
Effect of Harvest Time on Non-Volatile Metabolites in Japonica Rice
 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Rice Sample Preparation
2.3. Non-Volatile Metabolite Analysis
2.4. Statistical Analysis
3. Results and Discussion
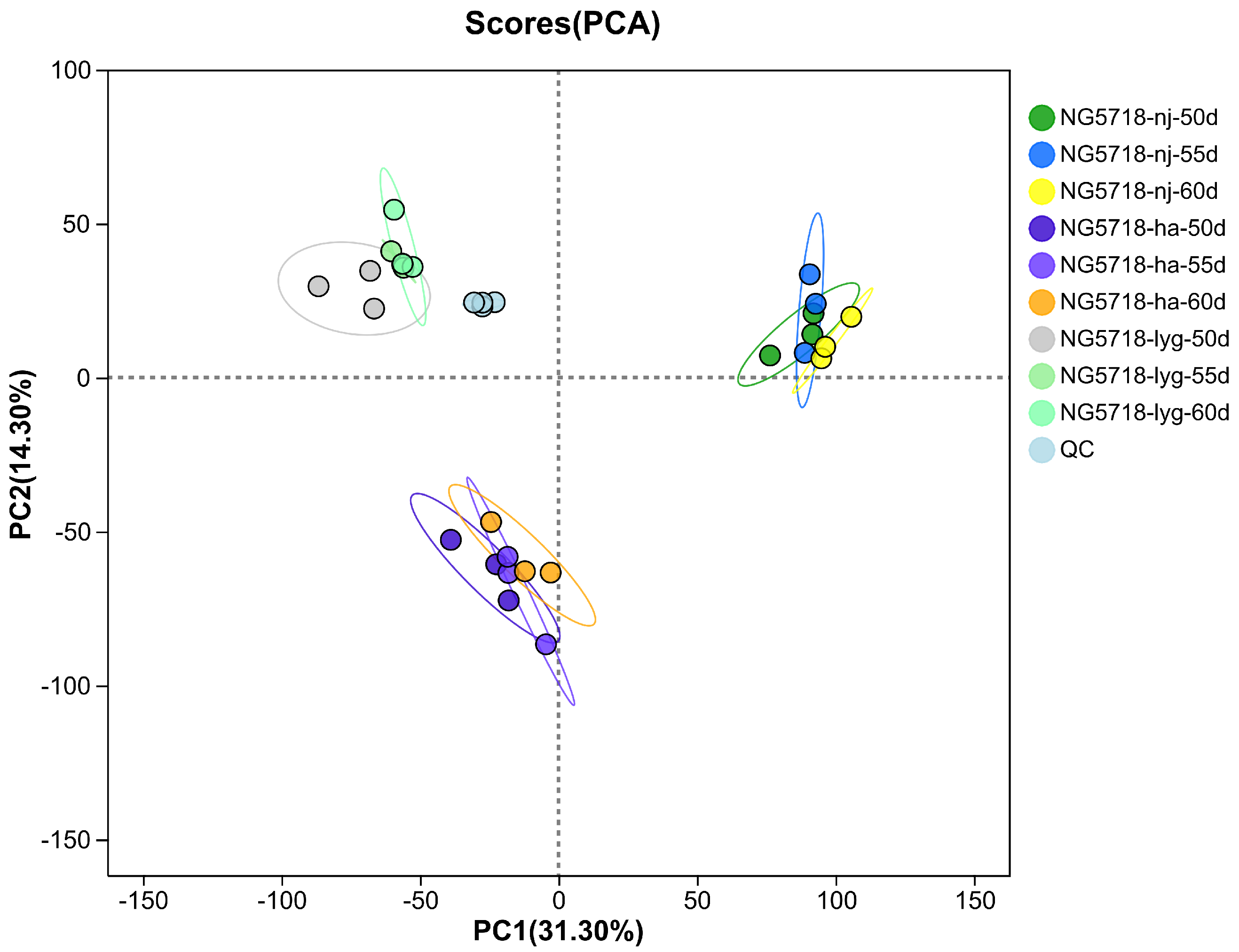
3.1. Data Quality Assessment and Statistical Analysis
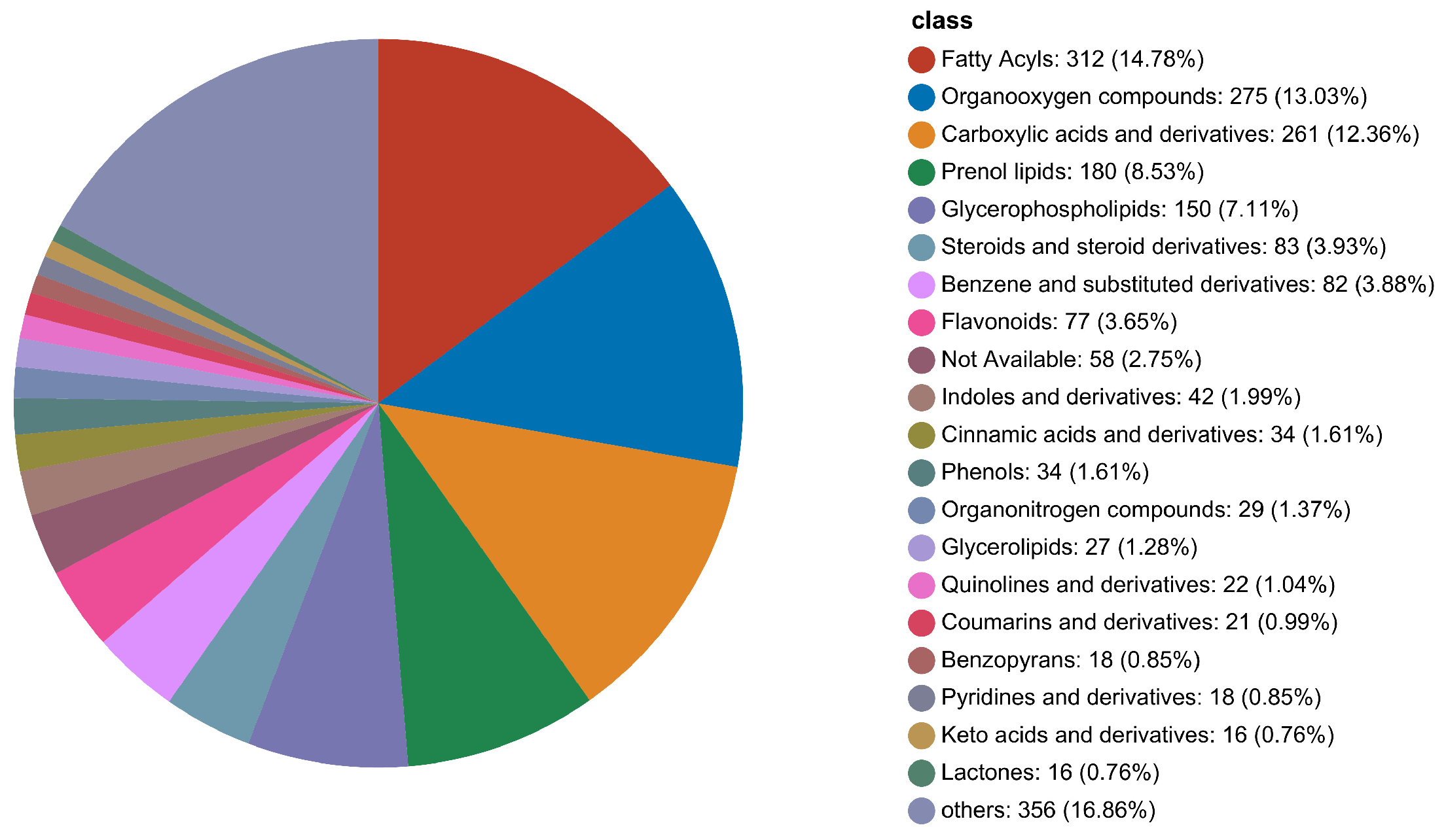
3.2. Compound Classification by HMDB
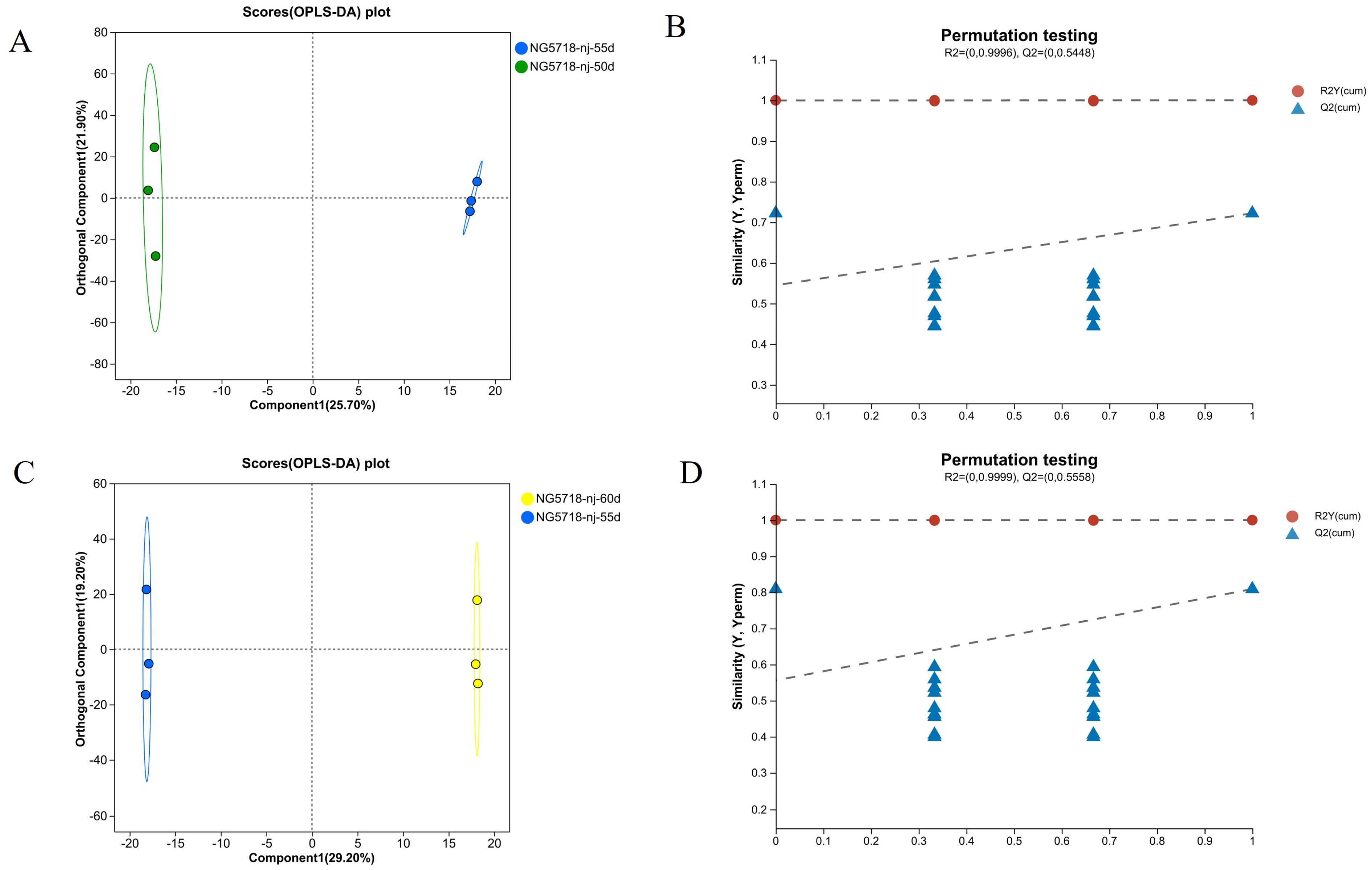
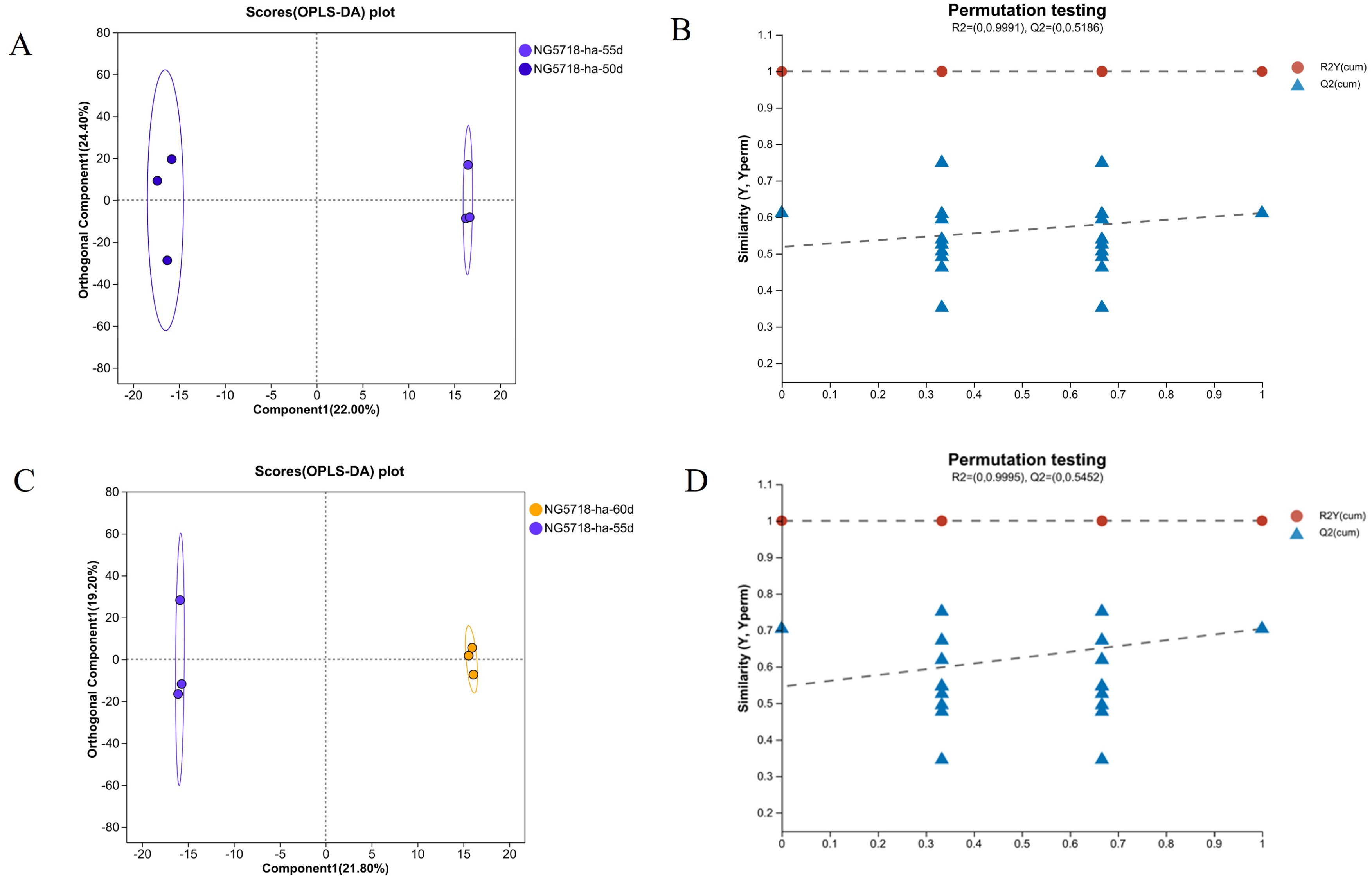
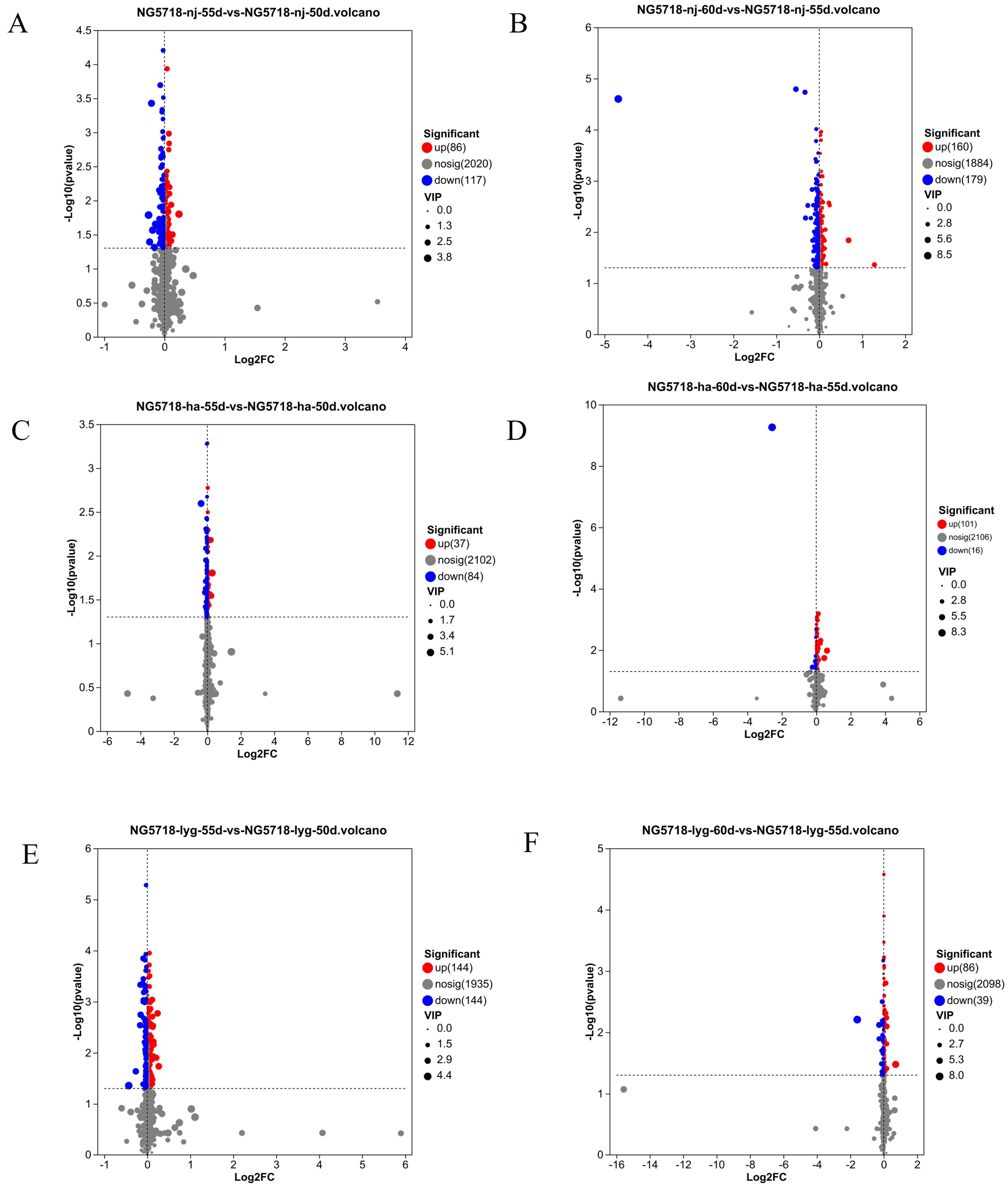
3.3. Screening and Analysis of Differential Metabolites
3.4. KEGG Enrichment Analysis
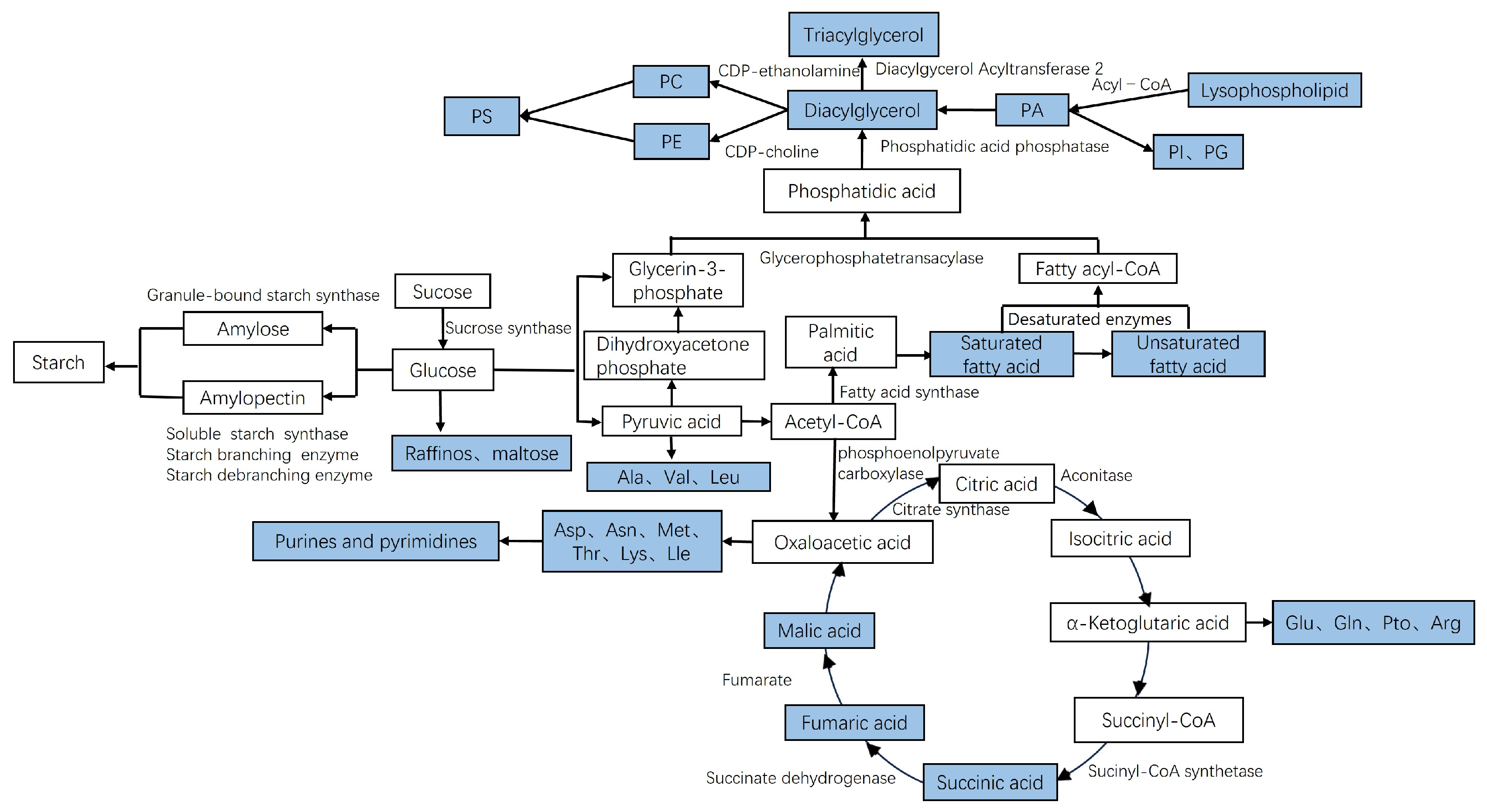
3.5. Metabolic Mechanism Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saleh, A.S.M.; Wang, P.; Wang, N.; Yang, L.; Xiao, Z.G. Brown Rice Versus White Rice: Nutritional Quality, Potential Health Benefits, Development of Food Products, and Preservation Technologies. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1070–1096. [Google Scholar] [PubMed]
- Nirmala, B. Rice Production in Asia: Key to Global Food Security. Proc. Natl. Acad. Sci. India Sect. B 2018, 88, 1323–1328. [Google Scholar]
- Kim, J.G.; Chung, E.S.; Seo, S.; Kim, M.J.; Lee, J.K.; Yoon, S.H.; Lim, Y.C.; Cho, Y.M. Effect of Growth Stage and Variety on the Quality of Whole Crop Rice Silage. J. Korean Soc. Grassl. Forage Sci. 2008, 28, 29–34. [Google Scholar]
- Wang, J.W.; Sun, X.B.; Xu, Y.N.; Wang, Q.; Tang, H.; Zhou, W.Q. The effect of harvest date on yield loss of long and short-grain rice cultivars (Oryza sativa L.) in Northeast China. Eur. J. Agron. 2021, 131, 126382. [Google Scholar] [CrossRef]
- Xangsayasane, P.; Vongxayya, K.; Phongchanmisai, S.; Mitchell, J.; Fukai, S. Rice milling quality as affected by drying method and harvesting time during ripening in wet and dry seasons. Plant Prod. Sci. 2019, 22, 98–106. [Google Scholar]
