
Chemical Profile Characterization of Fruit and Vegetable Juices after Fermentation with Probiotic Strains


 ,
,  , , , ,
, , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit and Vegetable Juice Preparation
2.2. Juice Fermentation
2.3. Microbiological Analysis of Juices
2.4. Determination of Total Phenolic Content (TPC)
2.5. Determination of Carotenoid Content
2.6. Antioxidant Activity
2.7. Sugars, Organic Acids and Ethanol Analysis
2.8. LC-QToF/MS Analysis
2.9. Statistical Analysis
3. Results
3.1. Microbiological Analyses
3.2. Total Phenolic Content (TPC) Results
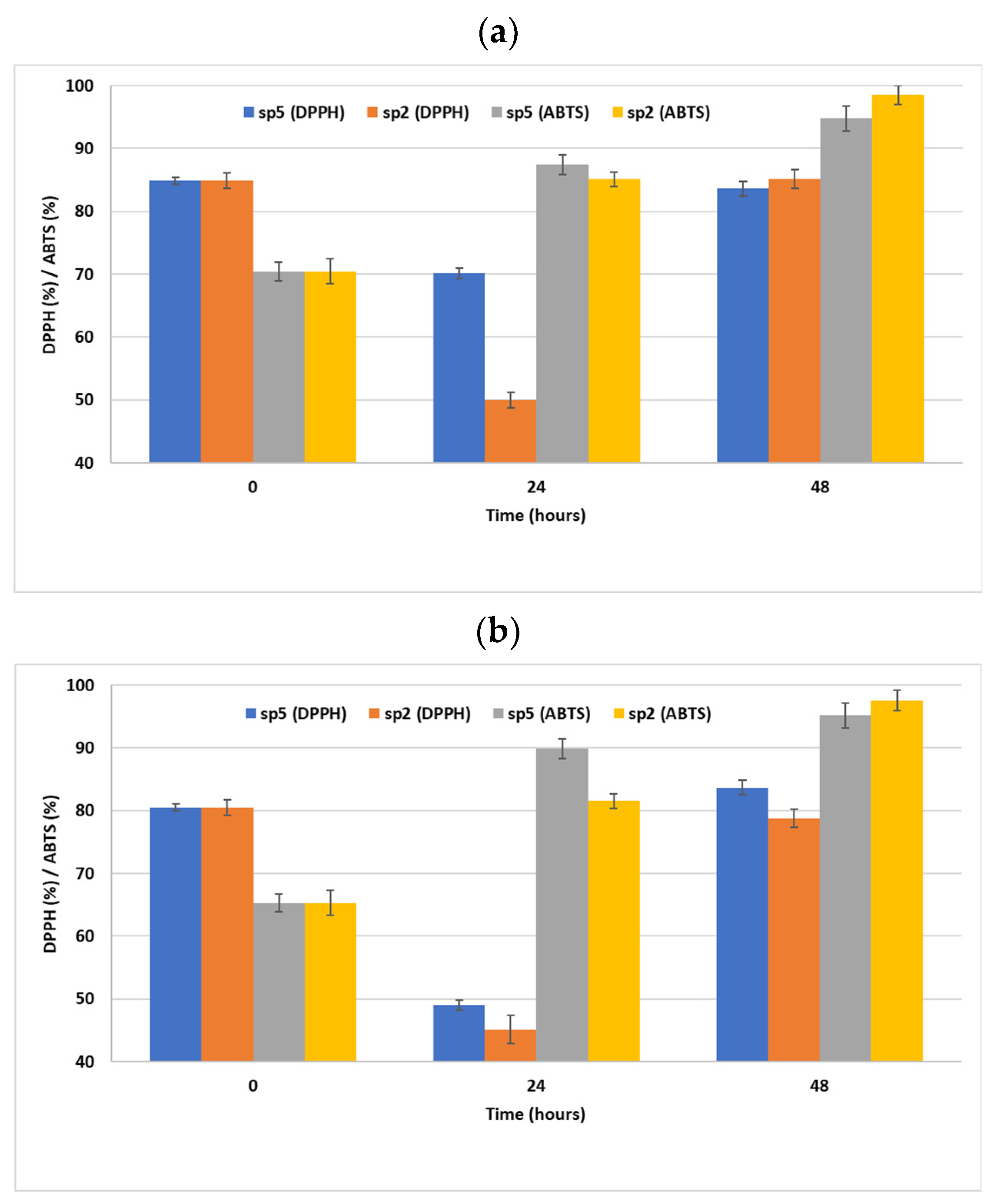
3.3. Antioxidant Activity
3.4. Carotenoid Content of Juice Samples
3.5. HPLC Analysis
3.6. LC-QToF/MS Phenolic Profiling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Prete, R.; Alam, M.K.; Perpetuini, G.; Perla, C.; Pittia, P.; Corsetti, A. Lactic acid bacteria exopolysaccharides producers: A sustainable tool for functional foods. Foods 2021, 10, 1653. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, P.; Wang, J.; Wang, J.; Gu, B.; Ge, F.; Chen, X.D. In vitro gastric digestion and emptying of cooked white and brown rice using a dynamic human stomach system. Food Struct. 2022, 31, 100245. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent research on the health benefits of blueberries and their anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Costa, E.M.; Veiga, M.; Morais, R.M.; Calhau, C.; Pintado, M. Health promoting properties of blueberries: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Inayah, I.; Wibowo, M.S.; Julianti, E.; Suciati, T. Characterization of Lactobacillus zeae as probiotic and starter culture for tamarillo fermented product. Food Sci. Technol. 2022, 42, e54021. [Google Scholar] [CrossRef]
- Gao, B.; Wang, J.; Wang, Y.; Xu, Z.; Li, B.; Meng, X.; Sun, X.; Zhu, J. Influence of fermentation by lactic acid bacteria and in vitro digestion on the biotransformations of blueberry juice phenolics. Food Control 2022, 133, 108603. [Google Scholar] [CrossRef]
- Vivek, K.; Mishra, S.; Pradhan, R.C.; Jayabalan, R. Effect of probiotification with Lactobacillus plantarum MCC 2974 on quality of Sohiong juice. LWT 2019, 108, 55–60. [Google Scholar] [CrossRef]
- Mantzourani, I.; Nikolaou, A.; Kourkoutas, Y.; Alexopoulos, A.; Plessas, S. Biotechnological Features of a Functional Non-Dairy Mixed Juice Fermented with Lacticaseibacillus paracasei SP5. Fermentation 2023, 9, 489. [Google Scholar] [CrossRef]
- Plessas, S.; Mantzourani, I.; Bekatorou, A. Evaluation of Pediococcus pentosaceus SP2 as starter culture on sourdough bread making. Foods 2020, 9, 77. [Google Scholar] [CrossRef] [PubMed]
