
Nonlinear SAR Modelling of Mosquito Repellents for Skin Application

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Repellent Activity
2.2. Molecular Descriptors
2.3. Nonlinear Supervised Machine Learning Method
2.4. Performance Evaluation Metrics
2.5. In Vivo Evaluation of Repellent Activity
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Devillers, J. Computational Design of Chemicals for the Control of Mosquitoes and Their Diseases; CRC Press: Boca Raton, CA, USA, 2018. [Google Scholar]
- WHO. Vector-Borne Diseases. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 1 September 2023).
- Melgarejo-Colmenares, K.; Cardo, M.V.; Vezzani, D. Blood feeding habits of mosquitoes: Hardly a bite in South America. Parasitol. Res. 2022, 121, 1829–1852. [Google Scholar] [CrossRef]
- Hernandez-Colina, A.; Gonzalez-Olvera, M.; Lomax, E.; Townsend, F.; Maddox, A.; Hesson, J.C.; Sherlock, K.; Ward, D.; Eckley, L.; Vercoe, M.; et al. Blood-feeding ecology of mosquitoes in two zoological gardens in the United Kingdom. Parasit. Vectors 2021, 14, 249. [Google Scholar] [CrossRef] [PubMed]
- Norris, E.J.; Coats, J.R. Current and future repellent technologies: The potential of spatial repellents and their place in mosquito-borne disease control. Int. J. Environ. Res. Public Health 2017, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.; Potter, C.J. Insect repellents mediate species-specific olfactory behaviours in mosquitoes. Malar. J. 2020, 19, 127. [Google Scholar] [CrossRef]
- Leal, W.S. The enigmatic reception of DEET—The gold standard of insect repellents. Curr. Opin. Insect Sci. 2014, 6, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.; Betz, J.F.; Riabinina, O.; Lahondère, C.; Potter, C.J. Commonly used insect repellents hide human odors from Anopheles mosquitoes. Curr. Biol. 2019, 29, 3669–3680. [Google Scholar] [CrossRef]
- Karcher, W.; Devillers, J. Practical Applications of Quantitative Structure-Activity Relationships (QSAR) in Environmental Chemistry and Toxicology; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Hansch, C.; Leo, A. Exploring QSAR: Fundamentals and Applications in Chemistry and Biology; American Chemical Society: Washington, DC, USA, 1995. [Google Scholar]
- Tinkov, O.V.; Grigorev, V.Y.; Grigoreva, L.D.; Osipov, V.N.; Kolotaev, A.V.; Khachatryan, D.S. QSAR analysis and experimental evaluation of new quinazoline-containing hydroxamic acids as histone deacetylase. SAR QSAR Environ. Res. 2022, 33, 513–532. [Google Scholar] [CrossRef]
- Devillers, J.; Devillers, H. Toxicity profiling and prioritization of plant-derived antimalarial agents. SAR QSAR Environ. Res. 2019, 30, 801–824. [Google Scholar] [CrossRef]
- Toropov, A.A.; Toropova, A.P.; Benfenati, E. The QSAR-search of effective agents towards coronaviruses applying the Monte Carlo method. SAR QSAR Environ. Res. 2021, 32, 689–698. [Google Scholar] [CrossRef]
- Wu, Y.; Huo, D.; Chen, G.; Yan, A. SAR and QSAR research on tyrosinase inhibitors using machine learning methods. SAR QSAR Environ. Res. 2021, 32, 85–110. [Google Scholar] [CrossRef]
- Devillers, J. 2D and 3D structure-activity modelling of mosquito repellents: A review. SAR QSAR Environ. Res. 2018, 29, 693–723. [Google Scholar] [CrossRef] [PubMed]
