Antioxidant Activities of Solanum nigrum L. Leaf Extracts Determined in In Vitro Cellular Models


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Plant Material and Extraction Procedures
2.2.2. Determination of Total Phenolic Content
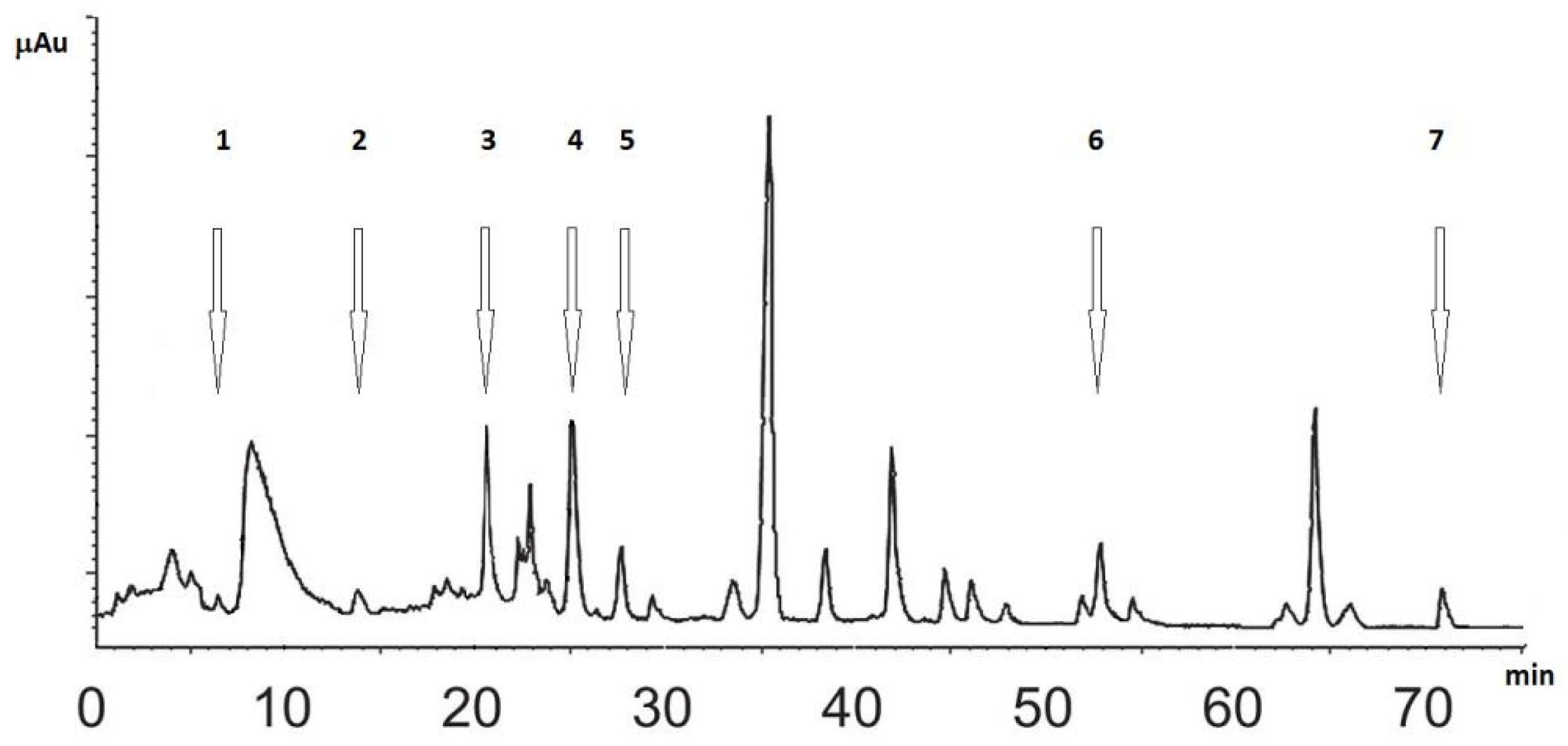
2.2.3. Chromatography
2.2.4. Determination of Antioxidant Activity in an In Vitro Cellular Free System
2.2.5. Determination of Antioxidant Activity within an In Vitro Cellular System
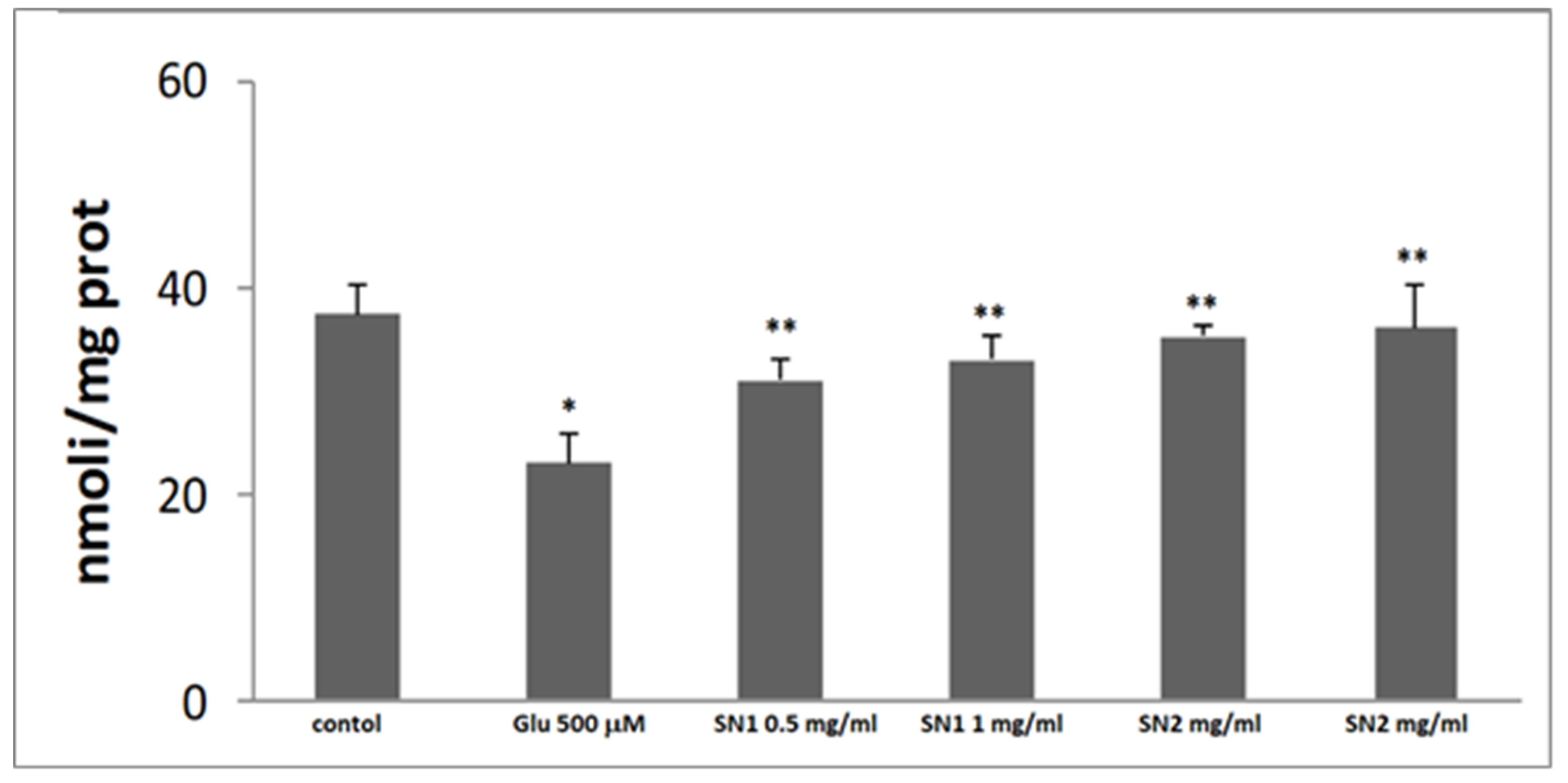
2.2.6. Glutathione Measurement
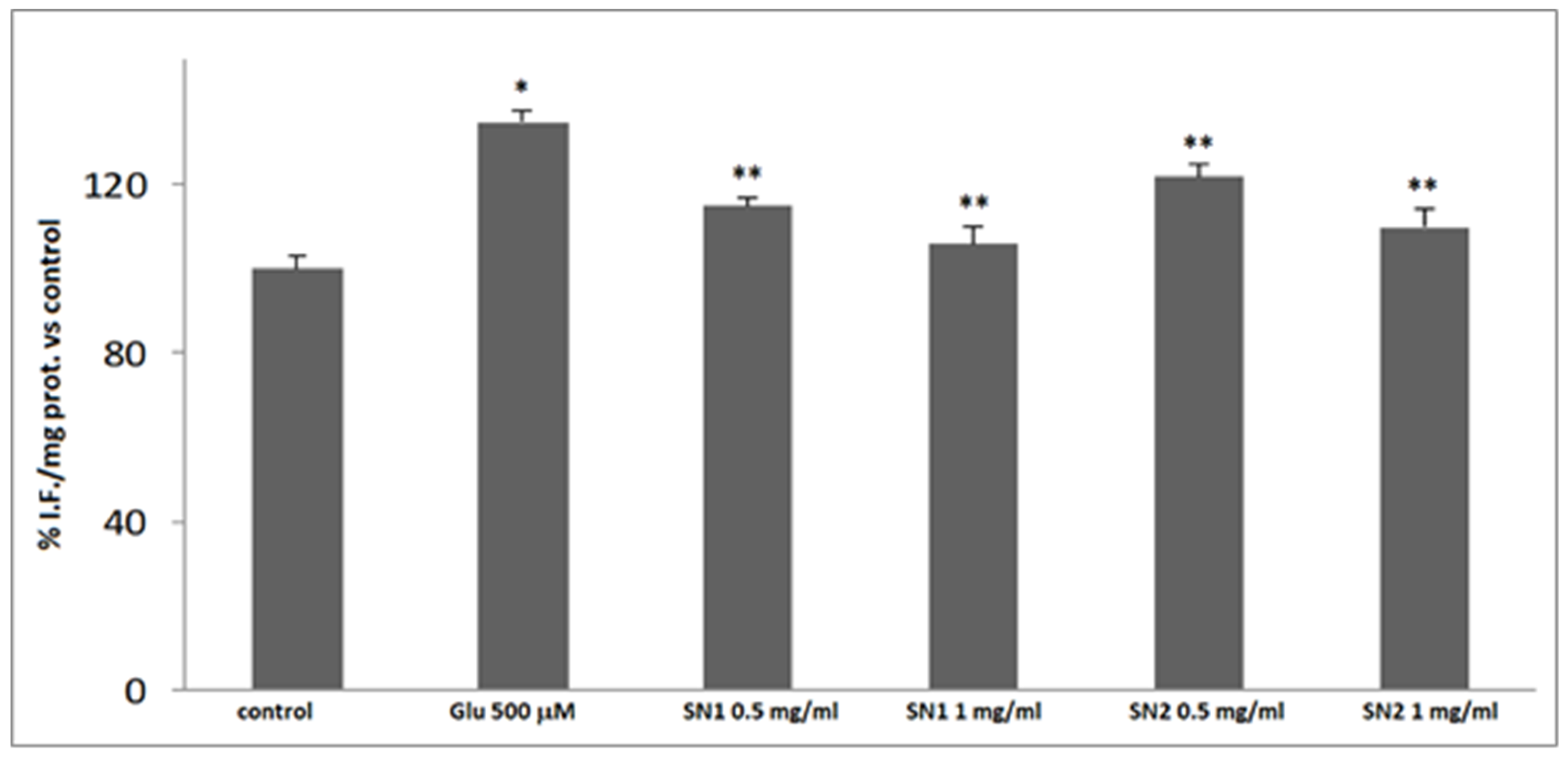
2.2.7. ROS Levels Determination
2.3. Statistical Analysis
3. Results
3.1. Analysis of SN Extracts
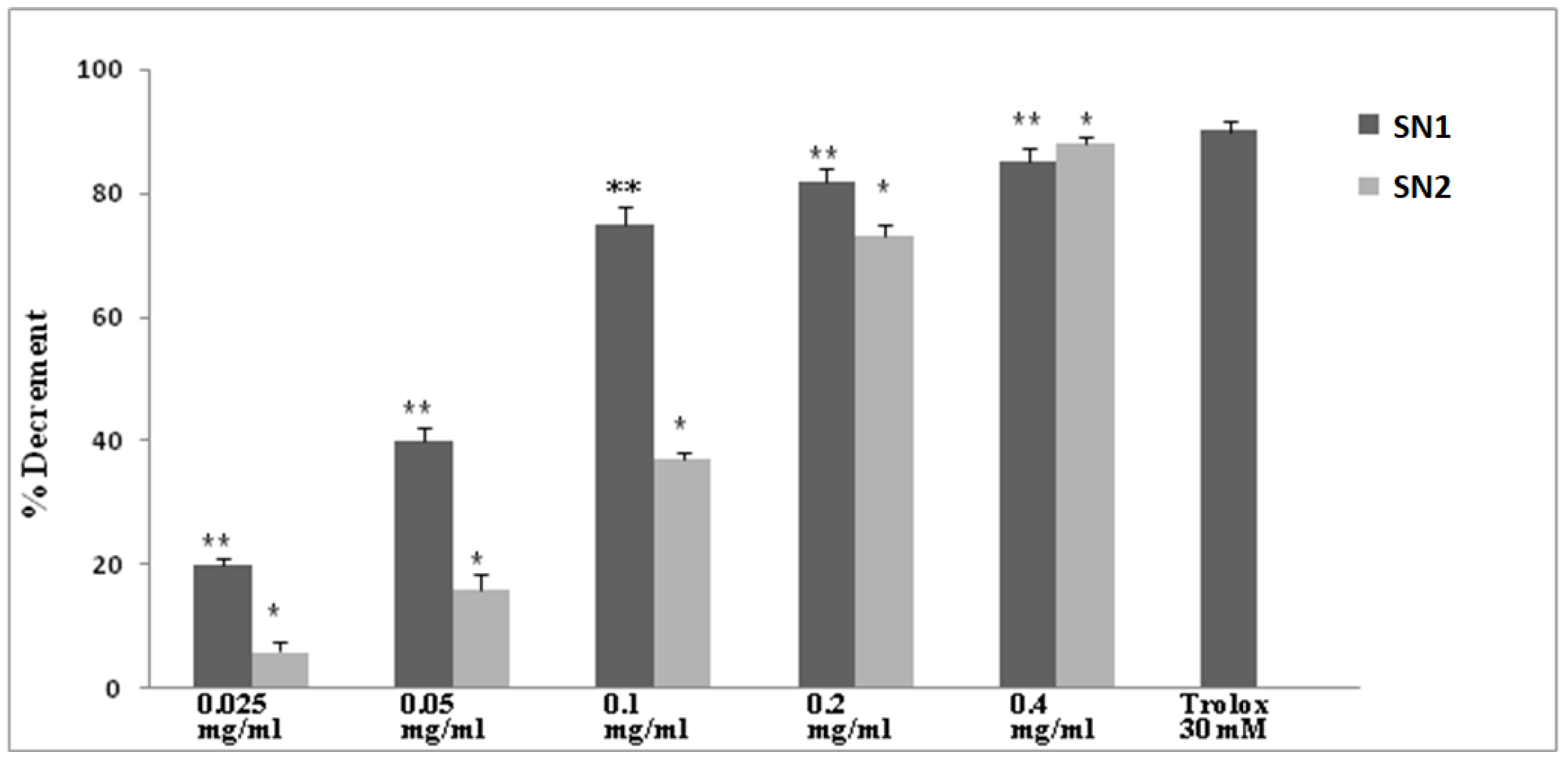
3.2. Antioxidant Activity in a Cellular Free System
3.3. Antioxidant Activity in the Cellular System
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ATP | Adenosine Triphosphate |
| DCF | 2′,7′-dichlorofluorescein |
| DCFH-DA | 2′,7′-dichlorofluorescein diacetate |
| DIV | days in vitro |
| DMEM | Dulbecco’s Modified Eagle’s medium |
| DPPH | 1,1-Diphenyl-2-Picrylhydrazyl radical |
| EDTA | ethylenediaminetetracetic acid |
| FBS | Heat Inactivated Fetal Bovine Serum |
| FITC | fluorescein isothiocyanate |
| GFAP | Glial Fibrillary Acidic Protein |
| GSH | reduced glutathione |
| HPLC | High performance liquid chromatography |
| MTT | 3(4,5-dimethyl-thiazol-2-yl)2,5-diphenyl-tetrazolium bromide |
| NADH | β-Nicotinamide adenine dinucleotide |
| O2•− | superoxide anion |
| •OH | hydroxyl radical |
| ROO• | peroxyl radical |
| ROS | reactive oxygen species |
| SOD | Superoxide dismutase |
| SN | Solanum Nigrum L |
| XO | Xanthine oxidase |
References
- Särkinen, T.; Poczai, P.; Barboza, G.E.; van der Weerden, G.M.; Baden, M.; Knapp, S. A revision of the Old World Black Nightshades (Morelloid clade of Solanum L., Solanaceae). PhytoKeys 2018, 106, 1–223. [Google Scholar]
