Effect of Neutral Protease on Freshness Quality of Shucked Pacific Oysters at Different Storage Conditions

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of Total Volatile Basic Nitrogen (TVB-N)
2.3. Determination of the Total Viable Counts (TVC)
2.4. Sensory Evaluation
2.5. Determination of Glycogen Content
2.6. pH Measurement
2.7. Determination of ATP-Related Compounds
2.8. Statistical Analysis
3. Results and Discussion
3.1. TVB-N Value
3.2. The Total Viable Counts (TVC)
3.3. Sensory Evaluation
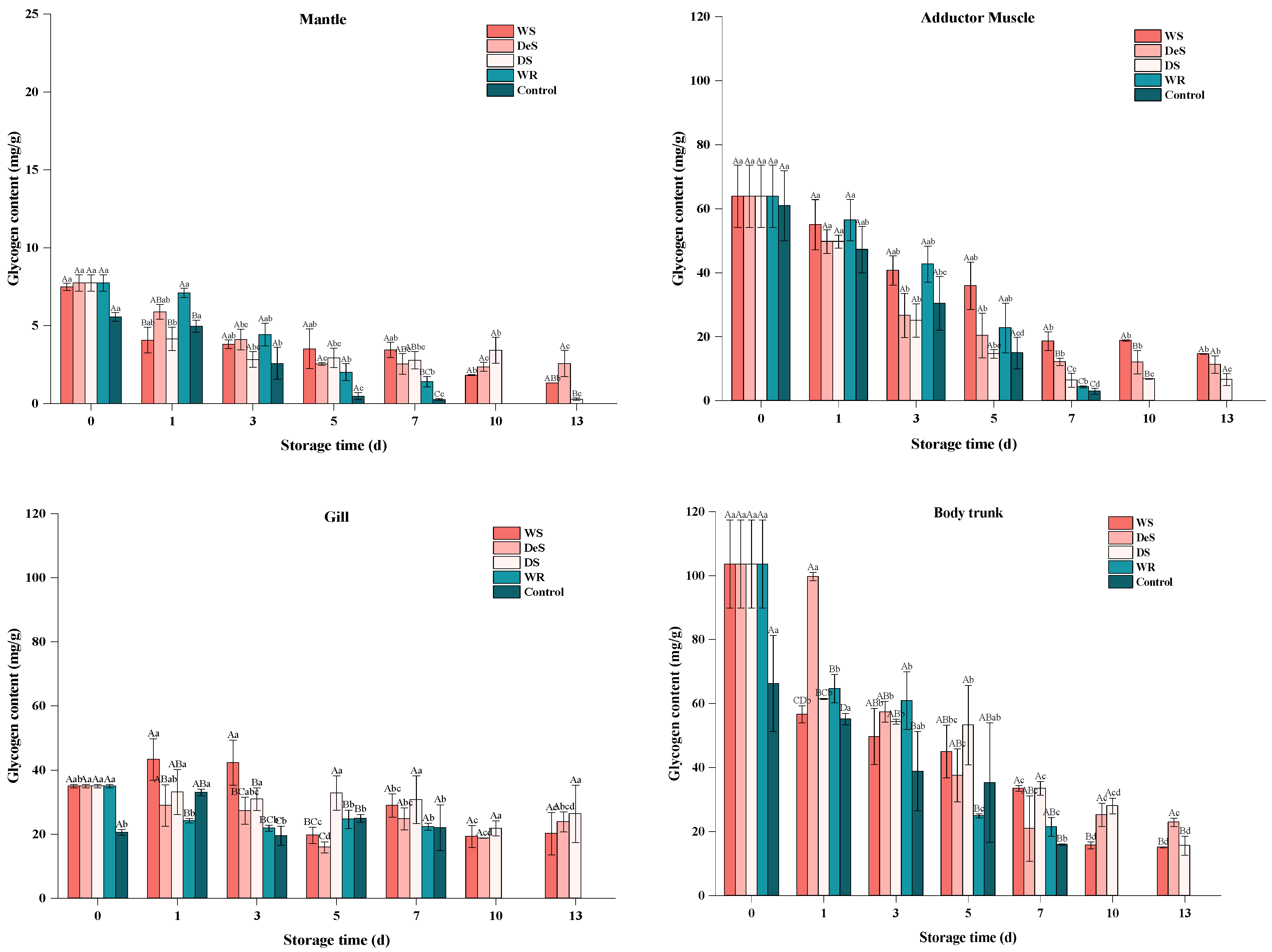
3.4. Glycogen Content
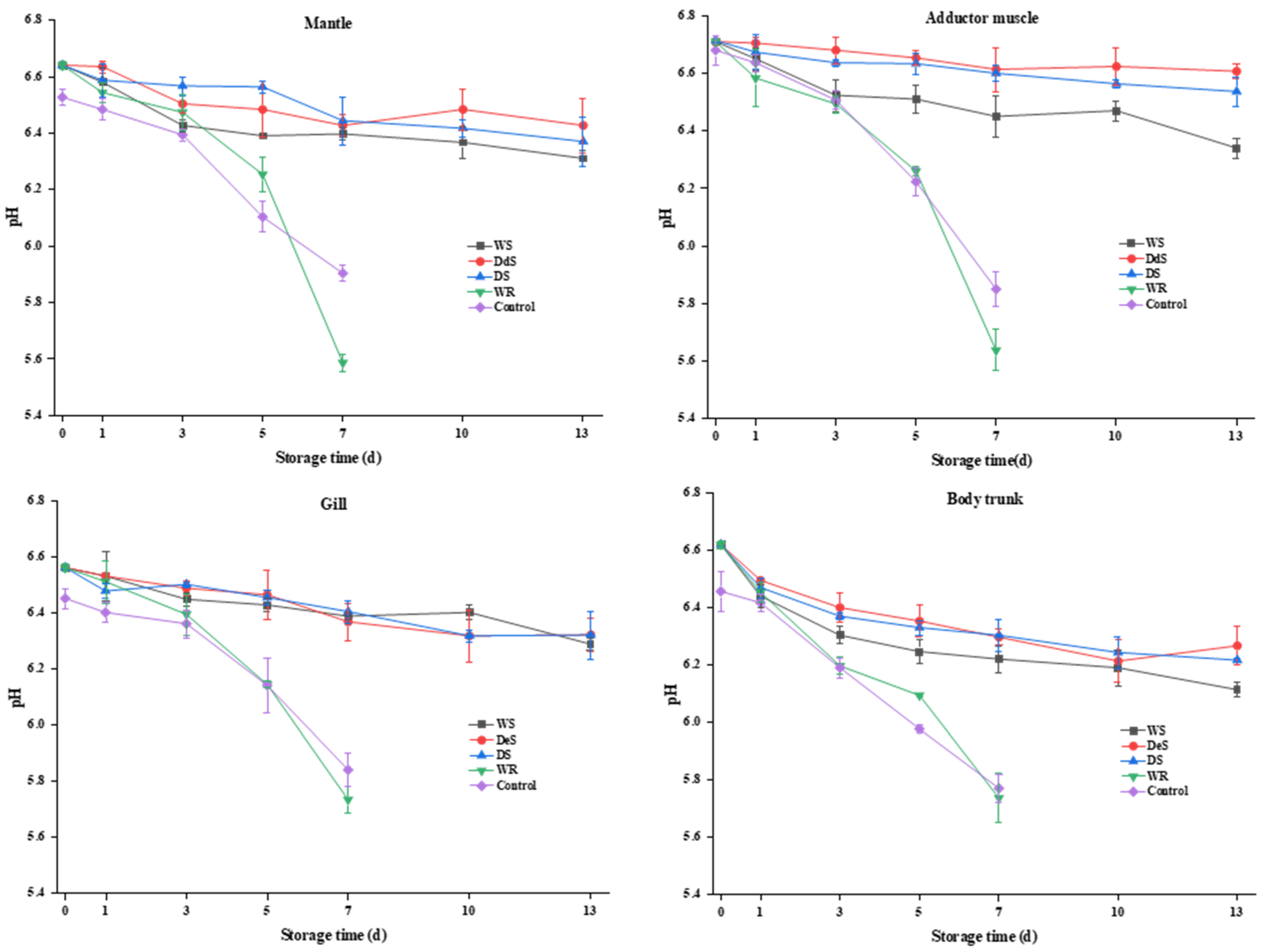
3.5. pH
3.6. ATP-Related Compounds
3.7. Changes in K, K′, and AEC Values in Oyster Tissues during Storage
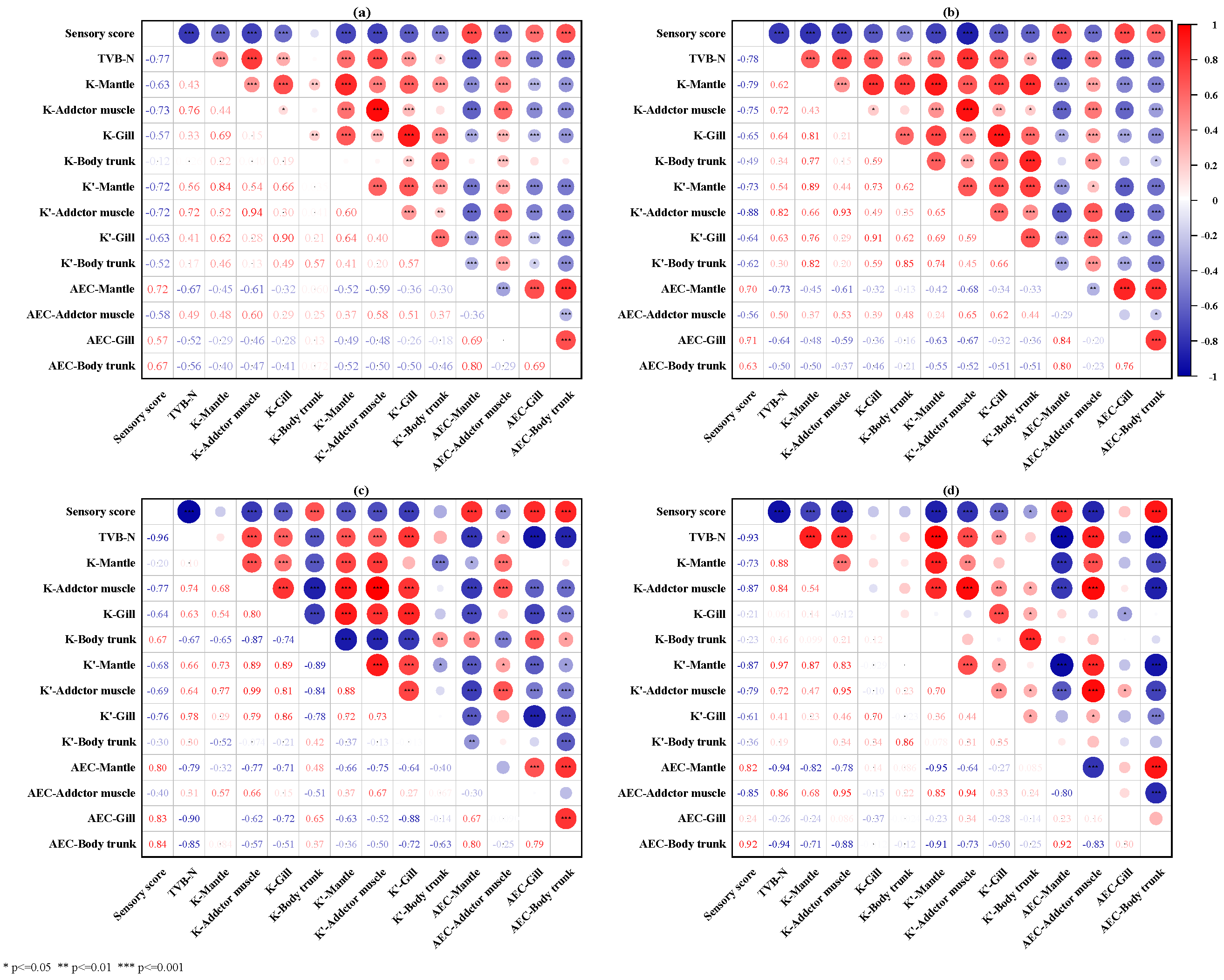
3.8. Relationships among Freshness Indicators of Oyster Tissues, TVB-N Value, and Sensory Score during Storage
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, J.; Meng, L.; Wang, S.; Li, J.; Mao, X. Inactivation of Vibrio Parahaemolyticus and Retardation of Quality Loss in Oyster (Crassostrea gigas) by Ultrasound Processing during Storage. Food Res. Int. 2023, 168, 112722. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.; Wang, F.; Jiang, H.; Zhou, M.; Lin, H. Preparation of Mesoporous Silica Nanosphere-Doped Color-Sensitive Materials and Application in Monitoring the TVB-N of Oysters. Foods 2022, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Mueller Loose, S.; Peschel, A.; Grebitus, C. Quantifying Effects of Convenience and Product Packaging on Consumer Preferences and Market Share of Seafood Products: The Case of Oysters. Food Qual. Prefer. 2013, 28, 492–504. [Google Scholar] [CrossRef]