- Mantzourani, I.; Kazakos, S.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Bekatorou, A.; Plessas, S. Potential of the Probiotic Lactobacillus plantarum ATCC 14917 Strain to Produce Functional Fermented Pomegranate Juice. Foods 2019, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Gemenetzis, E.G.; Alygizakis, N.A. Development and Validation of an HPLC-UV Method for the Determination Bis (2-ethylhexyl) Phthalate Ester in Alcoholic Beverages. Appl. Sci. 2023, 13, 3194. [Google Scholar] [CrossRef]
- Wahed, P.; Razzaq, M.A.; Dharmapuri, S.; Corrales, M. Determination of formaldehyde in food and feed by an in-house validated HPLC method. Food Chem. 2016, 202, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.A. Spectrophotometric determination of antioxidant activity. Redox Rep. 1996, 2, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.J.; Hsien-Jung, C.; Chun-Der, L.I.N.; Yaw-Huei, L.I.N. Antioxidant and antiproliferative activities of water spinach (Ipomoea aquatica Forsk) constituents. Bot. Bull. Acad. Sin. 2005, 46, 99–106. [Google Scholar] [CrossRef]
- Nikolaou, A.; Galanis, A.; Kanellaki, M.; Tassou, C.; Akrida-Demertzi, K.; Kourkoutas, Y. Assessment of free and immobilized kefir culture in simultaneous alcoholic and malolactic cider fermentations. LWT 2017, 76, 67–78. [Google Scholar] [CrossRef]
- Dasenaki, M.E.; Thomaidis, N.S. Multi-residue methodology for the determination of 16 coccidiostats in animal tissues and eggs by hydrophilic interaction liquid chromatography–tandem mass spectrometry. Food Chem. 2019, 275, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, C.; Liu, H.; Liu, J.; Jiao, Z. Profiles of sugar and organic acid of fruit juices: A comparative study and implication for authentication. J. Food Qual. 2020, 2020, 7236534. [Google Scholar] [CrossRef]
- Mahmood, T.; Anwar, F.; Abbas, M.; Boyce, M.C.; Saari, N. Compositional variation in sugars and organic acids at different maturity stages in selected small fruits from Pakistan. Int. J. Mol. Sci. 2012, 13, 1380–1392. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, G.A.; Andres, D.H.; Klaus, D.K. Isolation of polysaccharides from pineapple fruit pulp and their enzymatic liquifaction. Int. Food Res. J. 2010, 17, 193–203. [Google Scholar]
- Nualkaekul, S.; Charalampopoulos, D. Survival of Lactobacillus plantarum in model solutions and fruit juices. Int. J. Food Microbiol. 2011, 146, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Nualkaekul, S.; Salmeron, I.; Charalampopoulos, D. Investigation of the factors influencing the survival of Bifidobacterium longum in model acidic solutions and fruit juices. Food Chem. 2011, 129, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Sun, Y.; Gao, T.; Wu, Y.; Sun, H.; Zhu, Q.; Tao, Y. Fermentation and storage characteristics of “Fuji” apple juice using Lactobacillus acidophilus, Lactobacillus casei and Lactobacillus plantarum: Microbial growth, metabolism of bioactives and in vitro bioactivities. Front. Nutr. 2022, 9, 833906. [Google Scholar] [CrossRef] [PubMed]