- Devillers, J.; Sartor, V.; Doucet, J.P.; Doucet-Panaye, A.; Devillers, H. In silico prediction of mosquito repellents for clothing application. SAR QSAR Environ. Res. 2022, 33, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Devillers, J.; Sartor, V.; Devillers, H. Predicting mosquito repellents for clothing application from molecular fingerprint-based artificial neural network SAR models. SAR QSAR Environ. Res. 2022, 33, 729–751. [Google Scholar] [CrossRef] [PubMed]
- Knippling, E.F.; McAlister, L.C.; Jones, H.A. Results of Screening Tests with Materials Evaluated as Insecticides, Miticides, and Repellents at the Orlando, Fla., Laboratory, April 1942 to April 1947; USDA Publication E-733; United States Department of Agriculture, Agriculture Research Administration, Bureau of Entomology and Plant Quarantine: Orlando, FL, USA, 1947. [Google Scholar]
- Moriguchi, I.; Hirono, S.; Liu, Q.; Nakagome, I.; Matsushita, Y. A simple method of calculating octanol/water partition coefficient. Chem. Pharm. Bull. 1992, 40, 127–130. [Google Scholar] [CrossRef]
- Moriguchi, I.; Hirono, S.; Nakagome, I.; Hirano, H. Comparison of reliability of log P values for drugs calculated by several methods. Chem. Pharm. Bull. 1994, 42, 976–978. [Google Scholar] [CrossRef]
- Devillers, J. Neural Networks in QSAR and Drug Design; Academic Press: London, UK, 1996. [Google Scholar]
- Grieco, J.P.; Achee, N.L.; Sardelis, M.R.; Chauhan, K.R.; Roberts, D.R. A novel high-throughput screening system to evaluate the behavioral response of adult mosquitoes to chemicals. J. Am. Mosq. Control Assoc. 2005, 21, 404–411. [Google Scholar] [CrossRef]
- WHO. World Health Organization, Guidelines for Efficacy Testing of Spatial Repellents, Control of Neglected Tropical Diseases; WHO Pesticide Evaluation Scheme: Geneva, Switzerland, 2013; Available online: https://www.who.int/publications/i/item/9789241505024 (accessed on 27 September 2023).
- Devillers, J. Strengths and weaknesses of the backpropagation neural network in QSAR and QSPR studies. In Neural Networks in QSAR and Drug Design; Devillers, J., Ed.; Academic Press: London, UK, 1996; pp. 1–46. [Google Scholar]
- Devillers, J.; Balaban, A.T. Topological Indices and Related Descriptors in QSAR and QSPR; Gordon and Breach Science Publishers: Amsterdam, The Netherlands, 1999. [Google Scholar]
- Balaban, A.T. Highly discriminating distance-based topological index. Chem. Phys. Lett. 1982, 89, 399–404. [Google Scholar] [CrossRef]
- Bertz, S.H. The first general index of molecular complexity. J. Chem. Am. Soc. 1981, 103, 3599–3601. [Google Scholar] [CrossRef]
- Hall, L.H.; Kier, L.B. Molecular connectivity Chi indices for database analysis and structure-property modeling. In Topological Indices and Related Descriptors in QSAR and QSPR; Devillers, J., Balaban, A.T., Eds.; Gordon and Breach Science Publishers: Amsterdam, The Netherlands, 1999; pp. 307–360. [Google Scholar]
- Kier, L.B.; Hall, L.H. The kappa indices for modeling molecular shape and flexibility. In Topological Indices and Related Descriptors in QSAR and QSPR; Devillers, J., Balaban, A.T., Eds.; Gordon and Breach Science Publishers: Amsterdam, The Netherlands, 1999; pp. 455–489. [Google Scholar]