- Jagatheeswari, D.; Bharathi, T.; Sheik Jahabar Ali, H. Black Night Shade (Solanum nigrum L.)-An Updated Overview. Int. J. Pharm. Biol. Arch. 2013, 4, 288–295. [Google Scholar]
- Akubugwo, I.E.; Obasi, A.N.; Ginika, S.C. Nutritional Potential of the Leaves and Seeds of Black Nightshade-Solanum nigrum L. Var virginicum from Afikpo-Nigeria. Pak. J. Nutr. 2007, 6, 323–326. [Google Scholar] [CrossRef]
- Leporatti, M.L.; Ghedira, K. Comparative analysis of medicinal plants used in traditional medicine in Italy and Tunisia. J. Ethnobiol. Ethnomed. 2009, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Sharma, A.; Gupta, S.; Sarethy, I.P.; Gabrani, R. Solanum nigrum: Current perspectives on therapeutic properties. Altern. Med. Rev. 2011, 16, 78–85. [Google Scholar] [PubMed]
- Wang, Z.; Li, J.; Ji, Y.; An, P.; Zhang, S.; Li, Z. Traditional herbal medicine: A review of potential of inhibitory hepatocellular carcinoma in basic research and clinical trial. Evid. Based Complement. Altern. Med. 2013, 2013, 268963. [Google Scholar] [CrossRef]
- Patel, A.; Biswas, S.; Shoja, M.H.; Ramalingayya, G.V.; Nandakumar, K. Protective effects of aqueous extract of Solanum nigrum Linn. leaves in rat models of oral mucositis. Sci. World J. 2014, 2014, 345939. [Google Scholar]
- Wang, C.K.; Lin, Y.F.; Tai, C.J. Integrated treatment of aqueous extract of Solanum nigrum-potentiated cisplatin- and doxorubicin-induced cytotoxicity in human hepatocellular carcinoma cells. Evid. Based Complement. Alternat. Med. 2015, 2015, 675270. [Google Scholar]
- Wang, C.W.; Chen, C.L.; Wang, C.K. Cisplatin-, doxorubicin-, and docetaxel-induced cell death promoted by the aqueous extract of Solanum nigrum in human ovarian carcinoma cells. Integr. Cancer Ther. 2015, 14, 546–555. [Google Scholar] [CrossRef]