- Mantzourani, I.; Terpou, A.; Bekatorou, A.; Plessas, S. Valorization of lactic acid fermentation of pomegranate juice by an acid tolerant and potentially probiotic LAB isolated from kefir grains. Fermentation 2022, 8, 142. [Google Scholar] [CrossRef]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Mangos and their bioactive components: Adding variety to the fruit plate for health. Food Funct. 2017, 8, 3010–3032. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y.; Du, J.; Li, M.L.; Li, C.M. Degradation kinetics and pathways of red raspberry anthocyanins in model and juice systems and their correlation with color and antioxidant changes during storage. LWT 2020, 128, 109448. [Google Scholar] [CrossRef]
- Fröhling, B.; Patz, C.; Dietrich, H.; Will, F. Anthocyanins, total phenolics and antioxidant capacities of commercial red grape juices, black currant and sour cherry nectars. Fruit Process 2012, 3, 100–104. [Google Scholar]
- Candrawinata, V.I.; Blades, B.; Golding, J.; Stathopoulos, C.; Roach, P. Effect of clarification on the polyphenolic compound content and antioxidant activity of commercial apple juices. Int. Food Res. J. 2012, 19, 1055–1061. [Google Scholar]
- Kechinski, C.P.; Guimaraes, P.V.; Norena, C.P.; Tessaro, I.C.; Marczak, L.D. Degradation kinetics of anthocyanin in blueberry juice during thermal treatment. J. Food Sci. 2010, 75, C173–C176. [Google Scholar] [CrossRef]
- Tezcan, F.; Gültekin-Özgüven, M.; Diken, T.; Özçelik, B.; Erim, F.B. Antioxidant activity and total phenolic, organic acid and sugar content in commercial pomegranate juices. Food Chem. 2009, 115, 873–877. [Google Scholar] [CrossRef]
- Almeida, M.M.B.; de Sousa, P.H.M.; Arriaga, Â.M.C.; Do Prado, G.M.; de Calvalho Magalhães, C.E.; Maia, G.A.; de Lemos, T.L.G. Bioactive compounds and antioxidant activity of fresh exotic fruits from Northeastern Brazil. Food Res. Int. 2011, 44, 2155–2159. [Google Scholar] [CrossRef]
- Wern, K.H.; Haron, H.; Keng, C.B. Comparison of total phenolic contents (TPC) and antioxidant activities of fresh fruit juices, commercial 100% fruit juices and fruit drinks. Sains Malays. 2016, 45, 1319–1327. [Google Scholar]
- Ng, C.C.; Wang, C.Y.; Wang, Y.P.; Tzeng, W.S.; Shyu, Y.T. Lactic acid bacterial fermentation on the production of functional antioxidant herbal Anoectochilus formosanus Hayata. J. Biosci. Bioeng. 2011, 111, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Feng, Y.; Yang, N.; Jiang, T.; Xu, H.; Lei, H. Fermentation of kiwifruit juice from two cultivars by probiotic bacteria: Bioactive phenolics, antioxidant activities and flavor volatiles. Food Chem. 2022, 373, 131455. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tao, Y.; Zhang, X.; Shao, S.; Han, Y.; Chu, D.-T.; Xie, G.; Ye, X. Metabolic profile of ginkgo kernel juice fermented with lactic aicd bacteria: A potential way to degrade ginkgolic acids and enrich terpene lactones and phenolics. Process Biochem. 2019, 76, 25–33. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, J.; Fan, L.; Qin, Z.; Chen, Q.; Zhao, L. Antioxidant properties of a vegetable–fruit beverage fermented with two Lactobacillus plantarum strains. Food Sci. Biotechnol. 2018, 27, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Cele, N.P.; Akinola, S.A.; Shoko, T.; Manhevi, V.E.; Remize, F.; Sivakumar, D. The Bioaccessibility and Antioxidant Activities of Fermented Mango Cultivar Juices after Simulated In Vitro Digestion. Foods 2022, 11, 2702. [Google Scholar] [CrossRef] [PubMed]
- Hagi, T.; Kobayashi, M.; Nomura, M. Aerobic conditions increase isoprenoid biosynthesis pathway gene expression levels for carotenoid production in Enterococcus gilvus. FEMS Microbiol. Lett. 2015, 362, fnv075. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xu, H.; Liu, S.; Fang, M.; Wu, Y.; Gong, Z. Study on the bioaccessibility and bioavailability of perchlorate in different food matrices in vitro. Food Chem. 2020, 333, 127470. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.K.; Marriott, H.E. Alcohol Consumption in the New Millennium–Weighing up the Risks and Benefits for Our Health. Nutr. Bullettin 2006, 31, 286–331. [Google Scholar] [CrossRef]
- Nikolaou, A.; Tsakiris, A.; Kanellaki, M.; Bezirtzoglou, E.; Akrida-Demertzi, K.; Kourkoutas, Y. Wine production using free and immobilized kefir culture on natural supports. Food Chem. 2019, 272, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Mantzourani, I.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Bekatorou, A.; Plessas, S. Production of a potentially synbiotic fermented Cornelian cherry (Cornus mas L.) beverage using Lactobacillus paracasei K5 immobilized on wheat bran. Biocatal. Agric. Biotechnol. 2019, 17, 347–351. [Google Scholar] [CrossRef]
- Li, T.; Jiang, T.; Liu, N.; Wu, C.; Xu, H.; Lei, H. Biotransformation of phenolic profiles and improvement of antioxidant capacities in jujube juice by select lactic acid bacteria. Food Chem. 2021, 339, 127859. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Kang, S.; Li, F.; Wang, X.; Xiao, Y.; Wang, J.; Xu, H. Relationship between fruit density and physicochemical properties and bioactive composition of mulberry at harvest. J. Food Compos. Anal. 2022, 106, 104322. [Google Scholar] [CrossRef]
- Jia, T.; Zhao, M.; Pan, L.; Deng, C.; Zou, J.J.; Zhang, X. Effect of phenolic antioxidants on the thermal oxidation stability of high-energy–density fuel. Chem. Eng. Sci. 2022, 247, 117056. [Google Scholar] [CrossRef]