- Suryanarayana, M.V.; Pandey, K.S.; Prakash, S.; Raghuveeran, C.D.; Dangi, R.S.; Swamy, R.V.; Rao, K.M. Structure-activity relationship studies with mosquito repellent amides. J. Pharm. Sci. 1991, 80, 1055–1057. [Google Scholar] [CrossRef]
- Rayner, H.B.; Wright, R.H. Far infrared spectra of mosquito repellents. Can. Entomol. 1966, 98, 76–80. [Google Scholar] [CrossRef]
- Mekenyan, O.; Bonchev, D.; Sabljic, A.; Trinajstic, N. Applications of topological indices to QSAR. The use of the Balaban index and the electropy index for correlations with toxicity of ethers on mice. Acta Pharm. Jugosl. 1987, 37, 75–86. [Google Scholar]
- Devillers, J.; Doucet, J.P.; Doucet-Panaye, A.; Decourtye, A.; Aupinel, P. Linear and non-linear QSAR modelling of juvenile hormone esterase inhibitors. SAR QSAR Environ. Res. 2012, 23, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Gramatica, P.; Kim, M.G.; Kim, D.; Tratnyek, P.G. QSAR modelling of water quality indices of alkylphenol pollutants. SAR QSAR Environ. Res. 2007, 18, 729–743. [Google Scholar] [CrossRef]
- Wang, J.; Si, H.; Liu, Y.; Song, J.; Wang, P.; Luo, H.; Chen, S.; Fan, G.; Rao, X.; Wang, Z.; et al. Experimental evaluation and structure-activity relationship analysis of bridged-ring terpenoid derivatives as novel Blattella germanica repellent. SAR QSAR Environ. Res. 2022, 33, 969–986. [Google Scholar] [CrossRef]
- Branco, P.; Torgo, L.; Ribeiro, R.P. A survey of predictive modeling on imbalanced domains. ACM Comput. Surv. 2016, 1, 1–56. [Google Scholar] [CrossRef]
- Garcia, V.; Mollineda, R.A.; Sanchez, J.S. A new performance evaluation method for two-class imbalanced problems. In Structural, Syntactic, and Statistical Pattern Recognition, Proceedings of the Joint IAPR International Workshop, SSPR & SPR 2008, Orlando, FL, USA, 4–6 December 2008; da Vitoria Lobo, N., Kasparis, T., Roli, F., Kwok, J.T., Georgiopoulos, M., Anagnostopoulos, G.C., Loog, M., Eds.; Lect. Notes Comput. Sci.. Springer: Geneva, Switzerland, 2008; pp. 917–925. [Google Scholar]
- Paluch, G.; Bartholomay, L.; Coats, J. Mosquito repellents: A review of chemical structure diversity and olfaction. Pest Manag. Sci. 2010, 66, 925–935. [Google Scholar] [CrossRef]
- Tunón, H.; Thorsell, W.; Mikiver, A.; Malander, I. Arthropod repellency, especially tick (Ixodes ricinus), exerted by extract from Artemisia abrotanum and essential oil from flowers of Dianthus caryophyllum. Fitoterapia 2006, 77, 257–261. [Google Scholar] [CrossRef]
- Cantrell, C.L.; Jones, A.M.; Ali, A. Isolation and identification of mosquito (Aedes aegypti) biting-deterrent compounds from the native American ethnobotanical remedy plant Hierochloë odorata (sweetgrass). J. Agric. Food Chem. 2016, 64, 8352–8358. [Google Scholar] [CrossRef]
- Khater, H.F.; Selim, A.M.; Abouelella, G.A.; Abouelella, N.A.; Murugan, K.; Vaz, N.P.; Govindarajan, M. Commercial mosquito repellents and their safety concerns. In Malaria; Kasenga, F.H., Ed.; IntechOpen Ltd.: London, UK, 2019; Available online: https://www.intechopen.com/chapters/68538 (accessed on 1 September 2023).