- Tai, C.J.; Wang, C.K.; Chang, Y.J.; Lin, C.S.; Tai, C.J. Aqueous extract of Solanum nigrum leaf activates autophagic cell death and enhances docetaxel-induced cytotoxicity in human endometrial carcinoma cells. Evid. Based Complement. Alternat. Med. 2012, 2012, 859185. [Google Scholar]
- Tai, C.J.; Wang, C.K.; Tai, C.J. Aqueous extract of Solanum nigrum leaves induces autophage and enhances cytotoxicity of cisplatin, doxoorubicin, docetaxel, and 5-flurouracil in human colorectal carcinoma cells. Evid. Based Complement. Alternat. Med. 2013, 2013, 514719. [Google Scholar]
- Ikeda, T.; Tsumagari, H.; Nohara, T. Steroidal oligoglycosides from Solanum nigrum. Chem. Pharm. Bull. 2000, 48, 1062–1064. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.Y.; Shen, X.F.; Wang, L.; Wu, Z.W.; Li, F.; Chen, B.; Zhang, G.L.; Wang, M.K. Bioactive steroidal alkaloids from the fruits of Solanum nigrum. Phytochemistry 2018, 147, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Wang, Y.; Yi, X.; He, X. Anti-inflammatory steroidal glycosides from the berries of Solanum nigrum L. (European black nightshade). Phytochemistry 2018, 148, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, A.; Tahazadeh, A.R.; Ebrahimzadeh, M.A. Investigation of composition extracts, biological activities and optimization of Solanum nigrum L. extraction growing in Iran. Pak. J. Pharm. Sci. 2017, 30, 473–480. [Google Scholar] [PubMed]
- Wang, H.C.; Chung, P.J.; Wu, C.H.; Lan, K.P.; Yang, M.Y.; Wang, C.J. Solanum nigrum L. polyphenolic extract inhibits hepatocarcinoma cell growth by inducing G2/M phase arrest and apoptosis. J. Sci. Food Agric. 2011, 91, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Syu, K.Y.; Lin, J.K. Chemical Composition of Solanum nigrum Linn Extract and Induction of Autophagy by Leaf Water Extract and Its Major Flavonoids in AU565 Breast Cancer Cells. J. Agric. Food Chem. 2010, 58, 8699–8708. [Google Scholar] [CrossRef]