- Multari, S.; Carafa, I.; Barp, L.; Caruso, M.; Licciardello, C.; Larcher, R.; Tuohy, K.; Martens, S. Effects of Lactobacillus spp. on the phytochemical composition of juices from two varieties of Citrus sinensis L. Osbeck: ‘Tarocco’ and ‘Washington navel’. LWT 2020, 125, 109205. [Google Scholar] [CrossRef]
- Tabasco, R.; Sánchez-Patán, F.; Monagas, M.; Bartolomé, B.; Moreno-Arribas, M.V.; Peláez, C.; Requena, T. Effect of grape polyphenols on lactic acid bacteria and bifidobacteria growth: Resistance and metabolism. Food Microbiol. 2011, 28, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Xu, H.; Liu, X.; He, W.; Yuan, F.; Hou, Z.; Gao, Y. Identification of phenolic compounds from pomegranate (Punica granatum L.) seed residues and investigation into their antioxidant capacities by HPLC–ABTS+ assay. Food Res. Int. 2011, 44, 1161–1167. [Google Scholar] [CrossRef]
- Ankolekar, C.; Johnson, K.; Pinto, M.; Johnson, D.; Labbe, R.G.; Greene, D.; Shetty, K. Fermentation of whole apple juice using Lactobacillus acidophilus for potential dietary management of hyperglycemia, hypertension, and modulation of beneficial bacterial responses. J. Food Biochem. 2012, 36, 718–738. [Google Scholar] [CrossRef]
- Attri, S.; Sharma, K.; Raigond, P.; Goel, G. Colonic fermentation of polyphenolics from Sea buckthorn (Hippophae rhamnoides) berries: Assessment of effects on microbial diversity by Principal Component Analysis. Food Res. Int. 2018, 105, 324–332. [Google Scholar] [CrossRef]
- Li, J.; Zhao, W.; Pan, X.; Lao, F.; Liao, X.; Shi, Y.; Wu, J. Improvement of antioxidant properties of jujube puree by biotransformation of polyphenols via Streptococcus thermophilus fermentation. Food Chem. X 2022, 13, 100214. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wu, P.; Dai, Y.; Luo, X.; Manickam, S.; Li, D.; Han, Y.; Show, P.L. Bridge between mass transfer behavior and properties of bubbles under two-stage ultrasound-assisted physisorption of polyphenols using macroporous resin. Chem. Eng. J. 2022, 436, 135158. [Google Scholar] [CrossRef]
- Cai, L.; Wang, W.; Tong, J.; Fang, L.; He, X.; Xue, Q.; Li, Y. Changes of bioactive substances in lactic acid bacteria and yeasts fermented kiwifruit extract during the fermentation. LWT 2022, 164, 113629. [Google Scholar] [CrossRef]
- Xie, H.; Gao, P.; Lu, Z.M.; Wang, F.Z.; Chai, L.J.; Shi, J.S.; Zhang, H.L.; Geng, Y.; Zhang, X.J.; Xu, Z.H. Changes in physicochemical characteristics and metabolites in the fermentation of goji juice by Lactiplantibacillus plantarum. Food Biosci. 2023, 54, 102881. [Google Scholar] [CrossRef]
- Filannino, P.; Cardinali, G.; Rizzello, C.G.; Buchin, S.; De Angelis, M.; Gobbetti, M.; Di Cagno, R. Metabolic responses of Lactobacillus plantarum strains during fermentation and storage of vegetable and fruit juices. Appl. Environ. Microbiol. 2014, 80, 2206–2215. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Fruit Juice | Strains | Time | LAB | Yeasts/Fungi |