- Robert, L.L.; Hallam, J.A.; Seeley, D.C.; Roberts, L.W.; Wirtz, R.A. Comparative sensitivity of four Anopheles (Diptera: Culicidae) to five repellents. J. Med. Entomol. 1991, 28, 417–420. [Google Scholar] [CrossRef]
- Fossati, F.P.; Maroli, M. Laboratory tests of three repellents against Phlebotomus perniciosus (Diptera: Psychodidae). Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 771–773. [Google Scholar] [CrossRef]
- Devillers, J.; Larghi, A.; Lagneau, C. QSAR modelling of synergists to increase the efficacy of deltamethrin against pyrethroid-resistant Aedes aegypti mosquitoes. SAR QSAR Environ. Res. 2018, 29, 613–629. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Poupardin, R.; Darriet, F.; Reynaud, S.; Bonnet, J.; Strode, C.; Brengues, C.; Yébakima, A.; Ranson, H.; Corbel, V.; et al. Exploring the molecular basis of insecticide resistance in the dengue vector Aedes aegypti: A case study in Martinique Island (French West Indies). BMC Gen. 2009, 10, 494. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Carron, A.; Darriet, F.; Etienne, M.; Agnew, P.; Tolosa, M.; Yp-Tcha, M.M.; Lagneau, C.; Yébakima, A.; Corbel, V. Reduced efficacy of pyrethroid space sprays for dengue control in an area of Martinique with pyrethroid resistance. Am. J. Trop. Med. Hyg. 2009, 80, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Marcombe, S.; Mathieu, R.B.; Pocquet, N.; Riaz, M.A.; Poupardin, R.; Sélior, S.; Darriet, F.; Reynaud, S.; Yébakima, A.; Corbel, V.; et al. Insecticide resistance in the dengue vector Aedes aegypti from Martinique: Distribution, mechanisms and relations with environmental factors. PLoS ONE 2012, 7, e30989. [Google Scholar] [CrossRef] [PubMed]
- Devillers, J.; Pham-Delègue, M.H. Honey Bees: Estimating the Environmental Impact of Chemicals; Taylor & Francis: London, UK, 2002. [Google Scholar]
- Moreira-Filho, J.T.; Braga, R.C.; Lemos, J.M.; Alves, V.M.; Borba, J.V.V.B.; Costa, W.S.; Kleinstreur, N.; Muratov, E.N.; Andrate, C.H.; Neves, B.J. BeeToxAI: An artificial intelligence-based web app to assess acute toxicity of chemicals to honey bees. Artif. Intel. Life Sci. 2021, 1, 100013. [Google Scholar] [CrossRef]
- Wang, F.; Yang, J.F.; Wang, M.Y.; Jia, C.Y.; Shi, X.X.; Hao, G.F.; Yang, G.F. Graph attention convolutional neural network model for chemical poisoning of honey bees’ prediction. Sci. Bull. 2020, 65, 1184–1191. [Google Scholar] [CrossRef]
- WHO. Guidelines for Efficacy Testing of Mosquito Repellents for Human Skin; WHO/HTM/NTD/WHOPES/2009.4; WHO: Geneva, Switzerland, 2009; Available online: https://www.who.int/publications/i/item/WHO-HTM-NTD-WHOPES-2009.4 (accessed on 1 September 2023).