- Zhang, D.; Shen, D.Y.; Cao, Y.P.; Duan, X.L.; Sun, H. Widely targeted metabolomic approach reveals dynamic changes in non-volatile and volatile metabolites of peanuts during roasting. Food Chem. 2023, 412, 135577. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.L.; Wang, W.S.; Zhang, H.Y.; Liu, X.Q.; Yu, S.B.; Xiong, L.Z.; Luo, J. A Novel Integrated Method for Large-Scale Detection, Identification, and Quantification of Widely Targeted Metabolites: Application in the Study of Rice Metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar]
- Huang, S.S.; Zhang, D.; Wang, Q.; Shang, B.; Liu, J.L.; Xing, X.T.; Hong, Y.; Duan, X.L.; Sun, H. Shotgun lipidomics reveals the changes in phospholipids of brown rice during accelerated aging. Food Res. Int. 2023, 171, 113073. [Google Scholar] [CrossRef]
- Long, X.K.; Guan, C.M.; Wang, L.; Jia, L.T.; Fu, X.J.; Lin, Q.L.; Huang, Z.Y.; Liu, C. Rice Storage Proteins: Focus on Composition, Distribution, Genetic Improvement and Effects on Rice Quality. Rice Sci. 2023, 30, 207–221. [Google Scholar]
- Jin, S.K.; Xu, L.N.; Leng, Y.J.; Zhang, M.Q.; Yang, Q.Q.; Wang, S.L.; Jia, S.W.; Song, T.; Wang, R.A.; Tao, T.; et al. The OsNAC24-OsNAP protein complex activates OsGBSSI and OsSBEI expression to fine-tune starch biosynthesis in rice endosperm. Plant Biotechnol. J. 2023, 21, 2224–2240. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, M.; Ouwerkerk, P.B.F. Molecular and environmental factors determining grain quality in rice. Food Energy Secur. 2012, 1, 111–132. [Google Scholar] [CrossRef]
- Zhang, D.; Duan, X.L.; Wang, Y.Y.; Shang, B.; Liu, H.; Sun, H.; Wang, Y.H. A comparative investigation on physicochemical properties, chemical composition, and in vitro antioxidant activities of rice bran oils from different japonica rice (Oryza sativa L.) varieties. J. Food Meas. Charact. 2021, 15, 2064–2077. [Google Scholar]