- Wu, H.; Shi, W.; Huang, H.; Chen, L.; Li, N.; Du, Y.; Pei, N.; Sun, J. The Impact of Flavourzyme on Flavor, Antioxidant Activity, and Sensory Attributes of Salted Grass Carp by Brine Injection and Brining. J. Sci. Food Agric. 2022, 102, 1665–1673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Song, H.; Pan, W.; Chen, W. The Fishy Off-Odor Removal and Umami Enhancing Effect of Enzymatic Hydrolysis of Fish By-Products by Proteases. J. Aquat. Food Prod. 2023, 32, 218–234. [Google Scholar] [CrossRef]
- Feng, Y.; Jiang, S.; Wang, Z.; Li, S.; Liu, Z.; Zeng, M. Oyster Hydrolysate-Zinc Complex Ameliorates Carrageenan-Induced Rat Prostatitis via an Anti-Inflammatory Mechanism and Reduced Oxidative Stress. J. Funct. Foods 2020, 72, 104066. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, F.; Liu, X.; Zhao, M. Particulate Nanocomposite from Oyster (Crassostrea rivularis) Hydrolysates via Zinc Chelation Improves Zinc Solubility and Peptide Activity. Food Chem. 2018, 258, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.S.; Hsiao, Y.T.; Weng, Y.M. Effects of Individual and Block Freezing on the Quality of Pacific Oyster (Crassostrea gigas) during Storage under Different Pretreatment Conditions. Sustainability 2022, 14, 9404. [Google Scholar] [CrossRef]
- Hernández, K.L.; Sedas, V.P.; Dehaibes, S.R.; Valencia, V.S.; Mozo, I.R.; Herrera, D.M.; Primo, A.F.; Serrano, R.U. Improved Microbial Safety of Direct Ozone-Depurated Shellstock Eastern Oysters (Crassostrea virginica) by Superchilled Storage. Front. Microbiol. 2018, 9, 2802. [Google Scholar] [CrossRef]
- Banerjee, R.; Maheswarappa, N.B. Superchilling of Muscle Foods: Potential Alternative for Chilling and Freezing. Crit. Rev. Food Sci. Nutr. 2019, 59, 1256–1263. [Google Scholar] [CrossRef]
- Kaale, L.D.; Eikevik, T.M.; Rustad, T.; Kolsaker, K. Superchilling of Food: A Review. J. Food Eng. 2011, 107, 141–146. [Google Scholar] [CrossRef]