- Devillers, J.; Lagadic, L.; Yamada, O.; Darriet, F.; Delorme, R.; Deparis, X.; Jaeg, J.P.; Lagneau, C.; Lapied, B.; Quiniou, F.; et al. Use of multicriteria analysis for selecting candidate insecticides for vector control. In Juvenile Hormones and Juvenoids. Modeling Biological Effects and Environmental Fate; Devillers, J., Ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 333–366. [Google Scholar]
- Stanczyk, N.M.; Brookfield, J.F.; Field, L.M.; Logan, J.G. Aedes aegypti mosquitoes exhibit decreased repellency by DEET following previous exposure. PLoS ONE 2013, 8, e54438. [Google Scholar] [CrossRef]
- Yang, L.; Norris, E.J.; Jiang, S.; Bernier, U.R.; Linthicum, K.J.; Bloomquist, J.R. Reduced effectiveness of repellents in a pyrethroid-resistant strain of Aedes aegypti (Diptera: Culicidae) and its correlation with olfactory sensitivity. Pest Manag. Sci. 2020, 76, 118–124. [Google Scholar] [CrossRef]
- da Costa, K.S.; Galúcio, J.M.; da Costa, C.H.S.; Santana, A.R.; Dos Santos Carvalho, V.; do Nascimento, L.D.; Lima e Lima, A.H.; Neves Cruz, J.; Alves, C.N.; Lameira, J. Exploring the potentiality of natural products from essential oils as inhibitors of odorant-binding proteins: A structure- and ligand-based virtual screening approach to find novel mosquito repellents. ACS Omega 2019, 4, 22475–22486. [Google Scholar] [CrossRef]