- Wang, S.Q.; Yang, S.W.; Tang, Q.H.; Lai, H.F.; Xie, Z.B.; Ye, C.Y.; Tang, X.; Zhao, H.H. Low-temperature stress at the booting stage reduces high-quality aromatic rice quality by improving the albumin and amylose contents. Plant Physiol. Biochem. 2025, 219, 109434. [Google Scholar]
- Qu, S.S.; Wang, J.; Li, G.; Miao, C.; Yan, L.M.; Wang, W. Structural basis for the dual roles of DPW in lipid and UDP-sugar metabolism during rice anther development. Plant Physiol. Biochem. 2025, 222, 109762. [Google Scholar]
- Liang, C.G.; Zhang, Q.; Xu, G.L.; Wang, Y.; Ryu, O.; Li, T. High Temperature During Rice Grain Filling Enhances Aspartate Metabolism in Grains and Results in Accumulation of AspartateFamily Amino Acids and Protein Components. Rice Sci. 2013, 20, 343–348. [Google Scholar] [CrossRef]
- Lin, F.F.; Christopher, R.; Pang, Z.Q.; Zou, J.N.; Lin, S.; Puleng, L.; Zhang, Z.X.; Lin, W.X. Metabolomic analysis reveals differential metabolites and pathways involved in grain chalkiness improvement under rice ratooning. Field Crop Res. 2022, 283, 108521. [Google Scholar] [CrossRef]
- Wang, J.; Guan, H.T.; Zhang, X.L.; Dai, C.J.; Wang, C.L.; Chen, G.L.; Li, K.; Xu, Z.H.; Zhang, R.Y.; Liu, B.H.; et al. Metabonomic and transcriptomic profiling reveals amino acid metabolism affects the quality of premium japonica rice varieties in Northeast China. Food Chem. Mol. Sci. 2024, 9, 100230. [Google Scholar] [CrossRef]
- Zhu, H.B.; Lu, X.Z.; Zhang, K.W.; Xing, Z.P.; Wei, H.Y.; Hu, Q.; Zhang, H.C. Optimum Basic Seedling Density and Yield and Quality Characteristics of Unmanned Aerial Seeding Rice. Agronomy 2023, 13, 1980. [Google Scholar] [CrossRef]
- Cao, J.L.; Chen, X.; Ma, Z.T.; Yu, J.H.; Wang, R.Z.; Zhu, Y.; Xu, F.F.; Hu, Q.; Liu, G.D.; Li, G.Y.; et al. Differences in the Appearance Quality of Soft Japonica Rice with Different Grain Shapes in the Yangtze River Delta and Their Relationship with Grain-Filling. Agronomy 2024, 14, 2377. [Google Scholar] [CrossRef]