|---|---|---|---|---|
| Log CFU/mL | ||||
| 0 h | 8.0 ± 0.0 | |||
| 4 FRUITS | P. pentosaceus SP2 | 24 h | 8.21 a ± 0.15 | nd |
| 48 h | 9.11 b ± 0.23 | nd | ||
| L. paracasei SP5 | 24 h | 8.44 a ± 0.19 | nd | |
| 48 h | 9.45 b ± 0.32 | nd | ||
| 0 h | 8.4 ± 0.0 | |||
| 9 FRUITS | P. pentosaceus SP2 | 24 h | 8.56 a ± 0.21 | nd |
| 48 h | 9.35 b ± 0.25 | nd | ||
| L. paracasei SP5 | 24 h | 8.41 a ± 0.26 | nd | |
| 48 h | 9.54 b ± 0.11 | nd | ||
| Fruit Juice | Strains | Time | TPC |
|---|---|---|---|
| mg/mL GA | |||
| 4 FRUITS | 0 h | 154.34 a ± 4.21 | |
| P. pentosaceus SP2 | 24 h | 164.87 b ± 3.85 | |
| 48 h | 166.05 b ± 3.41 | ||
| L. paracasei SP5 | 24 h | 162.48 b ± 4.10 | |
| 48 h | 171.30 b ± 2.90 | ||
| 9 FRUITS | 0 h | 143.17 a ± 3.42 | |
| P. pentosaceus SP2 | 24 h | 159.43 b ± 4.19 | |
| 48 h | 179.75 c ± 5.28 | ||
| L. paracasei SP5 | 24 h | 147.23 a ± 3.16 | |
| 48 h | 150.28 a ± 4.31 |
| Fruit Juice | Strains | Time | Zeaxanthin + Lutein | β-Carotene | Lycopene |
|---|---|---|---|---|---|
| C (mg/L) | |||||
| 4 FRUITS | 0 h | 0.033 | 0.45 | 0.007 | |
| P. pentosaceus SP2 | 24 h | 0.091 | 0.42 | 0.010 | |
| 48 h | 0.022 | 0.65 | 0.006 | ||
| L. paracasei SP5 | 24 h | 0.033 | 0.37 | 0.007 | |
| 48 h | 0.018 | 0.18 | 0.007 | ||
| 9 FRUITS | 0 h | 0.024 | 0.44 | nd | |
| P. pentosaceus SP2 | 24 h | 0.037 | 0.83 | nd | |
| 48 h | 0.036 | 0.80 | nd | ||
| L. paracasei SP5 | 24 h | 0.031 | 0.48 | nd | |
| 48 h | 0.026 | 0.70 | nd | ||
| Fruit Juice | Strains | Time | Citric Acid | Malic Acid | Lactic Acid | Acetic Acid | Propionic Acid | Total Sugars | Ethanol | pH |
|---|---|---|---|---|---|---|---|---|---|---|
| g/L | %vol | |||||||||
| 4 fruits | 0 h | 5.1 ± 0.1 | 4.1 ± 0.1 | nd | nd | nd | 118.9 ± 1.1 | nd | 3.34 | |
| P. pentosaceus SP2 | 24 h | 4.6 ± 0.2 a | 1.7 ± 0.2 a | 2.9 ± 0.2 a | 0.1 ± 0.1 a | nd | 112.1 ± 0.6 a | nd | 3.37 | |
| 48 h | 4.3 ± 0.2 a | 0.9 ± 0.1 b | 3.3 ± 0.2 a | 0.1 ± 0.1 a | nd | 84.1 ± 0.8 b | 0.5 ± 0.1 | 3.27 | ||
| L. paracasei SP5 | 24 h | 4.7 ± 0.1 a | 4.1 ± 0.2 a | 1.3 ± 0.1 a | nd | nd | 114.6 ± 0.4 a | nd | 3.28 | |
| 48 h | 4.2 ± 0.3 b | 3.7 ± 0.2 a | 1.2 ± 0.1 a | nd | nd | 85.3 ± 2.1 b | 0.5 ± 0.1 | 3.18 | ||
| 9 fruits | 0 h | 4.3 ± 0.1 | 2.9 ± 0.1 | nd | nd | nd | 116.5 ± 0.8 | nd | 3.59 | |
| P. pentosaceus SP2 | 24 h | 4.0 ± 0.1 a | 2.2 ± 0.1 a | 3.5 ± 0.2 a | 0.3 ± 0.1 a | 0.5 ± 0.1 a | 114.6 ± 0.3 a | nd | 3.55 | |
| 48 h | 3.0 ± 0.3 b | 2.0 ± 0.1 a | 4.1 ± 0.3 b | 0.1 ± 0.1 b | 0.5 ± 0.1 a | 84.5 ± 1.9 b | 0.7 ± 0.1 | 3.51 | ||
| L. paracasei SP5 | 24 h | 4.2 ± 0.1 a | 2.3 ± 0.1 a | 2.8 ± 0.1 a | nd | 0.5 ± 0.1 a | 116.7 ± 0.4 a | nd | 3.38 | |