- Zafar, Z.; Fatima, S.; Bhatti, M.F.; Shah, F.A.; Saud, Z.; Butt, T.M. Odorant binding proteins (OBPs) and odorant receptors (ORs) of Anopheles stephensi: Identification and comparative insights. PLoS ONE 2022, 17, e0265896. [Google Scholar] [CrossRef]
- Okoli, B.J.; Eltayb, W.A.; Gyebi, G.A.; Ghanam, A.R.; Ladan, Z.; Oguegbulu, J.C.; Abdalla, M. In silico study and excito-repellent activity of Vitex negundo L. Essential oil against Anopheles gambiae. Appl. Sci. 2022, 12, 7500. [Google Scholar]
- Devillers, J. Calculation of octanol/water partition coefficients for pesticides: A comparative study. SAR QSAR Environ. Res. 1999, 10, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Mannhold, R.; van de Waterbeemd, H. Substructure versus whole-molecule approaches for calculating log P. J. Comput. Aided Mol. Des. 2001, 15, 337–354. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | TP | TN | FP | FN | TTP | TTN |
|---|---|---|---|---|---|---|
| Training set | 243 | 1386 | 92 | 16 | 259 | 1478 |
| Test set | 50 | 336 | 39 | 9 | 59 | 375 |
| Whole set | 293 | 1722 | 131 | 25 | 318 | 1853 |
| Metrics | Training Set | Test Set | Whole Set |
|---|---|---|---|
| Sensitivity | 0.94 | 0.85 | 0.92 |