- Teng, M.N.; Duan, X.L.; Feng, R.; Zhang, D.; Guo, W.Q.; Sun, H.; Liu, X.Q. Effects of Harvest Time on Grinding Quality, Appearance Quality and Physical and Chemical Quality of Japonica Rice. Foods 2024, 13, 2868. [Google Scholar] [CrossRef]
- Zhou, H.; Li, Y.; Feng, S.J.; Wu, Z.H.; Zheng, D.F.; Feng, N.J. Joint analysis of transcriptome and metabolome revealed the difference in carbohydrate metabolism between super hybrid rice and conventional rice under salt stress. Plant Stress 2023, 10, 100251. [Google Scholar]
- Dellero, Y. Manipulating Amino Acid Metabolism to Improve Crop Nitrogen Use Efficiency for a Sustainable Agriculture. Front. Plant Sci. 2020, 11, 602548. [Google Scholar] [PubMed]
- Dahuja, A.; Kumar, R.R.; Sakhare, A.; Watts, A.; Singh, B.; Goswami, S.; Sachdev, A.; Praveen, S. Role of TP-binding cassette transporters in maintaining plant homeostasis under abiotic and biotic stresses. Physiol. Plant. 2021, 171, 785–801. [Google Scholar] [PubMed]
- Cui, J.; Li, J.L.; Dai, C.H.; Li, L.P. Transcriptome and Metabolome Analyses Revealed the Response Mechanism of Sugar Beet to Salt Stress of Different Durations. Int. J. Mol. Sci. 2022, 23, 9599. [Google Scholar] [CrossRef]
- Huang, S.Y.; Wang, P.; Li, C.J.; Zhou, Q.W.; Huang, T.; Cai, Y.C.; Cheng, Q.; Wang, H.; Zhong, Q.; Chen, Z.H.; et al. Jianmin BianNatural variation of an autophagy-family gene among rice subspecies affects grain size and weight. Crop J. 2024, 12, 121–132. [Google Scholar]
- Ma, D.Y.; Hu, H.Z.; Feng, J.C.; Xu, B.M.; Du, C.Y.; Yang, Y.; Xie, Y.X.; Wang, C.Y. Metabolomics and transcriptomics analyses revealed overexpression of TaMGD enhances wheat plant heat stress resistance through multiple responses. Ecotoxicol. Environ. Saf. 2025, 290, 117738. [Google Scholar]
- Zhang, Z.Y.; Chen, W.C.; Tao, L.; Wei, X.X.; Gao, L.L.; Gao, Y.D.; Suo, J.W.; Yu, W.W.; Hu, Y.Y.; Yang, B.R.; et al. Ethylene treatment promotes umami taste-active amino acids accumulation of Torreya grandis nuts post-harvest by comparative chemical and transcript analyses. Food Chem. 2023, 408, 135214. [Google Scholar]
