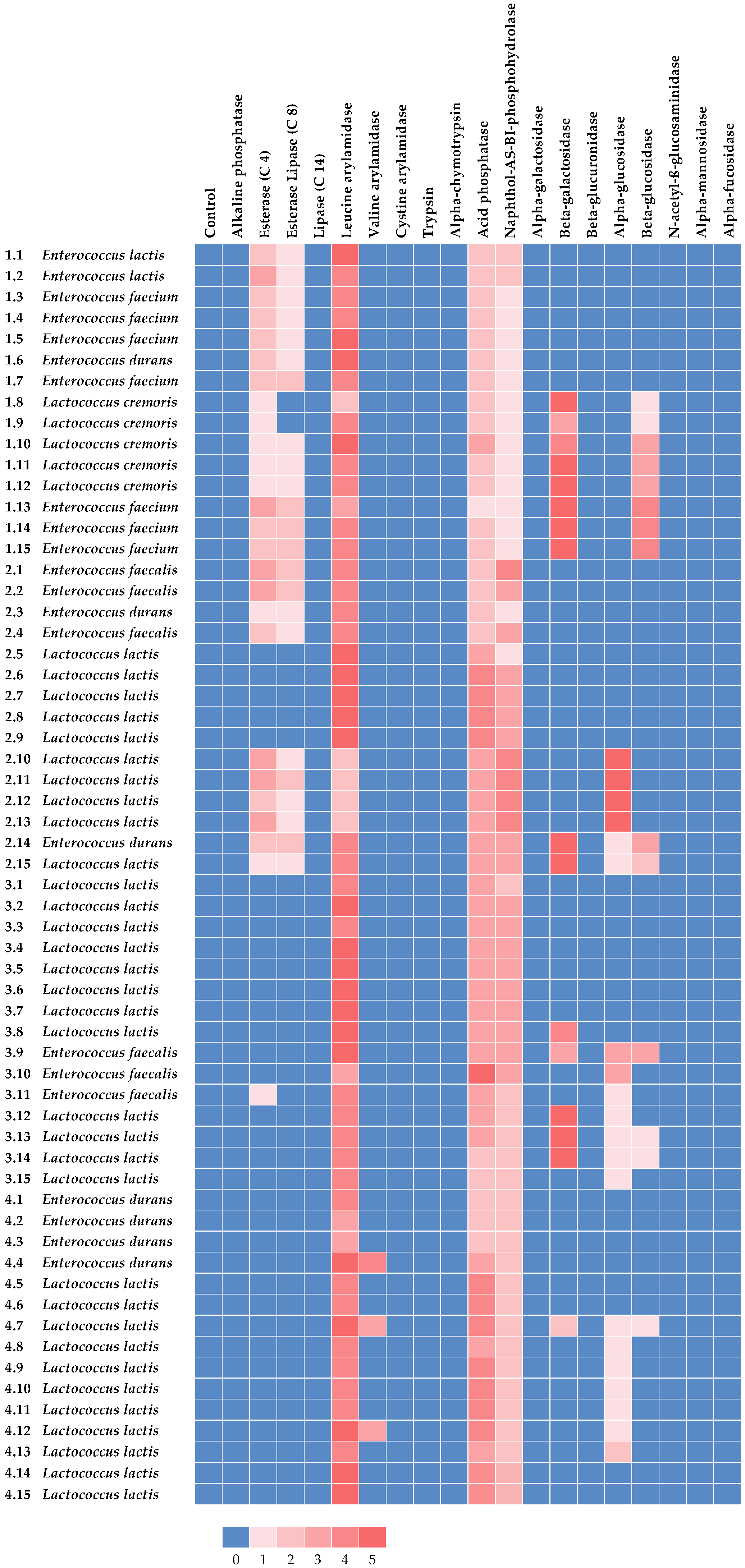
3.3. Enzymatic Activities of the Lactic Acid Bacteria Isolates
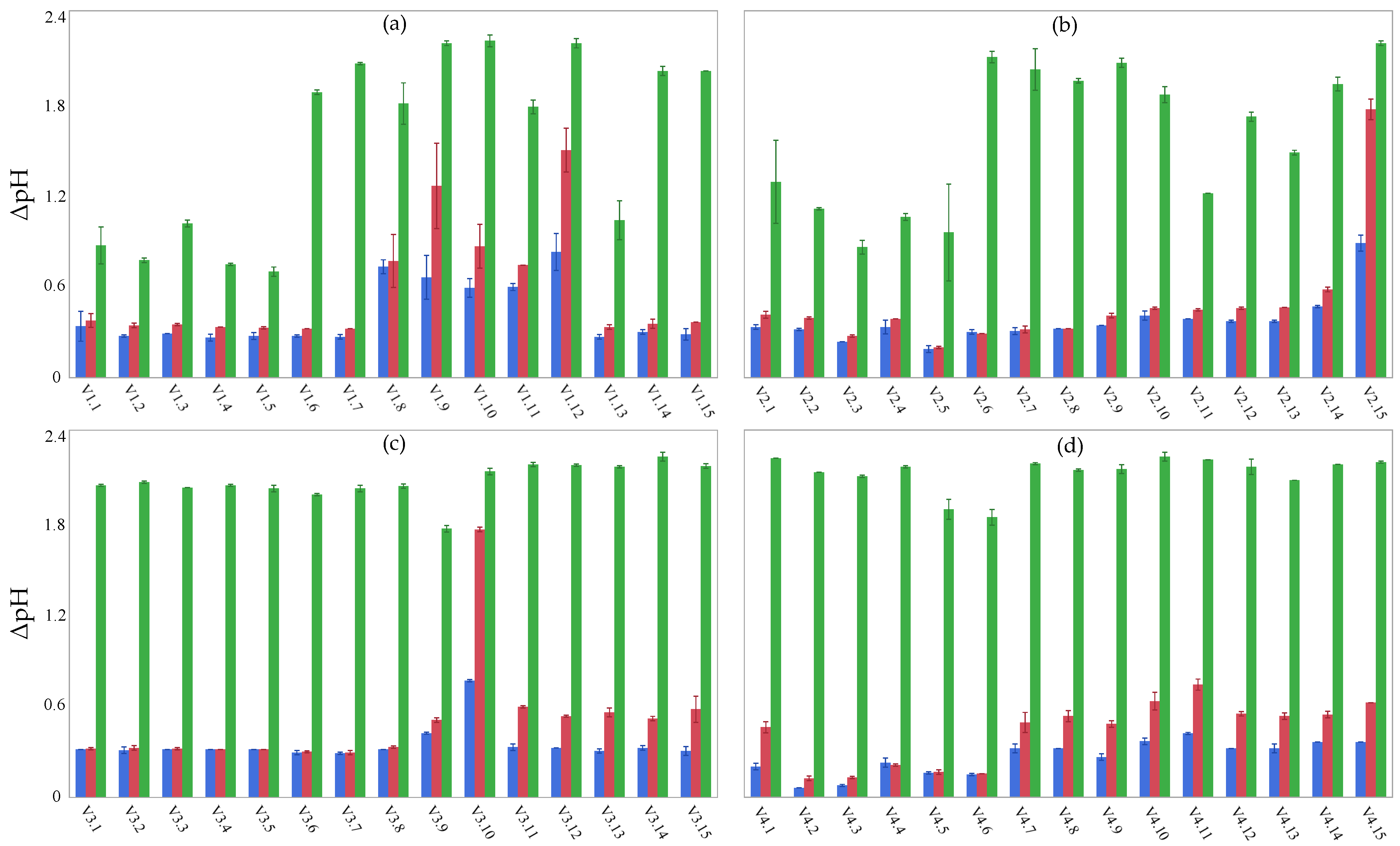
3.4. Acidification Performance
3.5. In Vitro EPS Production
3.6. Presence of hdcA Gene and Assessment of Antimicrobial Activity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tamime, A.Y. Fermented Milks: A Historical Food with Modern Applications—A Review. Eur. J. Clin. Nutr. 2002, 56, S2–S15. [Google Scholar] [CrossRef] [PubMed]
- Evershed, R.P.; Payne, S.; Sherratt, A.G.; Copley, M.S.; Coolidge, J.; Urem-Kotsu, D.; Kotsakis, K.; Özdoǧan, M.; Özdoǧan, A.E.; Nieuwenhuyse, O.; et al. Earliest Date for Milk Use in the Near East and Southeastern Europe Linked to Cattle Herding. Nature 2008, 455, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T.; Papademas, P. The Evolution of Fermented Milks, from Artisanal to Industrial Products: A Critical Review. Fermentation 2022, 8, 679. [Google Scholar] [CrossRef]
- Codex Stan 243-2003; Codex Standard for Fermented Milks. Codex Alimentarius: Rome, Italy, 2003.
- Macori, G.; Cotter, P.D. Novel Insights into the Microbiology of Fermented Dairy Foods. Curr. Opin. Biotechnol. 2018, 49, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.K.; Tamime, A.Y.; Wszolek, M. Microbiology of Fermented Milks. In Dairy Microbiology Handbook; Robinson, R.K., Ed.; John Wiley and Sons, Inc.: New York, NY, USA, 2002; pp. 367–421. [Google Scholar]
- Maoloni, A.; Blaiotta, G.; Ferrocino, I.; Mangia, N.P.; Osimani, A.; Milanović, V.; Cardinali, F.; Cesaro, C.; Garofalo, C.; Clementi, F.; et al. Microbiological Characterization of Gioddu, an Italian Fermented Milk. Int. J. Food Microbiol. 2020, 323, 108610. [Google Scholar] [CrossRef] [PubMed]
- Leroy, F.; De Vuyst, L. Lactic Acid Bacteria as Functional Starter Cultures for the Food Fermentation Industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Narvhus, J.A.; Abrahamsen, R.K. Traditional and Modern Nordic Fermented Milk Products: A Review. Int. Dairy J. 2023, 142, 105641. [Google Scholar] [CrossRef]
- Abi Khalil, R.; Yvon, S.; Couderc, C.; Jard, G.; El Rammouz, R.; Abi Nakhoul, P.; Eutamène, H.; Ayoub, M.J.; Tormo, H. Traditional Fermented Milk Products of Eastern Mediterranean Countries: A Cultural Heritage to Preserve. Int. Dairy J. 2023, 147, 105768. [Google Scholar] [CrossRef]
- Mallappa, R.H.; Balasubramaniam, C.; Nataraj, B.H.; Ramesh, C.; Kadyan, S.; Pradhan, D.; Muniyappa, S.K.; Grover, S. Microbial Diversity and Functionality of Traditional Fermented Milk Products of India: Current Scenario and Future Perspectives. Int. Dairy J. 2021, 114, 104941. [Google Scholar] [CrossRef]
- Moonga, H.B.; Schoustra, S.E.; Linnemann, A.R.; Shindano, J.; Smid, E.J. Towards Valorisation of Indigenous Traditional Fermented Milk: Mabisi as a Model. Curr. Opin. Food Sci. 2022, 46, 100835. [Google Scholar] [CrossRef]
- Ferrocino, I.; Rantsiou, K.; Cocolin, L. Investigating Dairy Microbiome: An Opportunity to Ensure Quality, Safety and Typicity. Curr. Opin. Biotechnol. 2022, 73, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Goa, T.; Beyene, G.; Mekonnen, M.; Gorems, K. Isolation and Characterization of Lactic Acid Bacteria from Fermented Milk Produced in Jimma Town, Southwest Ethiopia, and Evaluation of Their Antimicrobial Activity against Selected Pathogenic Bacteria. Int. J. Food Sci. 2022, 2022, 2076021. [Google Scholar] [CrossRef] [PubMed]
- Chammas, G.I.; Saliba, R.; Corrieu, G.; Béal, C. Characterisation of Lactic Acid Bacteria Isolated from Fermented Milk “Laban”. Int. J. Food Microbiol. 2006, 110, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Nicosia, F.D.; Pino, A.; Maciel, G.L.R.; Sanfilippo, R.R.; Caggia, C.; de Carvalho, A.F.; Randazzo, C.L. Technological Characterization of Lactic Acid Bacteria Strains for Potential Use in Cheese Manufacture. Foods 2023, 12, 1154. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y.; Dong, L.; Li, Y.; Liu, Y.; Liu, Y.; Liu, L.; Liu, L. Fermentation of Lactobacillus Fermentum NB02 with Feruloyl Esterase Production Increases the Phenolic Compounds Content and Antioxidant Properties of Oat Bran. Food Chem. 2024, 437, 137834. [Google Scholar] [CrossRef] [PubMed]