- Zaidi, S.K.; Hoda, M.N.; Tabrez, S.; Ansari, S.A.; Jafri, M.A.; Khan, M.S.; Hasan, S.; Alqahtani, M.H.; Abuzenadah, A.M.; Banu, N. Protective Effect of Solanum nigrum Leaves Extract on Immobilization Stress Induced Changes in Rat’s Brain. Evid. Based Complement. Altern. Med. 2014, 2014, 912450. [Google Scholar] [CrossRef]
- Upadhyay, P.; Ara, S.; Prakash, P. Antibacterial and Antioxidant Activity of Solanum nigrum Stem and Leaves. Chem. Sci. 2015, 4, 1013–1017. [Google Scholar]
- Sun, A.Y.; Wang, Q.; Simonyi, A.; Sun, G.Y. Botanical Phenolics and Neurodegeneration. In Herbal Medicine Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Sissi Wachtel-Galor, S., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2011; pp. 315–325. [Google Scholar]
- Ogunsuyi, O.B.; Ademiluyi, A.O.; Oboh, G.; Oyeleye, S.I.; Dada, A.F. Green leafy vegetables from two Solanum spp. (Solanum nigrum L and Solanum macrocarpon L.) ameliorate scopolamine-induced cognitive and neurochemical impairments in rats. Food Sci. Nutr. 2018, 6, 860–870. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chndra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Zitka, O.; Skalickova, S.; Gumulec, J.; Masarik, M.; Adam, V.; Hubalek, J.; Trnkova, L.; Kruseova, J.; Eckschlager, T.; Kizek, R. Redox status expressed as GSH:GSSG ratio as a marker for oxidative stress in paediatric tumour patients. Oncol Lett. 2012, 4, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Duan, X.; Fang, X.; Shang, W.; Tong, C. Mitochondrial protein import regulates cytosolic protein homeostasis and neuronal integrity. Autophagy 2018, 14, 1293–1309. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.B.; Waagepetersen, H.S.; Bak, L.K.; Schousboe, A.; Sonnewald, U. The glutamine glutamate/GABA cycle: Function, regional differences in glutamate and GABA production and effects of interference with GABA metabolism. Neurochem. Res. 2015, 40, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Vezzani, A.; Najjar, S.; De Lanerolle, N.C.; Rogawski, M.A. Glia and epilepsy: Excitability and inflammation. Trends Neurosci. 