- Xu, F.; Chen, J.; Wang, W.; Zhu, L.Z. Polybrominated diphenyl ethers (PBDEs) decreased the protein quality of rice grains by disturbing amino acid metabolism. Environ. Pollut. 2024, 353, 124162. [Google Scholar]
- Yao, Y.L.; Xiang, D.H.; Wu, N.; Wang, Y.; Chen, Y.; Yuan, Y.; Ye, Y.; Hu, D.; Zheng, C.; Yan, Y.; et al. Control of rice ratooning ability by a nucleoredoxin that inhibits histidine kinase dimerization to attenuate cytokinin signaling in axillary buds. Mol. Plant 2023, 16, 1911–1926. [Google Scholar]
- Wang, Q.; Zhang, D.; Zhao, L.Y.; Liu, J.L.; Shang, B.; Yang, W.Q.; Duan, X.L.; Sun, H. Metabolomic Analysis Reveals Insights into Deterioration of Rice Quality during Storage. Foods 2022, 11, 1729. [Google Scholar] [CrossRef]
- An, L.; Tao, Y.; Chen, H.; He, M.J.; Xiao, F.; Li, G.H.; Ding, Y.F.; Liu, Z.H. Embryo-Endosperm Interaction and Its Agronomic Relevance to Rice Quality. Front. Plant Sci. 2020, 11, 587641. [Google Scholar] [PubMed]
- Gu, J.F.; Yin, X.Y.; Stomph, T.J.; Wang, H.Q.; Struik, P.C. Physiological basis of genetic variation in leaf photosynthesis among rice (Oryza sativa L.) introgression lines under drought and well-watered conditions. J. Exp. Bot. 2012, 63, 5137–5153. [Google Scholar] [PubMed]
- Huang, L.C.; Tan, H.Y.; Zhang, C.Q.; Li, Q.F.; Liu, Q.Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 100237. [Google Scholar] [PubMed]
- Peng, L.G.; Chen, G.Y.; Tu, Y.B.; Wang, J.; Lan, Y.; Hu, M.M.; Li, C.M.; He, X.M.; Li, T. Effects of Phosphorus Application Rate on Lipid Synthesis and Eating Quality of Two Rice Grains. Agriculture 2022, 12, 667. [Google Scholar] [CrossRef]
- Setsuko, K.; Mayu, E.; Toshihisa, O. The Changes of Amino-Acid Metabolism between Wheat and Rice during Early Growth under Flooding Stress. Int. J. Mol. Sci. 2024, 25, 5229. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | VIP Value | p Value |