- Tao, Y.; Guo, Y.; Li, J.; Ye, K.; Zhang, Y.; Zeng, X.; Dou, H. Effect of Temperature Fluctuation during Superchilling Storage on the Microstructure and Quality of Raw Pork. Meat Sci. 2023, 198, 109096. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.; Dong, S.; Su, M.; Zhao, Y.; Zeng, M. Physicochemical, Microbiological and Sensory Quality Changes of Tissues from Pacific Oyster (Crassostrea gigas) during Chilled Storage. J. Food Sci. Technol. 2020, 57, 2452–2460. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, M.; Yang, C.; Wan, X.; Ding, H.H.; Shi, Y.; Zhao, C. Bacterial Spoilage Profiles in the Gills of Pacific Oysters (Crassostrea gigas ) and Eastern Oysters (C. virginica ) during Refrigerated Storage. Food Microbiol. 2019, 82, 209–217. [Google Scholar] [CrossRef]
- Wei, H.; Tian, Y.; Lin, Y.; Maeda, H.; Yamashita, T.; Yu, K.; Takaki, K.; Yuan, C. Condition-dependent Adenosine Monophosphate Decomposition Pathways in Striated Adductor Muscle from Japanese Scallop (Patinopecten yessoensis). J. Food Sci. 2020, 85, 1462–1469. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhao, W.; Dong, J.; Zang, J.; Regenstein, J.M.; Jiang, Q.; Xia, W. Multifunctional Bioactive Coatings Based on Water-Soluble Chitosan with Pomegranate Peel Extract for Fish Flesh Preservation. Food Chem. 2022, 374, 131619. [Google Scholar] [CrossRef] [PubMed]
- Bekhit, A.E.D.A.; Holman, B.W.B.; Giteru, S.G.; Hopkins, D.L. Total Volatile Basic Nitrogen (TVB-N) and Its Role in Meat Spoilage: A Review. Trends Food Sci. Technol. 2021, 109, 280–302. [Google Scholar] [CrossRef]
- Cao, R.; Liu, Q.; Yin, B.-z.; Zhu, L.-l. Combined Effect of Ozonated Water and Chitosan on the Shelf-Life of Pacific Oyster (Crassostrea gigas). Innov. Food Sci. Emerg. 2010, 11, 108–112. [Google Scholar]