2013, 36, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A.; Bak, L.K.; Waagepetersen, H.S. Astrocytic control of biosynthesis and turnover of the neurotransmitters glutamate and GABA. Front. Endocrinol. 2013, 4, 102. [Google Scholar] [CrossRef] [PubMed]
- Agbor, G.A.; Vinson, J.A.; Donnelly, P.E. Folin-Ciocalteau Reagent for Polyphenolic Assay. Int. J. Food Sci. Nutr. Diet. 2014, 3, 147–156. [Google Scholar] [CrossRef]
- Sak, K. Dependence of DPPH radical scavenging activity of dietary flavonoid quercetin on reaction environment. Mini-Rev. Med. Chem. 2014, 14, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Acquaviva, R.; Russo, A.; Galvano, F.; Galvano, G.; Barcellona, M.L.; Li Volti, G.; Vanella, A. Cyanidin and cyanidin 3-O-beta-D -glucoside as DNA cleavage protectors and antioxidants. Cell Biol. Toxicol. 2003, 19, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Campisi, A.; Caccamo, D.; Raciti, G.; Cannavò, G.; Macaione, V.; Currò, M.; Macaione, S.; Vanella, A.; Ientile, R. Glutamate-induced increases in transglutaminase activity in primary cultures of astroglial cells. Brain Res. 2003, 978, 24–30. [Google Scholar] [CrossRef]
- Li Volti, G.; Ientile, R.; Abraham, N.G.; Vanella, A.; Cannavò, G.; Mazza, F.; Currò, M.; Raciti, G.; Avola, R.; Campisi, A. Immunocytochemical localization and expression of heme oxygenase-1 in primary astroglial cell cultures during differentiation: Effect of glutamate. Biochem. Biophys. Res. Commun. 2004, 315, 517–524. [Google Scholar] [CrossRef]