|---|---|---|
| DG(i-16:0/0:0/20:3(6,8,11)-OH(5)) | 3.502 | 0.016 |
| PA(i-22:0/i-16:0) | 3.428 | 0.027 |
| Methionyl-Asparagine | 3.263 | 0.041 |
| PC(18:1(9Z)-O(12,13)/18:2(9Z,12Z)) | 3.041 | 0.048 |
| PE-NMe2(18:1(11Z)/18:1(11Z)) | 2.715 | 0.032 |
| 2-Hydroxycineol | 2.491 | 0.012 |
| DG(16:1(9Z)/16:0) | 2.271 | 0.039 |
| N-(4-Amino-2,5-diethoxyphenyl)benzamide | 2.226 | 0.019 |
| PE(18:1(9Z)/18:3(6Z,9Z,12Z)) | 2.196 | 0.012 |
| Enol-phenylpyruvate | 2.177 | 0.007 |
| 13-L-Hydroperoxylinoleic acid | 2.114 | 0.008 |
| 1-(2-Furyl)butan-3-one | 2.086 | 0.022 |
| 5-Hexyl-2-furanoctanoic acid | 2.061 | 0.002 |
| 1,2-Di-(9Z,12Z,15Z-octadecatrienoyl)-3-(galactosyl-alpha-1-6-galactosyl-beta-1)-glycerol | 2.031 | 0.022 |
| (13E)-11a-Hydroxy-9,15-dioxoprost-13-enoic acid | 1.987 | 0.035 |
| Metabolite | VIP Value | p Value |
|---|---|---|
| 5-Phenyl-1,3-oxazinane-2,4-dione | 4.502 | 0.015 |
| 7-Amino-4-methylcoumarin | 3.524 | 0.044 |
| Shikimic acid | 2.842 | 0.003 |
| PE(18:0/18:4(6Z,9Z,12Z,15Z)) | 2.679 | 0.042 |
| Ergocalciferol | 2.563 | 0.035 |
| (S)-(+)-1-(p-Hydroxy-trans-cinnamoyl)-glycerol | 2.540 | 0.003 |
| Valeric acid | 2.369 | 0.024 |
| N-[(2R)-6,7-Dihydroxy-3-oxo-1-sulfanylheptan-2-yl]acetamide | 2.347 | 0.010 |
| N-Palmitoyl lysine | 2.323 | 0.021 |
| Glutamine-glutamate | 2.310 | 0.004 |
| Subaphylline | 2.289 | 0.019 |
| Methylarbutin | 2.260 | 0.021 |
| 5-Megastigmen-7-yne-3,9-diol 3-glucoside | 2.254 | 0.015 |
| 1-Octen-3-yl glucoside | 2.200 | 0.013 |
| N-Ethyl trans-2-cis-6-nonadienamide | 2.184 | 0.011 |
| Metabolite | VIP Value | p Value |
|---|---|---|
| 3-(Octyloxy)propan-1-amine | 4.278 | 0.016 |
| TG(15:0/22:6(4Z,7Z,10Z,13Z,16Z,19Z)/14:1(9Z)) | 3.095 | 0.007 |
| 1-Amino-2-methylanthraquinone | 2.908 | 0.026 |
| 15-F2t-IsoP | 2.843 | 0.027 |