- Narvhus, J.A.; Abrahamsen, R.K. Nordic Fermented Milks. Encycl. Dairy Sci. 2021, 464–471. [Google Scholar] [CrossRef]
- Porcellato, D.; Tranvåg, M.; Narvhus, J. Short Communication: Presence of Lactococcus and Lactococcal Exopolysaccharide Operons on the Leaves of Pinguicula Vulgaris Supports the Traditional Source of Bacteria Present in Scandinavian Ropy Fermented Milk. J. Dairy Sci. 2016, 99, 7049–7052. [Google Scholar] [CrossRef]
- Chen, T.; Tan, Q.; Wang, M.; Xiong, S.; Jiang, S.; Wu, Q.; Li, S.; Luo, C.; Wei, H. Identification of Bacterial Strains in Viili by Molecular Taxonomy and Their Synergistic Effects on Milk Curd and Exopolysaccharides Production. Afr. J. Biotechnol. 2011, 10, 16969–16975. [Google Scholar] [CrossRef]
- Kahala, M.; Mäki, M.; Lehtovaara, A.; Tapanainen, J.M.; Katiska, R.; Juuruskorpi, M.; Juhola, J.; Joutsjoki, V. Characterization of Starter Lactic Acid Bacteria from the Finnish Fermented Milk Product Viili. J. Appl. Microbiol. 2008, 105, 1929–1938. [Google Scholar] [CrossRef] [PubMed]
- Ruas-Madiedo, P.; Gueimonde, M.; De Los Reyes-Gavilán, C.G.; Salminen, S. Short Communication: Effect of Exopolysaccharide Isolated from “Viili” on the Adhesion of Probiotics and Pathogens to Intestinal Mucus. J. Dairy Sci. 2006, 89, 2355–2358. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Handa, S.; Imai, M.; Harada, N.; Sakamoto, T.; Ishida, T.; Nakagaki, T.; Nakano, Y. Exopolysaccharides from a Scandinavian Fermented Milk Viili Increase Butyric Acid and Muribaculum Members in the Mouse Gut. Food Chem. Mol. Sci. 2021, 3, 100042. [Google Scholar] [CrossRef] [PubMed]
- ANSES (Agence Nationale de Sécurité Sanitaire de L’alimentation, de L’environnement et du Travail). EURL Lm TECHNICAL GUIDANCE DOCUMENT for Conducting Shelflife Studies on Listeria Monocytogenes in Ready-to-Eat Foods. Version 3 of 6 June 2014—Amendment 1 of 21 February 2019. Available online: https://eurl-listeria.anses.fr/en/system/files/LIS-Cr-201909D2.pdf (accessed on 14 November 2023).
- Rampanti, G.; Raffo, A.; Melini, V.; Moneta, E.; Nardo, N.; Saggia Civitelli, E.; Bande-De León, C.; Tejada Portero, L.; Ferrocino, I.; Franciosa, I.; et al. Chemical, microbiological, textural, and sensory characteristics of pilot-scale Caciofiore cheese curdled with commercial Cynara Cardunculus Rennet and crude extracts from spontaneous and cultivated Onopordum Tauricum. Food Res. Int. 2023, 173, 113459. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Garofalo, C.; Aquilanti, L.; Milanović, V.; Clementi, F. Unpasteurised Commercial Boza as a Source of Microbial Diversity. Int. J. Food Microbiol. 2015, 194, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Linares-Morales, J.R.; Cuellar-Nevárez, G.E.; Rivera-Chavira, B.E.; Gutiérrez-Méndez, N.; Pérez-Vega, S.B.; Nevárez-Moorillón, G.V. Selection of Lactic Acid Bacteria Isolated from Fresh Fruits and Vegetables Based on Their Antimicrobial and Enzymatic Activities. Foods 2020, 9, 1399. [Google Scholar] [CrossRef] [PubMed]
- Hilbig, J.; Gisder, J.; Prechtl, R.M.; Herrmann, K.; Weiss, J.; Loeffler, M. Influence of Exopolysaccharide-Producing Lactic Acid Bacteria on the Spreadability of Fat-Reduced Raw Fermented Sausages (Teewurst). Food Hydrocoll. 2019, 93, 422–431. [Google Scholar] [CrossRef]
- Belleggia, L.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Aquilanti, L.; Osimani, A. Prevalence of Histidine Decarboxylase Genes of Gram-Positive Bacteria in Surströmming as Revealed by QPCR. Indian J. Microbiol. 2021, 61, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.; Del Río, B.; Linares, D.M.; Martín, M.C.; Alvarez, M.A. Real-Time Polymerase Chain Reaction for Quantitative Detection of Histamine-Producing Bacteria: Use in Cheese Production. J. Dairy Sci. 2006, 89, 3763–3769. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, F.; Botta, C.; Harasym, J.; Reale, A.; Ferrocino, I.; Boscaino, F.; Orkusz, A.; Milanović, V.; Garofalo, C.; Rampanti, G.; et al. Tasting of Traditional Polish Fermented Cucumbers: Microbiology, Morpho-Textural Features, and Volatilome. Food Res. Int. 2024, 177, 113851. [Google Scholar] [CrossRef]