- Xu, H.; Xiao, N.; Xu, J.; Guo, Q.; Shi, W. Effect of Lactobacillus Plantarum and Flavourzyme on Physicochemical and Safety Properties of Grass Carp during Fermentation. Food Chem. X 2022, 15, 100392. [Google Scholar] [CrossRef]
- Chen, X.; Dong, P.; Li, K.; Zhu, L.; Yang, X.; Mao, Y.; Niu, L.; Hopkins, D.L.; Luo, X.; Liang, R.; et al. Effect of the Combination of Superchilling and Super-Chilled Storage on Shelf-Life and Bacterial Community Dynamics of Beef during Long-Term Storage. Meat Sci. 2022, 192, 108910. [Google Scholar] [CrossRef]
- Kong, S.; Chu, F.; Huang, Y.; Niu, L.; Lai, K. Effects of Salt Concentrations on the Advanced Glycation End-Products in Dried Salted Spanish Mackerel Fillets during Storage. J. Food Meas. Charact. 2022, 16, 3469–3476. [Google Scholar] [CrossRef]
- Cruz-Romero, M.; Kelly, A.L.; Kerry, J.P. Effects of High-Pressure Treatment on the Microflora of Oysters (Crassostrea gigas) during Chilled Storage. Innov. Food Sci. Emerg. 2008, 9, 441–447. [Google Scholar] [CrossRef]
- Songsaeng, S.; Sophanodora, P.; Kaewsrithong, J.; Ohshima, T. Effect of Different Storage Conditions on Quality of White-Scar Oyster (Crassostrea belcheri). Int. Food Res. J. 2010, 17, 491–500. [Google Scholar]
- Chen, J.; Kudo, H.; Kan, K.; Kawamura, S.; Koseki, S. Growth-Inhibitory Effect of D-Tryptophan on Vibrio Spp. in Shucked and Live Oysters. Appl. Environ. Microbiol. 2018, 84, e01543-18. [Google Scholar] [CrossRef] [PubMed]
- Olatunde, O.O.; Benjakul, S.; Vongkamjan, K. Combined Effects of High Voltage Cold Atmospheric Plasma and Antioxidants on the Qualities and Shelf-Life of Asian Sea Bass Slices. Innov. Food Sci. Emerg. 2019, 54, 113–122. [Google Scholar] [CrossRef]