- Acquaviva, R.; Campisi, A.; Murabito, P.; Raciti, G.; Avola, R.; Mangiamel, S.; Musumeci, I.; Barcellona, M.L.; Vanella, A.; Li Volti, G. Propofol attenuates peroxynitrite-mediated DNA damage and apoptosis in cultured astrocytes: An alternative protective mechanism. Anesthesiology 2004, 101, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.H.; Baraban, J.M. Glutamate toxicity in immature cortical neurons precedes development of glutamate receptor currents. Brain Res. Dev. Brain Res. 1990, 57, 146–150. [Google Scholar] [CrossRef]
- Chen, Y.H.; Du, G.H.; Zhang, J.T. Salvianolic acid B protects brain against injuries caused by ischemia-reperfusion in rats. Acta Pharmacol. Sin. 2000, 21, 463–466. [Google Scholar] [PubMed]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Nistico, S.; Ventrice, D.; Dagostino, C.; Lauro, F.; Ilari, S.; Gliozzi, M.; Strongoli, M.C.; Vecchio, I.; Rizzo, M.; Mollace, V.; Muscoli, C.; et al. Effect of MN (III) tetrakis (4-benzoic acid) porphyrin by photodynamically generated free radicals on SODs keratinocytes. J. Biol. Regul. Homeost. Agents 2013, 27, 781–790. [Google Scholar] [PubMed]
- Del Río, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar] [CrossRef] [PubMed]
- Carresi, C.; Musolino, V.; Gliozzi, M.; Maiuolo, J.; Mollace, R.; Nucera, S.; Maretta, A.; Sergi, D.; Muscoli, S.; Gratteri, S.; et al. Anti-oxidant effect of bergamot polyphenolic fraction counteracts doxorubicin-induced cardiomyopathy: Role of autophagy and c-kitposCD45negCD31neg cardiac stem cell activation. J. Mol. Cell. Cardiol. 2018, 119, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kaur, M.; Katnoria, J.K.; Nagpal, A.K. Polyphenols in Food: Cancer Prevention and Apoptosis Induction. Curr. Med. Chem. 2018, 25, 4740–4757. [Google Scholar] [CrossRef]
- Loffredo, L.; Perri, L.; Nocella, C.; Violi, F. Antioxidant and antiplatelet activity by polyphenol-rich nutrients: Focus on extra virgin olive oil and cocoa. Br. J. Clin. Pharmacol. 2017, 83, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.; Ventrice, D.; Giannetto, F.; Cirinnà, S.; Santagati, N.A.; Procopio, A.; Mollace, V.; Muscoli, C. Antioxidant activity of oleuropein and semisynthetic acetyl-derivatives determined by measuring malondialdehyde in rat brain. J. Pharm. Pharmacol. 2017, 69, 1502–1512. [Google Scholar] [CrossRef] [PubMed]