- Bassi, D.; Puglisi, E.; Cocconcelli, P.S. Comparing Natural and Selected Starter Cultures in Meat and Cheese Fermentations. Curr. Opin. Food Sci. 2015, 2, 118–122. [Google Scholar] [CrossRef]
- Cavanagh, D.; Fitzgerald, G.F.; McAuliffe, O. From Field to Fermentation: The Origins of Lactococcus Lactis and Its Domestication to the Dairy Environment. Food Microbiol. 2015, 47, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Rampanti, G.; Belleggia, L.; Cardinali, F.; Milanović, V.; Osimani, A.; Garofalo, C.; Ferrocino, I.; Aquilanti, L. Microbial Dynamics of a Specialty Italian Raw Ewe’s Milk Cheese Curdled with Extracts from Spontaneous and Cultivated Onopordum Tauricum Willd. Microorganisms 2023, 11, 219. [Google Scholar] [CrossRef] [PubMed]
- Shori, A.B. Comparative Study of Chemical Composition, Isolation and Identification of Micro-Flora in Traditional Fermented Camel Milk Products: Gariss, Suusac, and Shubat. J. Saudi Soc. Agric. Sci. 2012, 11, 79–88. [Google Scholar] [CrossRef]
- Arrizza, S.; Ledda, A.; Sarra, P.G.; Dellaglio, F. Identification of Lactic Acid Bacteria in Gioddu. Sci. Tech. Latt.-Casearia 1983, 34, 87–102. [Google Scholar]
- Chaves-López, C.; Serio, A.; Martuscelli, M.; Paparella, A.; Osorio-Cadavid, E.; Suzzi, G. Microbiological Characteristics of Kumis, a Traditional Fermented Colombian Milk, with Particular Emphasis on Enterococci Population. Food Microbiol. 2011, 28, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Dapkevicius, M.d.L.E.; Sgardioli, B.; Câmara, S.P.A.; Poeta, P.; Malcata, F.X. Current Trends of Enterococci in Dairy Products: A Comprehensive Review of Their Multiple Roles. Foods 2021, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Functionality of Enterococci in Dairy Products. Int. J. Food Microbiol. 2003, 88, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Bayjanov, J.R.; Renckens, B.; Nauta, A.; Siezen, R.J. The Proteolytic System of Lactic Acid Bacteria Revisited: A Genomic Comparison. BMC Genom. 2010, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic Systems of Lactic Acid Bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- García-Cano, I.; Rocha-Mendoza, D.; Ortega-Anaya, J.; Wang, K.; Kosmerl, E.; Jiménez-Flores, R. Lactic Acid Bacteria Isolated from Dairy Products as Potential Producers of Lipolytic, Proteolytic and Antibacterial Proteins. Appl. Microbiol. Biotechnol. 2019, 103, 5243–5257. [Google Scholar] [CrossRef] [PubMed]
- Mcsweeney, P.L.H.; Sousa, M.J. Biochemical Pathways for the Production of Flavour Compounds in Cheeses during Ripening: A Review. Le Lait 2000, 80, 293–324. [Google Scholar] [CrossRef]
- Kieliszek, M.; Pobiega, K.; Piwowarek, K.; Kot, A.M. Characteristics of the Proteolytic Enzymes Produced by Lactic Acid Bacteria. Molecules 2021, 26, 1858. [Google Scholar] [CrossRef] [PubMed]
- El-Ghaish, S.; Dalgalarrondo, M.; Choiset, Y.; Sitohy, M.; Ivanova, I.; Haertlé, T.; Chobert, J.M. Screening of Strains of Lactococci Isolated from Egyptian Dairy Products for Their Proteolytic Activity. Food Chem. 2010, 120, 758–764. [Google Scholar] [CrossRef]
- Saidi, Y.; del Rio, B.; Senouci, D.E.; Redruello, B.; Martinez, B.; Ladero, V.; Kihal, M.; Alvarez, M.A. Polyphasic Characterisation of Non-Starter Lactic Acid Bacteria from Algerian Raw Camel’s Milk and Their Technological Aptitudes. Food Technol. Biotechnol. 2020, 58, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Psomas, E.; Sakaridis, I.; Boukouvala, E.; Karatzia, M.A.; Ekateriniadou, L.V.; Samouris, G. Indigenous Lactic Acid Bacteria Isolated from Raw Graviera Cheese and Evaluation of Their Most Important Technological Properties. Foods 2023, 12, 370. [Google Scholar] [CrossRef] [PubMed]
- Treichel, H.; de Oliveira, D.; Mazutti, M.A.; Di Luccio, M.; Oliveira, J.V. A Review on Microbial Lipases Production. Food Bioproc. Tech. 2010, 3, 182–196. [Google Scholar] [CrossRef]
- Medina, R.B.; Katz, M.B.; González, S.; Oliver, G. Determination of Esterolytic and Lipolytic Activities of Lactic Acid Bacteria. In Public Health Microbiology: Methods and Protocols; Spencer, J.F., Ragout de Spencer, A.L., Eds.; Spencer Humana Press Inc.: Totowa, NJ, USA, 2004; Volume 268, pp. 465–470. [Google Scholar]