- Costa, C.; Conte, A.; Nobile, M.A. Del Effective Preservation Techniques to Prolong the Shelf Life of Ready-to-eat Oysters. J. Sci. Food Agric. 2015, 94, 2661–2667. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, G.; Xue, P.; Dong, X.; Xia, Y.; Regenstein, J.; Du, M.; Sun, L. Spoilage Microbes’ Effect on Freshness and IMP Degradation in Sturgeon Fillets during Chilled Storage. Food Biosci. 2021, 41, 101008. [Google Scholar] [CrossRef]
- Xiao, N.; Xu, H.; Guo, Q.; Shi, W. Effects of Flavourzyme Addition on Protein Degradation and Flavor Formation in Grass Carp during Fermentation. J. Food Biochem. 2022, 46, e14405. [Google Scholar] [CrossRef]
- Hong, H.; Regenstein, J.M.; Luo, Y. The Importance of ATP-Related Compounds for the Freshness and Flavor of Post-Mortem Fish and Shellfish Muscle: A Review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1787–1798. [Google Scholar] [CrossRef]
- Dai, W.; Yan, C.; Ding, Y.; Wang, W.; Gu, S.; Xu, Z.; Zhou, X.; Ding, Y. Effect of a Chitosan Coating Incorporating Epigallocatechin Gallate on the Quality and Shelf Life of Bighead Carp (Aristichthys nobilis) Fillets during Chilled Storage. Int. J. Biol. Macromol. 2022, 219, 1272–1283. [Google Scholar] [CrossRef]
- Cook, I.; Gimblett, C.G. A Risk Perspective on Fusion Safety Phenomena. Fusion Eng. Des. 1991, 17, 301–306. [Google Scholar] [CrossRef]
- Chung, W.H.; Howieson, J.; Chaklader, M.R. The Ameliorative Effects of Low-Temperature Pasteurization on Physicochemical and Microbiological Quality of Raw Akoya Pearl Oyster (Pinctada fucata). Food Control 2021, 129, 108241. [Google Scholar] [CrossRef]
- Dong, S.; Niu, Y.; Wei, H.; Lin, Y.; Lu, X.; Yamashita, T.; Yu, K.; Takaki, K.; Yuan, C. Effect of Super-Chilling Storage on Maintenance of Quality and Freshness of the Pacific Oyster (Crassostrea gigas). Food Qual. Saf. 2023, 7, fyad008. [Google Scholar] [CrossRef]