| Specificity | 0.94 | 0.90 | 0.93 |
| Accuracy | 0.94 | 0.89 | 0.93 |
| F1 | 0.82 | 0.68 | 0.79 |
| MCC | 0.79 | 0.63 | 0.76 |
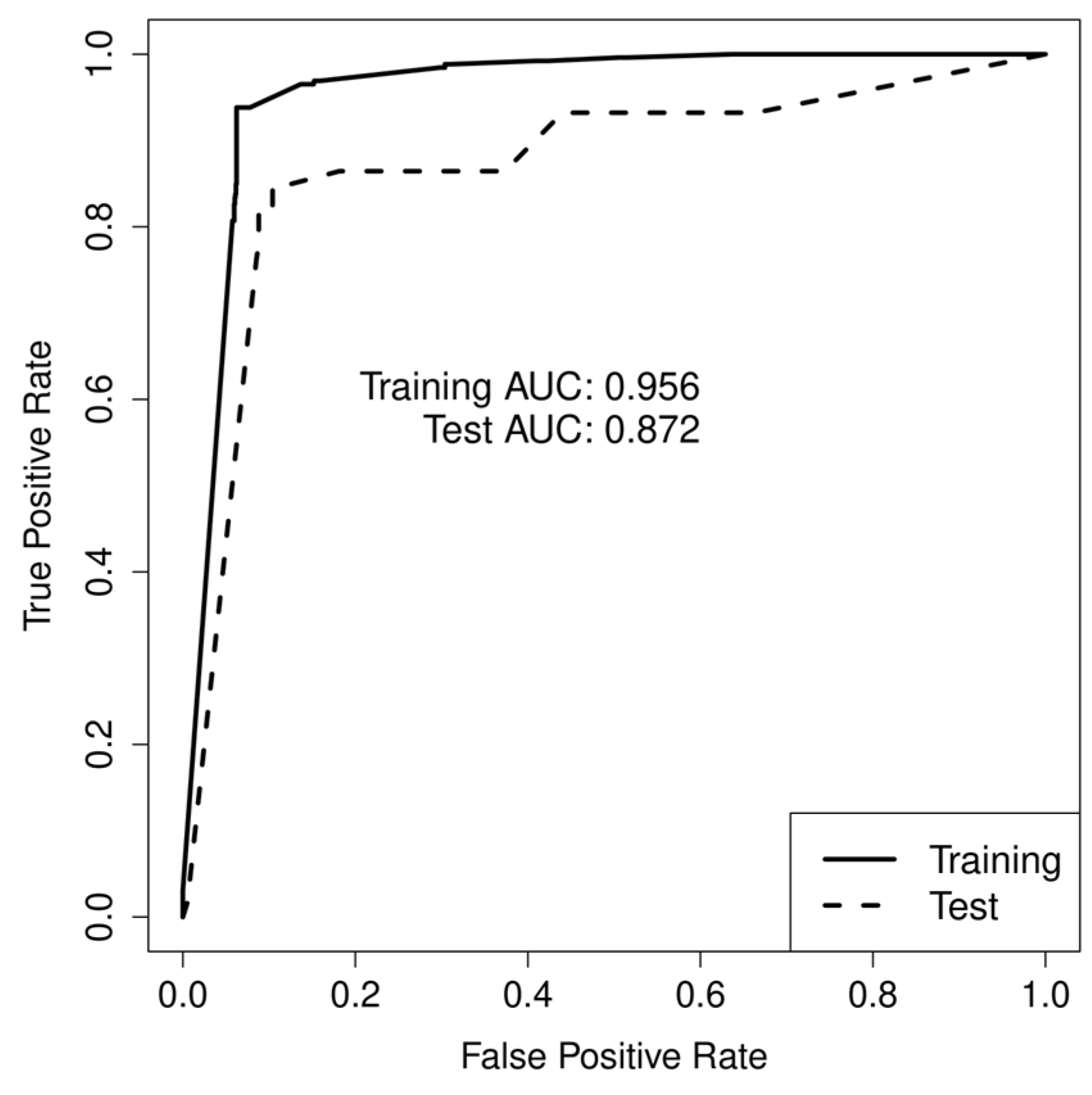
| AUC | 0.96 | 0.87 | 0.94 |
| G-mean | 0.94 | 0.87 | 0.93 |
| Dominance | 0 | −0.05 | −0.01 |
| Molecule | SAI * | Conf. int. | SAIW | Conf. int. |
|---|---|---|---|---|
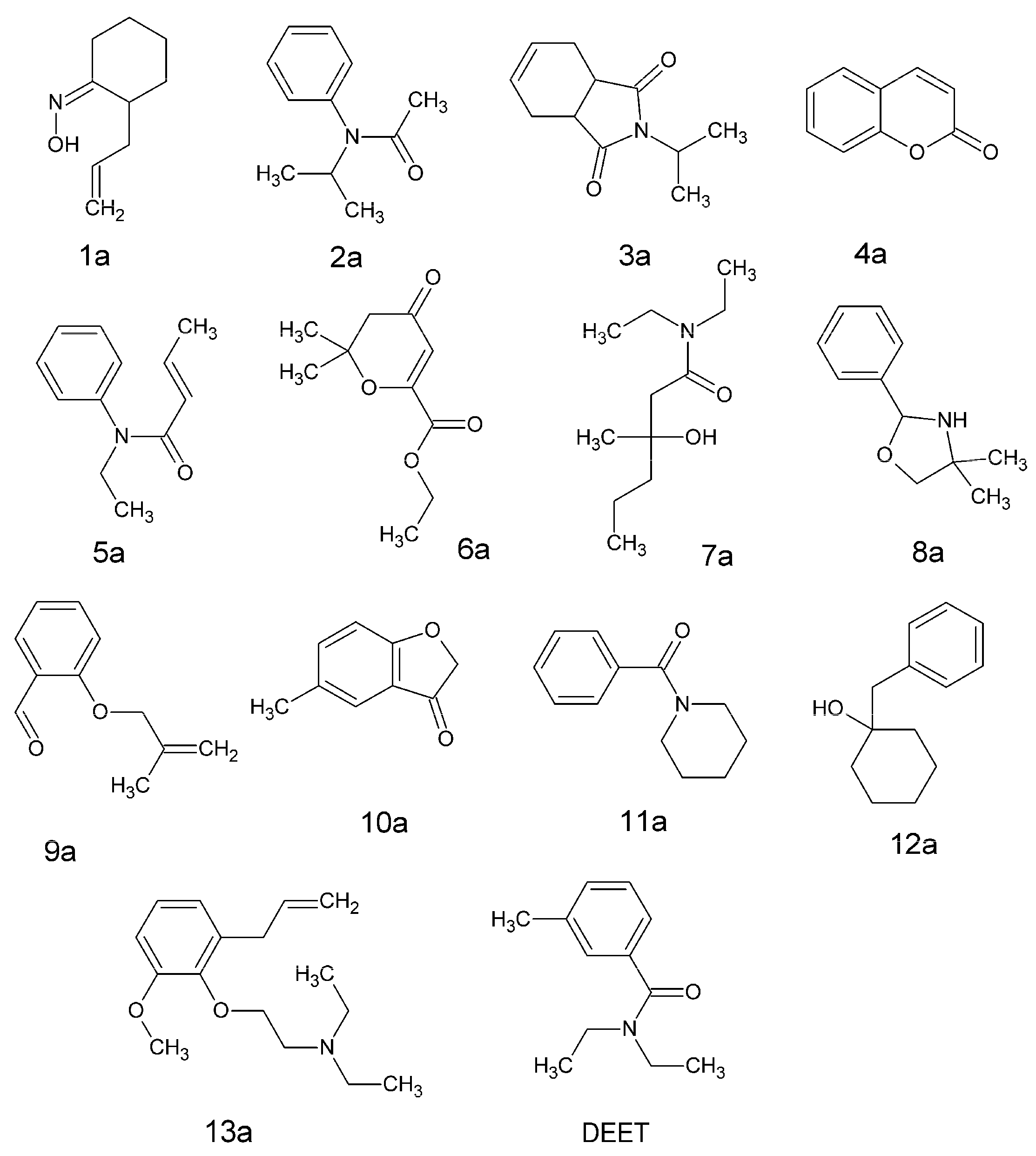
| DEET (7%) | 0.46 | 0.28–0.64 | 0.28 | 0.16–0.39 |
| 12a (7%) | 0.47 | 0.32–0.61 | 0.38 | 0.25–0.51 |
| 13a (7%) | 0.45 | 0.32–0.59 | 0.32 | 0.21–0.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devillers, J.; Larghi, A.; Sartor, V.; Setier-Rio, M.-L.; Lagneau, C.; Devillers, H. Nonlinear SAR Modelling of Mosquito Repellents for Skin Application. Toxics 2023, 11, 837. https://doi.org/10.3390/toxics11100837
Devillers J, Larghi A, Sartor V, Setier-Rio M-L, Lagneau C, Devillers H. Nonlinear SAR Modelling of Mosquito Repellents for Skin Application. Toxics. 2023; 11(10):837. https://doi.org/10.3390/toxics11100837
Chicago/Turabian StyleDevillers, James, Adeline Larghi, Valérie Sartor, Marie-Laure Setier-Rio, Christophe Lagneau, and Hugo Devillers. 2023. "Nonlinear SAR Modelling of Mosquito Repellents for Skin Application" Toxics 11, no. 10: 837. https://doi.org/10.3390/toxics11100837