- Katz, M.; Medina, R.; Gonzalez, S.; Oliver, G. Esterolytic and Lipolytic Activities of Lactic Acid Bacteria Isolated from Ewe’s Milk and Cheese. J. Food Prot. 2002, 65, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Yerlikaya, O. Probiotic Potential and Biochemical and Technological Properties of Lactococcus Lactis Ssp. Lactis Strains Isolated from Raw Milk and Kefir Grains. J. Dairy Sci. 2019, 102, 124–134. [Google Scholar] [CrossRef] [PubMed]
- El-Din, B.B.; El-Soda, M.; Ezzat, N. Proteolytic, Lipolytic and Autolytic Activities of Enterococci Strains Isolated from Egyptian Dairy Products. Lait 2002, 82, 289–304. [Google Scholar] [CrossRef][Green Version]
- Morandi, S.; Brasca, M.; Andrighetto, C.; Lombardi, A.; Lodi, R. Technological and Molecular Characterisation of Enterococci Isolated from North-West Italian Dairy Products. Int. Dairy J. 2006, 16, 867–875. [Google Scholar] [CrossRef]
- Graham, K.; Stack, H.; Rea, R. Safety, Beneficial and Technological Properties of Enterococci for Use in Functional Food Applications–a Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 3836–3861. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic Diversity, Safety and Technological Characterization of Lactic Acid Bacteria Isolated from Artisanal Pico Cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Abarquero, D.; Bodelón, R.; Flórez, A.B.; Fresno, J.M.; Renes, E.; Mayo, B.; Tornadijo, M.E. Technological and Safety Assessment of Selected Lactic Acid Bacteria for Cheese Starter Cultures Design: Enzymatic and Antimicrobial Activity, Antibiotic Resistance and Biogenic Amine Production. LWT 2023, 180, 114709. [Google Scholar] [CrossRef]
- Rampanti, G.; Ferrocino, I.; Harasym, J.; Foligni, R.; Cardinali, F.; Orkusz, A.; Milanović, V.; Franciosa, I.; Garofalo, C.; Mannozzi, C.; et al. Queijo Serra Da Estrela PDO Cheese: Investigation into Its Morpho-Textural Traits, Microbiota, and Volatilome. Foods 2023, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Kamarinou, C.S.; Papadopoulou, O.S.; Doulgeraki, A.I.; Tassou, C.C.; Galanis, A.; Chorianopoulos, N.G.; Argyri, A.A. Mapping the Key Technological and Functional Characteristics of Indigenous Lactic Acid Bacteria Isolated from Greek Traditional Dairy Products. Microorganisms 2022, 10, 246. [Google Scholar] [CrossRef] [PubMed]
- Kirilov, N.; Dimov, S.; Dalgalarrondo, M.; Ignatova, T.; Kambarev, S.; Stoyanovski, S.; Danova, S.; Iliev, I.; Haertlé, T.; Chobert, J.M.; et al. Characterization of Enterococci Isolated from Homemade Bulgarian Cheeses and Katuk. Eur. Food Res. Technol. 2011, 233, 1029–1040. [Google Scholar] [CrossRef]
- Serio, A.; Chaves-López, C.; Paparella, A.; Suzzi, G. Evaluation of Metabolic Activities of Enterococci Isolated from Pecorino Abruzzese Cheese. Int. Dairy J. 2010, 20, 459–464. [Google Scholar] [CrossRef]
- Tsanasidou, C.; Asimakoula, S.; Sameli, N.; Fanitsios, C.; Vandera, E.; Bosnea, L.; Koukkou, A.I.; Samelis, J. Safety Evaluation, Biogenic Amine Formation, and Enzymatic Activity Profiles of Autochthonous Enterocin-Producing Greek Cheese Isolates of the Enterococcus Faecium/Durans Group. Microorganisms 2021, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Akuzawa, R.; Fox, P.F. Acid Phosphatase in Cheese. Anim. Sci. J. 2004, 75, 385–391. [Google Scholar] [CrossRef]
- Coimbra-Gomes, J.; Reis, P.J.M.; Tavares, T.G.; Faria, M.A.; Malcata, F.X.; Macedo, A.C. Evaluating the Probiotic Potential of Lactic Acid Bacteria Implicated in Natural Fermentation of Table Olives, Cv. Cobrançosa. Molecules 2023, 28, 3285. [Google Scholar] [CrossRef] [PubMed]
- Kolev, P.; Rocha-Mendoza, D.; Ruiz-Ramírez, S.; Ortega-Anaya, J.; Jiménez-Flores, R.; García-Cano, I. Screening and Characterization of β-Galactosidase Activity in Lactic Acid Bacteria for the Valorization of Acid Whey. JDS Commun. 2022, 3, 1–6. [Google Scholar] [CrossRef]