- David, E.; Tanguy, A.; Pichavant, K.; Moraga, D. Response of the Pacific Oyster Crassostrea Gigas to Hypoxia Exposure under Experimental Conditions. FEBS J. 2010, 272, 5635–5652. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Score | |||
|---|---|---|---|---|
| 4 | 3 | 2 | 1 | |
| Odor | Strong inherent odor | Light inherent odor | Light putrefaction | Putrefaction |
| Body color | Cream white | White | Tawny/beige | Yellow/light brown |
| Texture | Firm and elastic | Soft and less elastic | Slightly mushy | Mushy |
| Adductor | Pale white Translucent | Light gray Translucent | Light gray Partially opaque | White Opaque |
| Mantle | Brown/Black | Slight fading | Mostly faded | Faded |
| Gill | Gill filaments Well defined | Filaments Less defined | Filaments Poorly defined | Filaments Undefined |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, L.; Yang, W.; Liu, S.; Yuan, C.; Huang, T.; Jia, R.; Wei, H. Effect of Neutral Protease on Freshness Quality of Shucked Pacific Oysters at Different Storage Conditions. Foods 2024, 13, 1273. https://doi.org/10.3390/foods13081273
Su L, Yang W, Liu S, Yuan C, Huang T, Jia R, Wei H. Effect of Neutral Protease on Freshness Quality of Shucked Pacific Oysters at Different Storage Conditions. Foods. 2024; 13(8):1273. https://doi.org/10.3390/foods13081273
Chicago/Turabian StyleSu, Lanxiang, Wenge Yang, Siyang Liu, Chunhong Yuan, Tao Huang, Ru Jia, and Huamao Wei. 2024. "Effect of Neutral Protease on Freshness Quality of Shucked Pacific Oysters at Different Storage Conditions" Foods 13, no. 8: 1273. https://doi.org/10.3390/foods13081273
APA StyleSu, L., Yang, W., Liu, S., Yuan, C., Huang, T., Jia, R., & Wei, H. (2024). Effect of Neutral Protease on Freshness Quality of Shucked Pacific Oysters at Different Storage Conditions. Foods, 13(8), 1273. https://doi.org/10.3390/foods13081273