| 48 h | 2.8 ± 0.2 b | 2.1 ± 0.1 a | 2.7 ± 0.2 a | nd | 0.5 ± 0.1 a | 83.3 ± 1.3 b | 0.3 ± 0.1 | 3.37 | ||
| 4 FRUITS | 9 FRUITS | |||||
|---|---|---|---|---|---|---|
| Control | P. pentosaceus SP2 | L. paracasei SP5 | Control | P. pentosaceus SP2 | L. paracasei SP5 | |
| Target Compounds | Concentration (mg/L) | |||||
| 2-cis,4-trans-Abscisic acid | 0.12 | 0.12 | 0.14 | 0.069 | 0.085 | 0.083 |
| Salicylic acid | 0.094 | 0.083 | 0.082 | 0.16 | 0.12 | 0.12 |
| Catechin | 0.45 | 0.44 | 0.36 | 0.61 | 0.68 | 0.66 |
| Epicatechin | 0.70 | 0.85 | 0.67 | 1.2 | 1.2 | 1.2 |
| Chlorogenic acid | 18 | 14 | 12 | 27 | 21 | 21 |
| Ethyl caffeate | 0.022 | 0.030 | 0.027 | ND | ND | ND |
| Ethyl gallate | 0.14 | 0.16 | 0.24 | 0.081 | 0.11 | 0.10 |
| Gallic acid | 1.1 | 1.7 | 1.1 | 0.43 | 0.53 | 0.51 |
| Hesperetin | ND | ND | ND | 0.091 | 0.073 | 0.094 |
| Hydroxytyrosol | 0.14 | 0.13 | 0.13 | 0.074 | 0.072 | 0.063 |
| Phloridzin | 4.7 | 5.6 | 4.8 | 1.8 | 1.5 | 1.4 |
| Polydatin | 0.12 | 0.11 | 0.063 | 0.21 | 0.092 | 0.22 |
| Quercetin | ND | ND | ND | 0.22 | 0.14 | 0.17 |
| Quinic acid | 10 | 22 | 16 | 16 | 23 | 19 |
| Rutin | ND | ND | ND | 0.86 | 1.0 | 0.95 |
| Tyrosol | 0.14 | 5.1 | 2.5 | 0.15 | 2.5 | 1.2 |
| Naringenin | ND | ND | ND | 0.046 | 0.045 | 0.049 |
| 3,4-Dihydroxybenzoic acid (Protocatechuic acid) | 1.0 | 1.5 | ND | 1.1 | 1.1 | 1.0 |
| 2,5-Dihydroxybenzoic acid (Gentisic acid) | 0.10 | 0.17 | 0.092 | 0.31 | 0.31 | 0.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantzourani, I.; Nikolaou, A.; Kourkoutas, Y.; Alexopoulos, A.; Dasenaki, M.; Mastrotheodoraki, A.; Proestos, C.; Thomaidis, N.; Plessas, S. Chemical Profile Characterization of Fruit and Vegetable Juices after Fermentation with Probiotic Strains. Foods 2024, 13, 1136. https://doi.org/10.3390/foods13071136
Mantzourani I, Nikolaou A, Kourkoutas Y, Alexopoulos A, Dasenaki M, Mastrotheodoraki A, Proestos C, Thomaidis N, Plessas S. Chemical Profile Characterization of Fruit and Vegetable Juices after Fermentation with Probiotic Strains. Foods. 2024; 13(7):1136. https://doi.org/10.3390/foods13071136
Chicago/Turabian StyleMantzourani, Ioanna, Anastasios Nikolaou, Yiannis Kourkoutas, Athanasios Alexopoulos, Marilena Dasenaki, Artemis Mastrotheodoraki, Charalampos Proestos, Nikolaos Thomaidis, and Stavros Plessas. 2024. "Chemical Profile Characterization of Fruit and Vegetable Juices after Fermentation with Probiotic Strains" Foods 13, no. 7: 1136. https://doi.org/10.3390/foods13071136
APA StyleMantzourani, I., Nikolaou, A., Kourkoutas, Y., Alexopoulos, A., Dasenaki, M., Mastrotheodoraki, A., Proestos, C., Thomaidis, N., & Plessas, S. (2024). Chemical Profile Characterization of Fruit and Vegetable Juices after Fermentation with Probiotic Strains. Foods, 13(7), 1136. https://doi.org/10.3390/foods13071136