- Câmara, S.P.; Dapkevicius, A.; Riquelme, C.; Elias, R.B.; Silva, C.C.G.; Malcata, F.X.; Dapkevicius, M.L.N.E. Potential of Lactic Acid Bacteria from Pico Cheese for Starter Culture Development. Food Sci. Technol. Int. 2019, 25, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, H.; Kneifel, W. β-Glucosidase Activities of Lactic Acid Bacteria: Mechanisms, Impact on Fermented Food and Human Health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dewan, S.; Tamang, J.P. Dominant Lactic Acid Bacteria and Their Technological Properties Isolated from the Himalayan Ethnic Fermented Milk Products. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2007, 92, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Jawan, R.; Abbasiliasi, S.; Mustafa, S.; Kapri, M.R.; Halim, M.; Ariff, A.B. In Vitro Evaluation of Potential Probiotic Strain Lactococcus Lactis Gh1 and Its Bacteriocin-Like Inhibitory Substances for Potential Use in the Food Industry. Probiotics Antimicrob. Proteins 2021, 13, 422–440. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dong, L.; Li, Y.; Chen, Q.; Wang, L.; Farag, M.A.; Liu, L.; Zhan, S.; Wu, Z.; Liu, L. Soy Protein Isolate-Citrus Pectin Composite Hydrogels Induced by TGase and Ultrasonic Treatment: Potential Targeted Delivery System for Probiotics. Food Hydrocoll. 2023, 143, 108901. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Coelho, M.C.; Silva, C.C.G. A Rapid Screening Method to Evaluate Acidifying Activity by Lactic Acid Bacteria. J. Microbiol. Methods 2021, 185, 106227. [Google Scholar] [CrossRef] [PubMed]
- Sionek, B.; Szydłowska, A.; Küçükgöz, K.; Kołożyn-Krajewska, D. Traditional and New Microorganisms in Lactic Acid Fermentation of Food. Fermentation 2023, 9, 1019. [Google Scholar] [CrossRef]
- Li, W.; Ren, M.; Duo, L.; Li, J.; Wang, S.; Sun, Y.; Li, M.; Ren, W.; Hou, Q.; Yu, J.; et al. Fermentation Characteristics of Lactococcus Lactis Subsp. Lactis Isolated from Naturally Fermented Dairy Products and Screening of Potential Starter Isolates. Front. Microbiol. 2020, 11, 1794. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, Z. Acid and Rennet-Induced Coagulation Behavior of Casein Micelles with Modified Structure. Food Chem. 2019, 291, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Beresford, T.P.; Fitzsimons, N.A.; Brennan, N.L.; Cogan, T.M. Recent Advances in Cheese Microbiology. Int. Dairy J. 2001, 11, 259–274. [Google Scholar] [CrossRef]
- Cogan, T.M.; Barbosa, M.; Beuvier, E.; Bianchi-salvadori, B.; Cocconcelli, P.S.; Fernandes, I.; Gomezr, J.; Gomez, R.; Kalantzopoulos, G.; Ledda, A.; et al. Characterization of the Lactic Acid Bacteria in Artisanal Dairy Products. J. Dairy Res. 1997, 64, 409–421. [Google Scholar] [CrossRef]
- Hadef, S.; Idoui, T.; Sifour, M.; Genay, M.; Dary-Mourot, A. Screening of Wild Lactic Acid Bacteria from Algerian Traditional Cheeses and Goat Butter to Develop a New Probiotic Starter Culture. Probiotics Antimicrob. Proteins 2022, 15, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Ayad, E.H.E.; Nashat, S.; El-Sadek, N.; Metwaly, H.; El-Soda, M. Selection of Wild Lactic Acid Bacteria Isolated from Traditional Egyptian Dairy Products According to Production and Technological Criteria. Food Microbiol. 2004, 21, 715–725. [Google Scholar] [CrossRef]
- Delgado, S.; Delgado, T.; Mayo, B. Technological Performance of Several Lactococcus and Enterococcus Strains of Dairy Origin in Milk. J. Food Prot. 2002, 65, 1590–1596. [Google Scholar] [CrossRef] [PubMed]
- Welman, A.D.; Maddox, I.S. Exopolysaccharides from Lactic Acid Bacteria: Perspectives and Challenges. Trends Biotechnol. 2003, 21, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cui, Y.; Qu, X. Exopolysaccharides of Lactic Acid Bacteria: Structure, Bioactivity and Associations: A Review. Carbohydr. Polym. 2019, 207, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Ruas-Madiedo, P.; De Los Reyes-Gavilán, C.G. Invited Review: Methods for the Screening, Isolation, and Characterization of Exopolysaccharides Produced by Lactic Acid Bacteria. J. Dairy Sci. 2005, 88, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Milanovic, V.; Osimani, A.; Garofalo, C.; Belleggia, L.; Maoloni, A.; Cardinali, F.; Mozzon, M.; Foligni, R.; Aquilanti, L.; Clementi, F. Selection of Cereal-Sourced Lactic Acid Bacteria as Candidate Starters for the Baking Industry. PLoS ONE 2020, 15, e0236190. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.T.; Nguyen, T.T.; Bui, D.C.; Hong, P.T.; Hoang, Q.K.; Nguyen, H.T. Exopolysaccharide Production by Lactic Acid Bacteria: The Manipulation of Environmental Stresses for Industrial Applications. AIMS Microbiol. 2020, 6, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.; Ladero, V.; Redruello, B.; Sanchez-Llana, E.; del Rio, B.; Fernandez, M.; Martin, M.C.; Alvarez, M.A. Nucleotide Sequence Alignment of HdcA from Gram-Positive Bacteria. Data Brief 2016, 6, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, J.L.; Troncoso, A.M.; García-Parrilla, M.D.C.; Callejón, R.M. Recent Trends in the Determination of Biogenic Amines in Fermented Beverages—A Review. Anal. Chim. Acta 2016, 939, 10–25. [Google Scholar] [CrossRef] [PubMed]
- EFSA Scientific Opinion on Risk Based Control of Biogenic Amine Formation in Fermented Foods. EFSA J. 2011, 9, 2393. [CrossRef]
- Moniente, M.; García-Gonzalo, D.; Ontañón, I.; Pagán, R.; Botello-Morte, L. Histamine Accumulation in Dairy Products: Microbial Causes, Techniques for the Detection of Histamine-Producing Microbiota, and Potential Solutions. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1481–1523. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, T.; Osimani, A.; Marulo, S.; Cardinali, F.; Mamone, G.; Puppo, C.; Garzón, A.G.; Drago, S.R.; Laurino, C.; Reale, A. Insight into the Role of Lactic Acid Bacteria in the Development of a Novel Fermented Pistachio (Pistacia vera L.) Beverage. Food Biosci. 2023, 53, 102802. [Google Scholar] [CrossRef]
- Osimani, A.; Belleggia, L.; Botta, C.; Ferrocino, I.; Milanović, V.; Cardinali, F.; Haouet, M.N.; Garofalo, C.; Mozzon, M.; Foligni, R.; et al. Journey to the Morpho-Textural Traits, Microbiota, and Volatilome of Ciauscolo PGI Salami. Food Biosci. 2023, 53, 102582. [Google Scholar] [CrossRef]
- Roig-Sagués, A.X.; Molina, A.P.; Hernández-Herrero, M.M. Histamine and Tyramine-Forming Microorganisms in Spanish Traditional Cheeses. Eur. Food Res. Technol. 2002, 215, 96–100. [Google Scholar] [CrossRef]
- Park, Y.K.; Jin, Y.H.; Lee, J.H.; Byun, B.Y.; Lee, J.; Jeong, K.C.C.; Mah, J.H. The Role of Enterococcus Faecium as a Key Producer and Fermentation Condition as an Influencing Factor in Tyramine Accumulation in Cheonggukjang. Foods 2020, 9, 915. [Google Scholar] [CrossRef] [PubMed]
- Tham, W.; Karp, G.; Danielsson-Tham, M.L. Histamine Formation by Enterococci in Goat Cheese. Int. J. Food Microbiol. 1990, 11, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Multiple Bacteriocin Production in Lactic Acid Bacteria. J. Biosci. Bioeng. 2022, 134, 277–287. [Google Scholar] [CrossRef]
- de Niederhäusern, S.; Camellini, S.; Sabia, C.; Iseppi, R.; Bondi, M.; Messi, P. Antilisterial Activity of Bacteriocins Produced by Lactic Bacteria Isolated from Dairy Products. Foods 2020, 9, 1757. [Google Scholar] [CrossRef] [PubMed]
- Alegría, Á.; Delgado, S.; Roces, C.; López, B.; Mayo, B. Bacteriocins Produced by Wild Lactococcus Lactis Strains Isolated from Traditional, Starter-Free Cheeses Made of Raw Milk. Int. J. Food Microbiol. 2010, 143, 61–66. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| pH | Titratable Acidity 1 | Presumptive Lactic Acid Bacteria 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V1 | V2 | V3 | V4 | V1 | V2 | V3 | V4 | V1 | V2 | V3 | V4 | |
| t0 | 6.49 ± 0.07 A | 6.35 ± 0.07 A | 6.48 ± 0.07 A | 6.55 ± 0.07 A | 0.14 ± 0.01 D | 0.16 ± 0.01 C | 0.12 ± 0.01 C | 0.14 ± 0.01 C | 1.85 ± 0.00 d,D | 2.68 ± 0.23 c,C | 3.95 ± 0.11 a,B | 3.26 ± 0.02 b,C |
| t1 | 6.18 ± 0.01 a,B | 6.11 ± 0.01 a,B | 4.53 ± 0.01 c,B | 5.26 ± 0.01 b,B | 0.15 ± 0.01 c,D | 0.18 ± 0.01 c,B,C | 0.62 ± 0.00 a,B | 0.42 ± 0.01 b,B | 7.51 ± 0.05 c,C | 6.29 ± 0.06 d,B | 9.25 ± 0.04 a,A | 8.78 ± 0.07 b,B |
| t2 | 5.81 ± 0.01 a,C | 5.77 ± 0.00 a,C | 4.30 ± 0.01 b,C | 4.34 ± 0.01 b,C | 0.25 ± 0.03 b,C | 0.24 ± 0.01 b,B | 0.74 ± 0.03 a,A | 0.76 ± 0.02 a,A | 8.99 ± 0.00 b,A | 9.55 ± 0.06 a,A | 9.58 ± 0.03 a,A | 9.60 ± 0.01 a,A |
| t3 | 4.70 ± 0.01 a,D | 4.28 ± 0.01 b,D | 4.32 ± 0.00 b,C | 4.25 ± 0.01 b,C | 0.55 ± 0.02 b,B | 0.72 ± 0.03 a,A | 0.74 ± 0.02 a,A | 0.76 ± 0.02 a,A | 8.93 ± 0.01 b,A | 9.59 ± 0.14 a,A | 9.48 ± 0.01 a,A | 9.56 ± 0.09 a,A |