| DG(16:0/18:3(9Z,12Z,15Z)) | 2.520 | 0.008 |
| Cotinine glucuronide | 2.498 | 0.023 |
| Ne,Ne dimethyllysine | 2.316 | 0.005 |
| Indole-3-acetyl-myo-inositol | 2.116 | 0.034 |
| Fructosyl valine | 2.069 | 0.027 |
| Indole-3-acetyl-tyrosine | 1.983 | 0.023 |
| 1-O-E-Cinnamoyl-(6-arabinosylglucose) | 1.821 | 0.005 |
| DG(18:1(9Z)/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | 1.759 | 0.012 |
| 6-Fluoro-DL-tryptophan | 1.748 | 0.023 |
| 1-Myristoyl-sn-Glycerol 3-Phosphate | 1.741 | 0.044 |
| 5-(6-hydroxy-6-methyloctyl)furan-2(5H)-one | 1.677 | 0.041 |
| Metabolite | VIP Value | p Value |
|---|---|---|
| 11-Hydroxy-9-tridecenoic acid | 8.283 | 5.481 |
| Glucomannan | 4.936 | 0.018 |
| DG(16:0/18:3(9Z,12Z,15Z)) | 3.169 | 0.001 |
| 5-Methyleriodictyol 7-[glucosyl-(1->4)-galactoside] | 2.961 | 0.009 |
| PC(18:4(6Z,9Z,12Z,15Z)/18:2(9Z,12Z)) | 2.498 | 0.011 |
| Cytosine | 2.324 | 0.001 |
| Beta-D-glucosamine | 2.280 | 0.003 |
| Sn-glycero-3-phosphoethanolamine | 2.271 | 0.002 |
| Cytidine | 2.226 | 0.017 |
| LL-2,6-Diaminopimelic acid | 2.176 | 0.043 |
| N-Feruloylaspartic acid | 2.098 | 0.010 |
| 1-Methylxanthine | 2.051 | 0.020 |
| N-(4-Amino-2,5-diethoxyphenyl)benzamide | 2.023 | 0.026 |
| L-Glycine | 2.023 | 0.024 |
| Vitamin A2 | 1.979 | 0.001 |
| Metabolite | VIP Value | p Value |
|---|---|---|
| Glucomannan | 4.197 | 0.044 |
| 1-(3-Methyl-2-butenoyl)-6-apiosylglucose | 3.212 | 0.018 |
| Desglymidodrine | 3.075 | 0.023 |
| (7′R,8′R)-4,7′-Epoxy-3′,5-dimethoxy-4′,9,9′-lignanetriol 9′-glucoside | 3.042 | 0.002 |
| N-Oleoyl asparagine | 2.713 | 0.003 |
| Indole-3-acetyl-tyrosine | 2.621 | 0.000 |
| Indole-3-acetamide | 2.576 | 0.006 |
| DG(18:4(6Z,9Z,12Z,15Z)/15:0) | 2.502 | 0.002 |
| PC(18:1(11Z)/18:1(11Z)) | 2.498 | 0.040 |
| (3S,7E,9S)-9-Hydroxy-4,7-megastigmadien-3-one 9-glucoside | 2.490 | 0.012 |
| Indole-3-acetyl-beta-4-D-glucose | 2.486 | 0.000 |
| Indole-3-acetyl-myo-inositol | 2.365 | 0.001 |