- Muscoli, C.; Lauro, F.; Dagostino, C.; Ilari, S.; Giancotti, L.A.; Gliozzi, M.; Costa, N.; Carresi, C.; Musolino, V.; Casale, F.; et al. Olea Europea-derived phenolic products attenuate antinociceptive morphine tolerance: An innovative strategic approach to treat cancer pain. J. Biol. Regul. Homeost. Agents 2014, 28, 105–116. [Google Scholar] [PubMed]
- Zhang, Y.; Li, X.; Wang, Z. Antioxidant activities of leaf extract of Salvia miltiorrhiza Bunge and related phenolic constituents. Food Chem. Toxicol. 2010, 48, 2656–2662. [Google Scholar] [CrossRef] [PubMed]
- Campisi, A.; Acquaviva, R.; Mastojeni, S.; Raciti, G.; Vanella, A.; De Pasquale, R.; Puglisi, S.; Iauk, L. Effect of berberine and Berberis aetnensis C. Presl. alkaloid extract on glutamate-evoked tissue transglutaminase up-regulation in astroglial cell cultures. Phytother. Res. 2001, 25, 816–820. [Google Scholar] [CrossRef]
- Pierozan, P.; Biasibetti, H.; Schmitz, F.; Ávila, H.; Parisi, M.M.; Barbe-Tuana, F.; Wyse, A.T.; Pessoa-Pureur, R. Quinolinic acid neurotoxicity: Differential roles of astrocytes and microglia via FGF-2-mediated signaling in redox-linked cytoskeletal changes. Biochim. Biophys. Acta 2016, 1863, 3001–3014. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, J.K.; Pandey, S.; Ribecco-Lutkiewicz, M.; Monette, R.; Borowy-Borowski, H.; Walker, P.R.; Sikorska, M. Molecular mechanisms of glutamate neurotoxicity in mixed cultures of NT2-derived neurons and astrocytes: Protective effects of coenzyme Q10. J. Neurosci. Res. 2003, 72, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Campisi, A.; Caccamo, D.; Li Volti, G.; Currò, M.; Parisi, G.; Avola, R.; Vanella, A.; Ientile, R. Glutamate-evoked redox state alterations are involved in tissue transglutaminase upregulation in primary astrocyte cultures. FEBS Lett. 2004, 578, 80–84. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Solvent | Physical | Yield % | Total Phenolic Content (mg/g of Extract) |
|---|---|---|---|---|
| SN1 | MeOH-H2O 80–20 v/v | Brown solid | 26.1 ± 3.9% | 92.2 ± 4.8 |
| SN2 | H2O | Brown solid | 19.5 ± 5.0% | 40.0 ± 6.9 |
| Extract | Gallic Acid | Protocatechuic Acid | Chlorogenic Acid | Gentisic Acid | Caffeic Acid | Luteolin | Apigenin |
|---|---|---|---|---|---|---|---|
| SN1 | 0.09 ± 0.02 | 0.24 ± 0.91 | 2.77 ± 0.45 | 1.50 ± 0.66 | 0.64 ± 0.87 | 0.98 ± 0.33 | 0.16 ± 0.74 |
| SN2 | 0.04 ± 0.05 | 0.19 ± 0.11 | 2.01 ± 0.98 | 1.81 ± 0.75 | 0.42 ± 0.54 | 0.8 ± 0.62 | 0.12 ± 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campisi, A.; Acquaviva, R.; Raciti, G.; Duro, A.; Rizzo, M.; Santagati, N.A. Antioxidant Activities of Solanum nigrum L. Leaf Extracts Determined in In Vitro Cellular Models. Foods 2019, 8, 63. https://doi.org/10.3390/foods8020063
Campisi A, Acquaviva R, Raciti G, Duro A, Rizzo M, Santagati NA. Antioxidant Activities of Solanum nigrum L. Leaf Extracts Determined in In Vitro Cellular Models. Foods. 2019; 8(2):63. https://doi.org/10.3390/foods8020063
Chicago/Turabian StyleCampisi, Agata, Rosaria Acquaviva, Giuseppina Raciti, Anna Duro, Milena Rizzo, and Natale Alfredo Santagati. 2019. "Antioxidant Activities of Solanum nigrum L. Leaf Extracts Determined in In Vitro Cellular Models" Foods 8, no. 2: 63. https://doi.org/10.3390/foods8020063
APA StyleCampisi, A., Acquaviva, R., Raciti, G., Duro, A., Rizzo, M., & Santagati, N. A. (2019). Antioxidant Activities of Solanum nigrum L. Leaf Extracts Determined in In Vitro Cellular Models. Foods, 8(2), 63. https://doi.org/10.3390/foods8020063