| t4 | 4.40 ± 0.01 E | 4.30 ± 0.00 D | 4.37 ± 0.01 C | 4.27 ± 0.01 C | 0.73 ± 0.02 A | 0.74 ± 0.02 A | 0.73 ± 0.01 A | 0.76 ± 0.05 A | 8.49 ± 0.02 b,B | 9.43 ± 0.02 a,A | 9.43 ± 0.14 a,A | 9.53 ± 0.02 a,A |
| Isolation Source | Isolate Code | Closest Relative | % Identity * | Accession Number ** |
|---|---|---|---|---|
| V1 | 1.1 | Enterococcus lactis | 99.25% | NR_117562 |
| 1.2 | Enterococcus lactis | 99.30% | NR_117562 | |
| 1.3 | Enterococcus faecium | 99.33% | NR_113904 | |
| 1.4 | Enterococcus faecium | 99.37% | NR_113904 | |
| 1.5 | Enterococcus faecium | 99.61% | NR_113904 | |
| 1.6 | Enterococcus durans | 99.58% | NR_036922 | |
| 1.7 | Enterococcus faecium | 99.35% | NR_113904 | |
| 1.8 | Lactococcus cremoris | 99.71% | NR_040954 | |
| 1.9 | Lactococcus cremoris | 98.97% | NR_040954 | |
| 1.10 | Lactococcus cremoris | 99.01% | NR_040954 | |
| 1.11 | Lactococcus cremoris | 99.81% | NR_040954 | |
| 1.12 | Lactococcus cremoris | 98.99% | NR_040954 | |
| 1.13 | Enterococcus faecium | 99.25% | NR_113904 | |
| 1.14 | Enterococcus faecium | 99.79% | NR_113904 | |
| 1.15 | Enterococcus faecium | 99.48% | NR_113904 | |
| V2 | 2.1 | Enterococcus faecalis | 99.64% | NR_115765 |
| 2.2 | Enterococcus faecalis | 99.53% | NR_115765 | |
| 2.3 | Enterococcus durans | 99.58% | NR_036922 | |
| 2.4 | Enterococcus faecalis | 99.22% | NR_115765 | |
| 2.5 | Lactococcus lactis | 99.16% | NR_040955 | |
| 2.6 | Lactococcus lactis | 99.16% | NR_040955 | |
| 2.7 | Lactococcus lactis | 99.66% | NR_040955 | |
| 2.8 | Lactococcus lactis | 99.87% | NR_040955 | |
| 2.9 | Lactococcus lactis | 99.57% | NR_040955 | |
| 2.10 | Lactococcus lactis | 98.85% | NR_040955 | |
| 2.11 | Lactococcus lactis | 99.68% | NR_040955 | |
| 2.12 | Lactococcus lactis | 99.48% | NR_040955 | |
| 2.13 | Lactococcus lactis | 99.36% | NR_040955 | |
| 2.14 | Enterococcus durans | 99.34% | NR_036922 | |
| 2.15 | Lactococcus lactis | 99.62% | NR_040955 | |
| V3 | 3.1 | Lactococcus lactis | 99.30% | NR_040955 |
| 3.2 | Lactococcus lactis | 99.84% | NR_040955 | |
| 3.3 | Lactococcus lactis | 99.71% | NR_040955 | |
| 3.4 | Lactococcus lactis | 99.26% | NR_040955 | |
| 3.5 | Lactococcus lactis | 99.75% | NR_040955 | |
| 3.6 | Lactococcus lactis | 99.72% | NR_040955 | |
| 3.7 | Lactococcus lactis | 99.41% | NR_040955 | |
| 3.8 | Lactococcus lactis | 99.57% | NR_040955 | |
| 3.9 | Enterococcus faecalis | 99.33% | NR_115765 | |
| 3.10 | Enterococcus faecalis | 99.85% | NR_115765 | |
| 3.11 | Enterococcus faecalis | 99.12% | NR_115765 | |
| 3.12 | Lactococcus lactis | 99.71% | NR_040955 | |
| 3.13 | Lactococcus lactis | 99.87% | NR_040955 | |
| 3.14 | Lactococcus lactis | 99.61% | NR_040955 | |
| 3.15 | Lactococcus lactis | 99.49% | NR_040955 | |
| V4 | 4.1 | Enterococcus durans | 99.23% | NR_036922 |
| 4.2 | Enterococcus durans | 99.29% | NR_036922 | |
| 4.3 | Enterococcus durans | 99.42% | NR_036922 | |
| 4.4 | Enterococcus durans | 99.00% | NR_036922 | |
| 4.5 | Lactococcus lactis | 99.54% | NR_040955 | |
| 4.6 | Lactococcus lactis | 99.86% | NR_040955 | |
| 4.7 | Lactococcus lactis | 99.19% | NR_040955 | |
| 4.8 | Lactococcus lactis | 99.26% | NR_040955 | |
| 4.9 | Lactococcus lactis | 99.70% | NR_040955 | |
| 4.10 | Lactococcus lactis | 100.00% | NR_040955 | |
| 4.11 | Lactococcus lactis | 99.61% | NR_040955 | |
| 4.12 | Lactococcus lactis | 99.88% | NR_040955 | |
| 4.13 | Lactococcus lactis | 99.15% | NR_040955 | |
| 4.14 | Lactococcus lactis | 99.73% | NR_040955 | |
| 4.15 | Lactococcus lactis | 99.00% | NR_040955 |
| Isolate Code | Closest Relative | Protease Activity * | Lipase Activity * | Esterase Activity * | EPS Production | hdcA Gene | Antimicrobial Activity |
|---|---|---|---|---|---|---|---|
| 1.1 | Enterococcus lactis | + | + | - | - | - | - |
| 1.2 | Enterococcus lactis | + | - | - | - | - | - |
| 1.3 | Enterococcus faecium | ++ | - | - | - | - | - |
| 1.4 | Enterococcus faecium | + | - | - | - | - | - |
| 1.5 | Enterococcus faecium | + | + | - | - | + | - |
| 1.6 | Enterococcus durans | - | - | - | - | - | - |