| (3beta,6beta)-Furanoeremophilane-3,6-diol 6-acetate | 2.336 | 0.003 |
| L-Tryptophan | 2.285 | 0.014 |
| Oryzalide B | 2.163 | 0.008 |
| Metabolite | VIP Value | p Value |
|---|---|---|
| PE(18:3(9Z,12Z,15Z)/20:3(5Z,8Z,11Z)) | 6.753 | 0.033 |
| MGDG(18:3/18:2) | 4.673 | 0.008 |
| DG(20:3(6,8,11)-OH(5)/16:0) | 4.395 | 0.008 |
| DG(18:2/18:1/0:0) | 4.275 | 0.006 |
| DG(15:0/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | 3.682 | 0.002 |
| Tyrosyl-proline | 3.356 | 0.013 |
| 2-Hydroxypyridine | 3.056 | 0.015 |
| DGDG(18:2/18:2) | 2.998 | 0.003 |
| 13-L-Hydroperoxylinoleic acid | 2.385 | 0.048 |
| Indole-3-acetamide | 2.307 | 0.008 |
| Erucamide | 2.189 | 0.014 |
| Cyclohexaneundecanoic acid | 2.165 | 0.022 |
| LysoPE(0:0/20:1(11Z)) | 2.153 | 0.004 |
| Glucoobtusifolin | 1.945 | 0.002 |
| Cis-p-Coumaric acid 4-[apiosyl-(1->2)-glucoside] | 1.876 | 0.046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teng, M.; Xing, X.; Jiang, P.; Duan, X.; Zhang, D.; Sun, H.; Zhao, C.; Liu, X.; Yao, Z.; Kawano, M. Effect of Harvest Time on Non-Volatile Metabolites in Japonica Rice. Foods 2025, 14, 1224. https://doi.org/10.3390/foods14071224
Teng M, Xing X, Jiang P, Duan X, Zhang D, Sun H, Zhao C, Liu X, Yao Z, Kawano M. Effect of Harvest Time on Non-Volatile Metabolites in Japonica Rice. Foods. 2025; 14(7):1224. https://doi.org/10.3390/foods14071224
Chicago/Turabian StyleTeng, Mengnan, Xiaoting Xing, Pengli Jiang, Xiaoliang Duan, Dong Zhang, Hui Sun, Chunfang Zhao, Xingquan Liu, Zhigang Yao, and Motonobu Kawano. 2025. "Effect of Harvest Time on Non-Volatile Metabolites in Japonica Rice" Foods 14, no. 7: 1224. https://doi.org/10.3390/foods14071224
APA StyleTeng, M., Xing, X., Jiang, P., Duan, X., Zhang, D., Sun, H., Zhao, C., Liu, X., Yao, Z., & Kawano, M. (2025). Effect of Harvest Time on Non-Volatile Metabolites in Japonica Rice. Foods, 14(7), 1224. https://doi.org/10.3390/foods14071224