| 1.7 | Enterococcus faecium | ++ | - | - | - | - | - |
| 1.8 | Lactococcus cremoris | + | - | - | - | - | - |
| 1.9 | Lactococcus cremoris | + | - | - | - | - | - |
| 1.10 | Lactococcus cremoris | + | - | - | - | - | - |
| 1.11 | Lactococcus cremoris | + | - | - | - | - | - |
| 1.12 | Lactococcus cremoris | ++ | - | - | - | - | - |
| 1.13 | Enterococcus faecium | + | ++ | - | - | - | - |
| 1.14 | Enterococcus faecium | + | ++ | - | - | - | - |
| 1.15 | Enterococcus faecium | + | ++ | - | - | - | - |
| 2.1 | Enterococcus faecalis | + | ++ | - | - | - | - |
| 2.2 | Enterococcus faecalis | + | ++ | - | - | - | - |
| 2.3 | Enterococcus durans | + | - | - | - | - | - |
| 2.4 | Enterococcus faecalis | + | - | - | - | - | - |
| 2.5 | Lactococcus lactis | + | - | - | - | - | - |
| 2.6 | Lactococcus lactis | + | - | - | - | - | - |
| 2.7 | Lactococcus lactis | - | - | - | - | + | - |
| 2.8 | Lactococcus lactis | - | - | - | - | - | - |
| 2.9 | Lactococcus lactis | - | - | - | - | - | - |
| 2.10 | Lactococcus lactis | + | - | - | - | - | - |
| 2.11 | Lactococcus lactis | ++ | - | - | - | + | - |
| 2.12 | Lactococcus lactis | + | - | - | - | - | - |
| 2.13 | Lactococcus lactis | + | - | - | - | - | - |
| 2.14 | Enterococcus durans | ++ | + | - | - | - | - |
| 2.15 | Lactococcus lactis | ++ | - | - | - | - | - |
| 3.1 | Lactococcus lactis | + | - | - | - | - | - |
| 3.2 | Lactococcus lactis | + | - | - | - | - | - |
| 3.3 | Lactococcus lactis | + | - | - | - | - | - |
| 3.4 | Lactococcus lactis | - | - | - | - | - | - |
| 3.5 | Lactococcus lactis | - | - | - | - | - | - |
| 3.6 | Lactococcus lactis | - | - | - | - | - | - |
| 3.7 | Lactococcus lactis | - | - | - | - | - | - |
| 3.8 | Lactococcus lactis | - | - | - | - | - | - |
| 3.9 | Enterococcus faecalis | - | - | - | - | - | - |
| 3.10 | Enterococcus faecalis | + | - | - | - | - | - |
| 3.11 | Enterococcus faecalis | - | - | - | - | - | - |
| 3.12 | Lactococcus lactis | + | - | - | - | - | - |
| 3.13 | Lactococcus lactis | + | - | - | - | + | - |
| 3.14 | Lactococcus lactis | + | - | - | - | - | - |
| 3.15 | Lactococcus lactis | + | - | - | - | - | - |
| 4.1 | Enterococcus durans | + | + | - | - | - | - |
| 4.2 | Enterococcus durans | + | + | - | - | - | - |
| 4.3 | Enterococcus durans | + | - | - | - | - | - |
| 4.4 | Enterococcus durans | + | + | - | - | - | - |
| 4.5 | Lactococcus lactis | - | - | - | - | - | - |
| 4.6 | Lactococcus lactis | - | - | - | - | - | - |
| 4.7 | Lactococcus lactis | - | - | - | - | - | - |
| 4.8 | Lactococcus lactis | - | - | - | - | - | - |
| 4.9 | Lactococcus lactis | - | - | - | - | - | - |
| 4.10 | Lactococcus lactis | + | - | - | - | - | - |
| 4.11 | Lactococcus lactis | - | - | - | - | - | - |
| 4.12 | Lactococcus lactis | + | - | - | - | - | - |
| 4.13 | Lactococcus lactis | + | - | - | - | - | - |
| 4.14 | Lactococcus lactis | + | - | - | - | + | - |
| 4.15 | Lactococcus lactis | ++ | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rampanti, G.; Cantarini, A.; Cardinali, F.; Milanović, V.; Garofalo, C.; Aquilanti, L.; Osimani, A. Technological and Enzymatic Characterization of Autochthonous Lactic Acid Bacteria Isolated from Viili Natural Starters. Foods 2024, 13, 1115. https://doi.org/10.3390/foods13071115
Rampanti G, Cantarini A, Cardinali F, Milanović V, Garofalo C, Aquilanti L, Osimani A. Technological and Enzymatic Characterization of Autochthonous Lactic Acid Bacteria Isolated from Viili Natural Starters. Foods. 2024; 13(7):1115. https://doi.org/10.3390/foods13071115
Chicago/Turabian StyleRampanti, Giorgia, Andrea Cantarini, Federica Cardinali, Vesna Milanović, Cristiana Garofalo, Lucia Aquilanti, and Andrea Osimani. 2024. "Technological and Enzymatic Characterization of Autochthonous Lactic Acid Bacteria Isolated from Viili Natural Starters" Foods 13, no. 7: 1115. https://doi.org/10.3390/foods13071115
APA StyleRampanti, G., Cantarini, A., Cardinali, F., Milanović, V., Garofalo, C., Aquilanti, L., & Osimani, A. (2024). Technological and Enzymatic Characterization of Autochthonous Lactic Acid Bacteria Isolated from Viili Natural Starters. Foods, 13(7), 1115. https://doi.org/10.3390/foods13071115